Copyright2000 by the Genetics Society of America
Subtelomeric Repeat Amplification Is Associated With Growth at Elevated
Temperature in
yku70
Mutants of
Saccharomyces cerevisiae
Barbara Fellerhoff,* Friederike Eckardt-Schupp* and Anna A. Friedl*
,†*GSF-Forschungszentrum, Institut fu¨r Strahlenbiologie, 85758 Oberschleißheim, Germany and†Strahlenbiologisches Institut der Ludwig-Maximilians-Universita¨t, 80336 Munich, Germany
Manuscript received March 2, 1998 Accepted for publication November 15, 1999
ABSTRACT
Inactivation of the Saccharomyces cerevisiae gene YKU70 (HDF1), which encodes one subunit of the Ku heterodimer, confers a DNA double-strand break repair defect, shortening of and structural alterations in the telomeres, and a severe growth defect at 37⬚. To elucidate the basis of the temperature sensitivity, we analyzed subclones derived from rare yku70 mutant cells that formed a colony when plated at elevated temperature. In all these temperature-resistant subclones, but not in cell populations shifted to 37⬚, we observed substantial amplification and redistribution of subtelomeric Y⬘element DNA. Amplification of Y⬘elements and adjacent telomeric sequences has been described as an alternative pathway for chromosome end stabilization that is used by postsenescence survivors of mutants deficient for the telomerase pathway. Our data suggest that the combination of Ku deficiency and elevated temperature induces a potentially lethal alteration of telomere structure or function. Both in yku70 mutants and in wild type, incubation at 37⬚results in a slight reduction of the mean length of terminal restriction fragments, but not in a significant loss of telomeric (C1-3A/TG1-3)n sequences. We propose that the absence of Ku, which is known to bind
to telomeres, affects the telomeric chromatin so that its chromosome end-defining function is lost at 37⬚.
T
ELOMERES are specialized structures at the ends for 50–100 generations (Singer and Gottschling1994; Lendvay et al. 1996). The same characteristic
of linear chromosomes. In Saccharomyces cerevisiae,
telomeric DNA consists of a simple repeat tract, com- phenotypes have been observed in strains mutated for
the genes EST1, EST3, or EST4 (CDC13;Lundbladand
monly abbreviated as (C1-3A/TG1-3)n, which in wild-type
cells isⵑ300-bp long (for review seeZakian1996). The Szostak1989;LundbladandBlackburn1993;
Lend-vayet al. 1996;Nugent et al. 1996). Although not re-exact length of individual telomeric repeat tracts varies,
reflecting a dynamic state that is kept approximately quired for telomerase activity in vitro (Ligner et al.
1997b), the products of these genes are thought to be stable by a balance between lengthening and shortening
reactions. Telomeres shorten after each round of repli- involved in proper telomerase function in vivo, e.g., by
loading the telomerase onto the chromosomal ends (for cation because of the inability of DNA polymerase to
start de novo synthesis; after removal of the most distal review see Nugent andLundblad1998). The loss of
viability that is characteristic for tlc1 and est mutants
telomeric RNA primer a gap is left at the 5⬘end of the
has been termed senescence (LundbladandSzostak
newly synthesized strand. This shortening, as well as
1989). In late cultures of senescent strains so-called post-shortening potentially arising from nucleolytic
degra-senescence survivors arise, which employ a RAD52-de-dation, is counteracted by the ribonucleoenzyme
te-pendent, telomerase-independent mechanism of
chro-lomerase, which elongates the 3⬘ends of telomeric DNA
mosome end stabilization that is manifested by global
by adding TG1-3repeats (for reviews seeGreider1996;
amplification of subtelomeric Y⬘elements and telomeric
Zakian1996). The S. cerevisiae genes TLC1 and EST2
repeat tracts (LundbladandBlackburn1993;
Lend-encode the RNA and the protein component of the
vayet al. 1996). This alternative chromosome end
stabi-ribonucleoprotein telomerase, respectively (Singerand
lization may either proceed by recombination of
inter-Gottschling1994;Counteret al. 1997;Ligneret al.
nal (C1-3A/TG1-3)n sequences onto the chromosome
1997a). As expected, inactivation of one or both genes
ends, as was initially proposed (Lundbladand
Black-results in a progressive decline of the length of the
burn1993), or by a break-induced replication
mecha-terminal telomeric repeat tract with each round of
repli-nism. According to the break-induced replication cation, as was seen by shortening of terminal restriction
model, chromosome ends, which, after telomere attri-fragments (TRFs). This telomere shortening is
accom-tion, appear as sites of chromosome breaks, invade ho-panied by a considerable drop in viability after growth
mologous targets (i.e., telomeric or subtelomeric re-gions of other chromosome ends), and replicate all the sequences to the end of the “donor” chromosome
Corresponding author: A. A. Friedl, Strahlenbiologisches Institut der
(BoscoandHaber1998;Leet al. 1999).
Ludwig-Maximilians-Universita¨t, Schillerstr. 42, 80336 Mu¨nchen,
Ger-many. E-mail: [email protected] Shortening of telomeric repeat tracts may result not
only from telomerase deficiency, but also from a distur- meres and their localization near the nuclear periphery
are also affected by inactivation of Ku (Larocheet al.
bance of proper telomere length regulation. For
exam-ple, tel1 and tel2 mutants do not respond to factors that 1998). Although the TRF lengths are comparable in
Ku⫺and tel1 mutants, the latter are normal with respect
influence telomere length in wild-type cells, suggesting
that these mutants are impaired in telomere length reg- to rapid shortening of elongated telomeres (Liand
Lus-tig 1996), appearance of G-rich tails (Gravel et al.
ulation (Runge andZakian 1996). In these mutants,
which do not exhibit senescence, reduction of the mean 1998), and telomere clustering and localization (
Laro-cheet al. 1998), and they are affected only slightly with
TRF length by ⵑ200 bp becomes fully manifest only
after ⵑ100 generations of growth; thereafter a new respect to telomere position effect (Boultonand
Jack-son1998), demonstrating that these phenotypes in hdf1
steady-state level of telomere size is established (Lustig
andPetes1986;RungeandZakian1996). or yku80 mutants are not simply a consequence of short telomeres. Rather, the data suggest a distinctive role Reduction of the mean TRF length has also been
observed in yku70 and yku80 mutants (Boulton and for the Ku proteins at the telomeres. Indeed, in vivo
crosslinking demonstrated that Yku80p binds to
telo-Jackson1996b;Porteret al. 1996). YKU70 (HDF1) and
YKU80 (HDF2) encode proteins that exhibit significant meric DNA (Gravelet al. 1998).
The present report aims at further characterizing the homology to the mammalian Ku70 and Ku80 proteins,
respectively (Feldmann and Winnacker 1993; Feld- role of Yku70p in telomere metabolism. We show that,
even after prolonged subcultivation, inactivation of
mannet al. 1996;Milneet al. 1996). Like their
mamma-lian counterparts, Yku70p and Yku80p form a hetero- YKU70 does not affect cell viability at 30⬚, although a
new steady-state TRF length is established that isⵑ200
dimeric complex that binds with high affinity to
double-stranded DNA ends (Milneet al. 1996). By a variety of bp shorter than in wild-type cells. In spite of a severe
growth defect at 37⬚, some yku70 mutant cells are able different assays it has been shown that the yeast Ku-like
proteins are required for the repair of DNA double- to escape the temperature-induced lethality and form
colonies at 37⬚. We observed considerable, RAD52-strand breaks (DSBs) by a nonhomologous end-joining
pathway (BoultonandJackson1996a,b; Milneet al. dependent amplification of subtelomeric Y⬘ DNA in
these temperature-resistant subclones of yku70 mutant
1996; Tsukamoto et al. 1996; Friedl et al. 1998). In
addition to the repair deficiency, yku70 and yku80 mu- strains, and we propose that the absence of the Ku-like
tants display a considerable growth defect at elevated proteins at the telomeres together with some
tempera-temperature. On solid media at 37⬚, the plating effi- ture-dependent effect initiates a potentially lethal
pro-ciency is very low in both mutants and the cells appear cess that can be bypassed by a telomerase-independent,
to arrest in a large-budded state (FeldmannandWin- alternative mechanism of chromosome end
stabiliza-nacker1993;BoultonandJackson1996b;Feldmann tion. No indications for an increased loss of telomeric et al. 1996;Siedeet al. 1996;Barnes and Rio1997). (C1-3A/TG1-3)nDNA in yku70 mutants compared to
wild-Several recent investigations addressed the role of the type or tel1 mutants were found after a shift to elevated
yeast Ku-like proteins in telomere metabolism. It has temperature. We propose that telomeres in Ku-deficient
been shown that inactivation of YKU70 or YKU80 leads cells lose their telomere-defining chromatin structure
to an alteration of the structure of the chromosomal at elevated temperature and appear as sites of
chromo-termini, in that the G-rich strand forms a long single- somal breaks, thus leading to cellular response reactions
stranded tail during all phases of the cell cycle (Gravel like recombinative events and G2 arrest.
et al. 1998;Polotniankaet al. 1998). In wild-type cells, this tail is detectable in late S-phase only and is thought
to arise by specific 5⬘-3⬘ exonuclease activity (Wel- MATERIALS AND METHODS
linger et al. 1993, 1996). Individual, artificially
elon-Yeast strains:The strains used in this study are described
gated telomeres shrink to the strain-specific average in Table 1. The diploid YKU70/yku70⌬::URA3 strain WS9132,
length more rapidly in the absence than in the presence which was obtained by mating strain SX46A yku70⌬with strain
of Ku, leading to the proposal that Ku protects telomeres WS8105-1C (Siede et al. 1996), exhibits the same telomere
length as does a cross of wild-type strains SX46A and
WS8105-against nucleolytic degradation and recombinational
1C (A. A. Friedl, unpublished data). In the construct used
events (Polotnianka et al. 1998). Lack of functional
for one-step gene disruption (Rothstein1989) of the YKU70
Ku-like proteins has also been shown to result in a
defi-gene, the URA3 gene replaced a HindIII fragment of the
ciency in transcriptional repression of genes located YKU70 open reading frame (ORF), thus disrupting the ORF
close to the telomeres [telomere position effect (TPE)] at amino acid position 300 (Siedeet al. 1996). Strain WS9132
was sporulated using standard methods. Two complete tetrads
but not in derepression of the silent mating type loci
(spore clones 1a–d and 2a–d) were further investigated. Some
(Boulton and Jackson 1998; Evans et al. 1998;
of the results obtained with the spore clones were confirmed
Larocheet al. 1998;Nugentet al. 1998). Ku interacts
by experiments performed in strains of different genetic
back-with the silencing protein Sir4p, which may explain its ground carrying the same yku70⌬::URA3 allele or a
role in TPE (Tsukamotoet al. 1997). Fluorescence in yku70⌬::TRP1 allele disrupting the ORF at the same position.
These strains were derived from MK166 (Liefshitzet al. 1995)
telo-TABLE 1
Strains used in this study
Strain Relevant genotype Source
SX46A MATa ade2 his3-532 trp1-289 ura3-52 Siedeet al. (1996)
SX46A yku70⌬ SX46A yku70⌬::URA3 Siedeet al. (1996)
SX46A yku70⌬rad52⌬ SX46A yku70⌬::URA3 rad52::TRP1 Siedeet al. (1996)
SX46A tel1⌬ SX46A tel1⌬::TRP1 This study
WS8105-1C MAT␣ade2 agr4-17 trp1-289 ura3-52 Siedeet al. (1996)
WS8105-1C yku70⌬rad52⌬ WS8105-1C yku70⌬::URA3 rad52::TRP1 Siedeet al. (1996)
WS9132 SX46A yku70⌬X WS8105-1C Siedeet al. (1996)
Spore 1a From WS9132; YKU70 This study
Spore 1b From WS9132; yku70⌬::URA3 This study
Spore 1c From WS9132; yku70⌬::URA3 This study
Spore 1d From WS9132; YKU70 This study
Spore 2a From WS9132; YKU70 This study
Spore 2b From WS9132; YKU70 This study
Spore 2c From WS9132; yku70⌬::URA3 This study
Spore 2d From WS9132; yku70⌬::URA3 This study
or W303, both kindly donated by M. Kupiec. Because of the milliliters of the suspensions was used to generate frozen stocks, and the remaining suspension was used to prepare formal possibility that these yku70 disruption mutations allow
expression of a truncated Yku70 protein, some results were DNA for genomic Southern analysis.
Fluctuation analyses were performed by transferring 6–12 also confirmed in a strain with a genetic background similar
to that of the spore clones (i.e., SX46A) carrying a complete complete single colonies (obtained after growth at 30⬚for 3 days) to 1 ml water and plating onto YPD plates after appro-deletion of the YKU70 open reading frame. This was obtained
by PCR-based gene deletion (Baudinet al. 1993) with a PCR priate dilution. Plates were incubated at 30⬚or 37⬚for 4 days. To determine the median frequency of cells forming a colony product representing the URA3 gene flanked by sequences
homologous to sequences located 5⬘ of the start codon and at 37⬚per cell growing at 30⬚, the data of two to three experi-ments were pooled. The rates of formation of temperature-3⬘of the stop codon of the genomic YKU70 gene. For
construc-tion of tel1 mutants, the 5⬘flanking region and the first 1770 resistant cells per cell generation were estimated for each experiment individually, using the mh estimator, which ac-bp of the TEL1 ORF were amplified by PCR from strain SX46A.
After cloning, an 883-bp EcoRI-AflIII fragment encompassing counts for situations where only a proportion of the cells of a colony is plated under selective conditions (Jones et al.
the first 657 bp of the ORF, the start codon, and the promotor
region was released and substituted by an EcoRI-Cfr10I frag- 1994), and the mean of the values obtained in the different experiments is indicated.
ment encompassing the TRP1 gene. The resulting tel1⌬::TRP1
construct was released from the vector and transformed into Determination of telomere length and Yⴕamplification in genomic Southern blots:Genomic DNA was isolated using a strain SX46A to construct strain SX46A tel1⌬. Construction
of strains SX46A yku70⌬ rad52⌬ and WS8105-1C yku70⌬ method based on cell wall disintegration by vortexing with glass beads and phenol extraction (HoffmanandWinston
rad52⌬has been described (Siedeet al. 1996).
Subcultivation and determination of growth characteristics: 1987), digested overnight with XhoI using standard procedures (Sambrooket al. 1989), and separated on 0.8% agarose gels.
The dissected spore products of strain WS9132 were allowed
to grow on solid YPD medium at 30⬚for 3 days, and then the After electrophoresis, the gels were blotted by alkaline capil-lary transfer onto nylon membranes (Qiagen, Inc., Chats-colonies were completely transferred to 20 ml of liquid YPD
medium. After overnight growth at 30⬚, the cell titer was deter- worth, CA). For a determination of telomere length, the blots were preincubated in blocking buffer (5⫻SSC, 1% N-lauroyl-mined to allow an estimation of the number of generations
passed since spore germination. A total of 5⫻105cells were sarcosine, 1⫻Denhardt’s solution) for 1 hr at 34⬚, and were hybridized for 3 hr at 34⬚in fresh blocking buffer containing used to inoculate fresh medium, and aliquots representing
100 and 1000 cells were plated on solid medium and incubated oligonucleotide probes that were end-labeled with32P by poly-nucleotide kinase. To probe for the terminal telomeric tract, at 30⬚ (4 days) and 37⬚(up to 14 days) for a determination
of the plating efficiency. The remainder of the suspension oligonucleotides (CA)10(C1-3A)n (CCC ACA CAC CAC ACC
CAC AC) or (G1-3T)n(GTG TGG GTG TGG TGT GTG GG)
was concentrated by centrifugation, resuspended in liquid
complete medium containing 10% glycerol, and frozen at were used. After washing for 15 min at room temperature in 2⫻ SSC, 0.1% SDS, the blots were autoradiographed. The ⫺20⬚. These subcultivation steps were repeated six times to
monitor the cells at intervals of ⵑ15 generations between autoradiographs were scanned with a GT 9000 scanner (Ep-son, Torrance, CA) and evaluated using the RFLPscan software generation 30 and generation 120. Strain SX46A tel1⌬ was
subcultivated in a similar manner, except that transformants (Scanalytics, Billerica, MA) to determine the peak of the signal intensity distribution of the major telomeric band.
arising after one-step gene disruption on selective solid
me-dium were first cultivated for one round in selective liquid For an estimation of the amount of Y⬘elements, the blots were stripped and rehybridized first with a Y⬘probe and then medium before they were further subcultivated in YPD
me-dium. Successful disruption of the TEL1 gene was later verified with a probe for CEN4, which can be used as internal control for the amount of DNA loaded (Conradet al. 1990). The Y⬘
by Southern analysis. To isolate single cell subclones, colonies
(see Ja¨gerand Philippsen1989). CEN4 was probed with a gel-purified BstYI fragment isolated from plasmid pDJ3, which was kindly provided by P. Philippsen. The fragments were radiolabeled by the random prime method (Feinberg and Vogelstein 1983). Hybridization was performed overnight at 68⬚ using a blocking buffer suggested by the membrane manufacturer (2⫻SSC, 1% SDS, 0.5% milk powder, 0.75 mg/ ml denatured salmon sperm DNA). After standard washing procedures, the blots were exposed to Fujifilm imaging plates and evaluated with a Fujix BAS 1000 Bio Imaging Analyzer (Fuji Photo Film Co., Ltd., Japan). To estimate the factor of Y⬘ amplification, the hybridization signals obtained for the long and the short Y⬘band in each lane were normalized with respect to the signal of the internal CEN4 sequence and then divided by the value obtained in control lanes.
Figure1.—TRF length in spores 2a (YKU70) and 2c (yku-Probes were stripped by washing the membrane two times
70⌬::URA3) in dependence of the number of generations
for 1 hr in 0.1nNaOH, 0.1% SDS at 68⬚. Complete removal
passed since germination. XhoI-digested DNA was probed with of probes was verified by exposure to imaging plates forⵑ24
a (CA)10oligonucleotide recognizing the sequence of the ter-hr (i.e., at least tter-hree times longer than typically necessary to
minal telomeric repeats. The ends of chromosomes bearing detect and quantify freshly hybridized probes).
Y⬘ elements appear as broad band at the bottom of the gel Temperature shift experiments: Logarithmic starter
cul-(arrow), and the ends of chromosomes lacking Y⬘ elements tures (grown at 30⬚) were used to inoculate liquid medium to
are visible as additional bands (asterisks). Bands of higher give a titer of 1⫻105cells/ml. Parallel samples were incubated
molecular weight represent additional Y⬘-lacking chromosome under vigorous shaking at 30⬚and 37⬚, respectively. At given
ends and fragments with internally located CA repeats. Lanes timepoints, the titer of the suspension was determined and
1–4, DNA isolated from spore 2a after 30 (1), 60 (2), 90 (3), samples were drawn for DNA preparation and determination
and 120 (4) generations. Lanes 5–11, DNA isolated from spore of the plating efficiency at 30⬚. Southern blots of XhoI-digested
2c after 30 (5), 45 (6), 60 (7), 75 (8), 90 (9), 105 (10), DNA were hybridized successively with (C1-3A)n, (G1-3T)n, and
and 120 (11) generations. Lane 12, DNA isolated from strain
CEN4. Hybridization, exposure to imaging plates, and
strip-SX46A yku70⌬after subcultivation for⬎200 generations. Size ping were done as described in the preceding section.
markers on the right side indicate 4, 3, 2, 1.6, and 1 kb (from Electrophoretic karyotyping: Cultures of single cell
sub-top to bottom). clones were concentrated by centrifugation, and chromosomal
DNA was prepared by the agarose plug method as described (Friedlet al. 1995), except that the protocol was downscaled.
which cuts at a conserved site in the subtelomeric Y⬘
Agarose plugs containing chromosomal DNA were loaded
onto 0.9% agarose gels for pulsed field gel electrophoresis elements (ChanandTye1983), resolved by gel
electro-(PFGE). Electrophoresis was performed using a CHEF DRII phoresis, transferred to nylon membranes, and probed system (Bio-Rad, Richmond, CA) as described (Friedlet al. with (CA)
10or (G1-3T)n. Using this method, the terminal
1995). After electrophoresis the gels were stained with ethid- telomeric fragments of chromosome ends bearing Y⬘ ium bromide, photographed, and UV-irradiated for 20 sec to
elements migrate in a single broad band, while
subtel-nick high-molecular-weight DNA. Then, the gels were treated
and blotted as described above for genomic Southern analysis. omeric C1-3A/TG1-3arrays and chromosome ends lacking Hybridization using the Y⬘probe was performed as described the Y⬘elements result in additional bands (Walmsley above. Chromosome-specific hybridizations were performed andPetes1985).
as described (Friedlet al. 1998).
Figure 1 shows that the length of the fragments in the main telomeric band of spore clone 2a (YKU70) remains constant between 30 and 120 generations after
RESULTS
spore germination (the earliest and latest timepoints
A new steady-state level of telomere length is estab- investigated), with the maximum of the hybridization
lished in yku70 mutant spore clones: To analyze the signal locating atⵑ1330 bp. In spore clone 2c (yku70),
phenotype of yku70 mutants in dependence of the num- the length of the fragments in the main telomeric band
ber of generations passed since inactivation of the gene, is reduced byⵑ180 bp after 30 generations (maximum
a YKU70/yku70⌬::URA3 heterozygous strain (WS9132) signal at 1150 bp), and byⵑ210 bp after 45 generations
was sporulated, and the products of two complete tet- (maximum signal at 1120 bp). The minor bands
origi-rads (spores 1a-d and 2a-d) were characterized further. nating from chromosome ends lacking Y⬘sequences are
Southern hybridization experiments revealed that spore also shortened byⵑ200 bp in the yku70 mutant spore.
clones 1a, 1d, 2a, and 2b carry the YKU70 wild-type Between generations 45 and 120, no further alterations
gene, while spore clones 1b, 1c, 2c, and 2d carry the were detected, and the new telomere length was found
yku70 mutant allele (data not shown). Each spore clone to be the same as in a sample of strain SX46A yku70⌬
was repeatedly subcultivated in liquid medium at 30⬚ grown for⬎200 generations (Figure 1, lane 12). Similar
and samples were taken after each subcultivation step length reductions were also observed in spore clones
to determine the plating efficiency at 30⬚and 37⬚(see 1b, 1c, and 2d, and in a variety of yku70 mutant strains
below) and the telomere length. To analyze the telo- of different genetic background (data not shown; see
TABLE 2
Plating efficiency at 37ⴗafter serial subcultivation ofyku70mutant spore products
Plating efficiency (%)
Generationsa
Spore 30 45 60 75 90 105 120
1b 0.55 0.60 ⬍0.1 ⬍0.1 0.30 0.30 ⬍0.1
1c 0.10 ⬍0.1 ⬍0.1 ⬍0.1 ⬍0.1 ⬍0.1 ⬍0.1
2c 0.40 0.45 ⬍0.1 ⬍0.1 ⬍0.1 ⬍0.1 ⬍0.1
2d 0.45 0.80 ⬍0.1 ⬍0.1 ⬍0.1 1.0 ⬍0.1
aGenerations passed between spore germination and plating at 37⬚.
conclude that telomeres do not shorten progressively 42 colonies of spores 1b, 2c, and 2d that arose during
the subcultivation experiment after plating at 37⬚were
in yku70 mutant cells, but rather that a new equilibrium
length is established after growth for ⵑ30–45 genera- picked and expanded in liquid medium at 30⬚. DNA
isolation, XhoI digest, and hybridization with a (CA)10
tions.
Inactivation ofYKU70does not affect viability at 30ⴗ, oligonucleotide revealed that the mean TRF length in these subclones is comparable to that observed in
con-but impairs colony formation at 37ⴗ:To determine the
trol samples grown at 30⬚ (Figure 2). In all samples
growth characteristics in dependence of the replicative
derived from temperature-resistant subclones, however, age, samples taken after the individual subcultivation
we observed that, at comparable amounts of total DNA steps were plated on solid medium and incubated at
loaded, a band ofⵑ7-kb length hybridized substantially
30⬚or 37⬚. In all eight spore clones, the plating efficiency
stronger than in the control samples. In
temperature-at 30⬚ was similar (close to 100%), and it remained
resistant subclones of spores 1b and 2c also a second constant between 30 and 120 generations of growth
band ofⵑ5.5 kb gave strongly enhanced hybridization
after sporulation. Furthermore, the colony size as well as the cellular yield after each subcultivation step were comparable for all spores (data not shown). Similarly,
even after extensive subcultivation (i.e., for⬎200
gener-ations), we have never detected any indication for
growth defects at 30⬚in any of our various yku70 mutant
strains (including a strain lacking the entire YKU70 open reading frame).
When plated at 37⬚, in the wild-type spore clones 1a,
1d, 2a, and 2b the plating efficiency at 37⬚was as high
as at 30⬚and remained constant until generation 120.
In contrast, already at generation 30 after germination,
the plating efficiency at 37⬚ was very low in the yku70
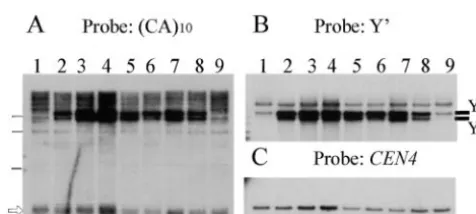
Figure2.—Y⬘element amplification in
temperature-resis-mutant spore clones 1b, 1c, 2c, and 2d, and it remained tant survivors isolated from spore 1b (yku70⌬::URA3). DNA
low (fluctuating between⬍0.1 and 1%) until generation from control clones of spore 1b grown at 30⬚(lanes 1 and 9)
and from temperature-resistant survivor clones of spore 1b
120 (Table 2). Many of the temperature-resistant
colo-that were obtained in the subcultivation experiment (lanes
nies became visible only after incubation for⬎8 days.
2–8) was digested with XhoI. Clone identification numbers of
Appearance of temperature-resistant subclones was also temperature-resistant clones are 320 (lane 2), 322 (3), 326
observed in yku70 mutant strains of different genetic (4), 324 (5), 325 (6), 323 (7), and 319 (8). (A) Probing with
background (A. A.Friedl, unpublished results). Gener- a labeled (CA)10oligonucleotide shows that the length of the
major TRF class (arrow) is similar in controls and
temperature-ally, colonies emerging from temperature-resistant
resistant subclones. Horizontal bars on the left edge indicate yku70 mutant cells were variable in size and often
irregu-6, 4, and 2 kb (from top to bottom). (B) Rehybridization with
lar in shape. When single cells derived from tempera- a Y⬘ probe reveals that the strong bands ofⵑ5.5 and 7 kb
ture-resistant subclones were replated at 37⬚, the plating represent the two size classes of Y⬘ elements, Y⬘long and Y⬘
efficiency generally was low (⬍30%), suggesting that short. The 5.5-kb band in the control samples reproduces
badly, but is clearly visible on the original autoradiographs.
the ability to grow at 37⬚is not stably transmitted.
Additional bands of higher molecular weight represent frag-Yⴕ elements are amplified in temperature-resistant
ments containing the most centromere-proximal Y⬘elements subclones of yku70 mutant spores: To elucidate the and adjacent sequences. (C) Rehybridization with a CEN4
mechanism by which the yku70 mutant cells obtain an probe labels an internal fragment that is used as a standard
for the amount of DNA loaded.
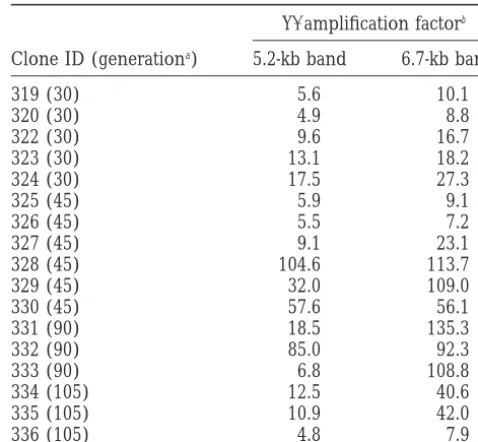
TABLE 3 signals (see Figure 2A for examples derived from spore
1b). Amplification of YⴕDNA in temperature-resistant
This phenotype was strongly reminiscent of that ob- subclones ofyku70spore 1b
served in postsenescence survivors of senescent mutants,
where fragments containing unit Y⬘elements and short Y⬘amplification factorb
stretches of internal telomeric sequence were found to
Clone ID (generationa) 5.2-kb band 6.7-kb band
be strongly amplified (LundbladandBlackburn1993;
319 (30) 5.6 10.1
Lendvay et al. 1996). Indeed, rehybridization of the
320 (30) 4.9 8.8
Southern blots with a Y⬘probe showed that the strongly
322 (30) 9.6 16.7
hybridizing fragments in the temperature-resistant
sub-323 (30) 13.1 18.2
clones represent Y⬘elements (Figure 2B). Furthermore,
324 (30) 17.5 27.3
we found that derivatives of spores 1b and 2c contain 325 (45) 5.9 9.1
both size classes of Y⬘elements (6.7 and 5.2 kb), where- 326 (45) 5.5 7.2
as in derivatives from spore 2d only the long Y⬘elements 327 (45) 9.1 23.1
328 (45) 104.6 113.7
appear to be present (data not shown).
329 (45) 32.0 109.0
To allow an estimation of the extent of Y⬘
amplifica-330 (45) 57.6 56.1
tion, in a subset of the temperature-resistant clones, the
331 (90) 18.5 135.3
signal intensities of both Y⬘bands were quantified after
332 (90) 85.0 92.3
hybridization with the Y⬘ probe and were normalized 333 (90) 6.8 108.8
against an internal standard that was obtained by subse- 334 (105) 12.5 40.6
quent rehybridization with a probe for CEN4. Using this 335 (105) 10.9 42.0
336 (105) 4.8 7.9
assay, we find that the amount of Y⬘DNA varies up to
2-fold between individual single cell clones of wild-type aGenerations passed between spore germination and
plat-spore 2b (data not shown). This level of variation, which ing at 37⬚.
is seen in both size classes of Y⬘ elements, probably bRatio between normalized Y⬘ hybridization signal
intensi-ties in temperature-resistant clones and 30⬚ control samples
reflects the normal recombinative activity at Y⬘elements
on the same gel. Normalization was performed with respect
(LouisandHaber1990a) and/or the technical
limita-to the CEN4 signal after rehybridization.
tions of the quantification procedure. In contrast, when
estimating the amount of Y⬘ DNA in 17
temperature-resistant subclones of spore 1b (Table 3) and 8
sub-banding pattern. The limited sensitivity of this method clones of spore 2d (data not shown), we observed a
precludes a detection of small alterations in chromo-clear increase compared to control samples that were
some length, especially if all chromosome species have
derived from subclones grown at 30⬚ and run on the
gained a similar and comparatively low number of new same gels. Table 3 shows that amplification factors differ
Y⬘elements. If, however, individual chromosome ends
between individual temperature-resistant clones,
rang-have acquired an exceedingly high number of new
re-ing fromⵑ5- to⬎100-fold, and that the relative degree
peats, a clear shift in the localization of the respective
of amplification of the two classes of Y⬘elements within
bands should occur, which can be detected easily upon given clones is variable.
visual inspection of the gels. Indeed, we detected in 27/ When nine subclones derived from yku70 mutant
42 samples a clearly visible shift of at least one
chromo-spore 1b grown on solid medium at 30⬚were analyzed
somal band; in 5 of these samples three or more bands in a similar manner, the normalized signal intensities
were found to be shifted (Figure 3A). The fuzziness
of theⵑ7-kb band varied less than twofold between the
of the altered bands (see below) precluded a detailed clones. In eight of these clones, also the signal intensities
analysis of the karyotypic alterations in samples carrying
of the ⵑ5.5-kb band varied less than twofold. In the
more than two shifted bands, but in those samples that
remaining clone, the intensity of theⵑ5.5-kb band was
were amenable to further analysis, the altered bands found to be 3.5 times stronger than the median intensity
were always found to be shifted towards higher molecu-of the other samples (data not shown). These results
lar weight, with apparent increases in chromosome size
suggest that Y⬘amplification is rare and comparatively
of up toⵑ100 kb. Southern hybridization using a probe
low in yku70 mutant cells when grown at 30⬚.
for the Y⬘elements appeared to give particularly high
Some chromosome ends acquire exceedingly high
signals with some of the lengthened chromosomal
numbers of Yⴕelements:To investigate whether, in the
bands (Figure 3, B and C), strongly suggesting that
course of Y⬘ amplification, a comparable number of
amplification of Y⬘elements is a major factor
contribut-repeat units is gained by all chromosome ends or
ing to the increase in chromosome size. In some cases, whether some individual ends acquire a
disproportion-the location of a shifted band was not immediately visi-ately high number of repeats, we separated the
chromo-ble in the altered banding pattern. For example, in somal DNA of the 42 temperature-resistant yku70
in temperature-resistant subclones of spore 1b, these
chromosomes hybridize with probes for Y⬘(Figure 3, B
and C), hinting at a redistribution of Y⬘ elements to
new chromosomal sites. Since bands indicative for
chro-mosome ends lacking Y⬘elements were still visible when
DNA of temperature-resistant subclones was digested with XhoI (Figure 2), however, it appears that within a population derived from a temperature-resistant colony
not all chromosome ends carry Y⬘elements.
When we karyotyped subclones of yku70 mutant spores 1b, 1c, 2c, and 2d that were obtained in the subcultivation experiment after plating at 30⬚, we found fuzzy lengthening of one chromosome in 1 out of 56 subclones analyzed; the karyotypes of the other
sub-Figure3.—Karyotypic alterations in temperature-resistant
clones were normal. Assuming that chromosome
elon-subclones of spore 1b (yku70⌬::URA3). A shows the negative
image of a PFGE gel after staining with ethidium bromide. gation is at least in part due to Y⬘ amplification, this
Lane 1, control sample of spore 1b grown at 30⬚. Lanes 2–6, shows again that the amplification process occurs rarely samples taken from temperature-resistant subclones 319 (lane in cells grown at 30⬚.
2), 322 (3), 324 (4), 321 (5), and 334 (6). Note that the
Amplification of Yⴕ elements in yku70 mutants re-amounts of DNA loaded on lanes 1 and 2 are similar; the
quires RAD52: Amplification of subtelomeric repeats
other lanes contain more DNA. Arrows indicate examples
for band shifts in temperature-resistant subclones. Asterisks in postsenescence survivors of senescent yeast mutants
indicate lost chromosomal bands in subclones 319 (band of depends on RAD52 (LundbladandBlackburn1993; chromosome I absent) and 334 (band of chromosome VI Lendvayet al. 1996). To estimate the rate of formation absent). B shows the results obtained after hybridizing a blot of
of temperature-resistant cells in yku70 rad52 as
com-the gel shown in A with a probe for Y⬘elements and overnight
pared to yku70 RAD52 strains, we performed fluctuation
exposure (only lanes 1 and 2 are shown). Indicated are the
positions of chromsomes VI and XI, which have gained Y⬘ analyses with the yku70 mutant spore clones 1c and 2d
elements in the temperature-resistant subclones. C shows the and two yku70 rad52 double mutant strains that were same blot after exposure for 5 hr. Note that shifted bands derived from strains SX46A and WS8105-1C, respec-often give rise to a particularly strong signal (arrows).
tively. All strains had been propagated for⬎150
genera-tions at 30⬚prior to the fluctuation experiments. The
frequency of temperature-resistant clones per cell in the (estimated size 260 kb) seems to be lacking (Figure 3A,
lane 2), but hybridization with a chromosome-specific colony varied from⬍0.2⫻10⫺3to 21⫻10⫺3(median
frequency 6.5⫻10⫺3) in spore 1c cells and from⬍1.2⫻
probe showed that chromosome I in this sample has a
new size ofⵑ300 kb, thus leading to comigration with 10⫺3 to 360⫻ 10⫺3(median frequency 3.9 ⫻ 10⫺3) in
spore 2d cells. This high range of frequencies suggests chromosome VI (data not shown).
Mostly, the bands of the lengthened chromosomes that at least a part of the temperature-resistant cells did
arise by spontaneous mutations as the colony grew. The were more fuzzy than normal chromosomal bands, thus
suggesting heterogeneous chromosome lengths within rates of formation of temperature-resistant clones in
spores 1c and 2d were estimated as 4.7⫻10⫺4and 34⫻
the cell population. In addition, we observed in many
clones a high degree of heterogeneous smear over the 10⫺4 per cell division, respectively. In the two yku70
rad52 double mutant strains, the median frequencies whole length of the gel lane (see, e.g., clone 334 in
lane 6 in Figure 3A). This kind of smear does not arise of temperature-resistant cells were 1.5⫻ 10⫺6 (range
⬍0.3 ⫻ 10⫺6 to 10 ⫻ 10⫺6) for strain SX46A yku70⌬
normally when applying our methods of DNA isolation
(see, e.g., the control lane 1 in Figure 3A), and it ap- rad52⌬ and 1.0 ⫻ 10⫺6 (range ⬍0.6 ⫻ 10⫺6 to 26 ⫻
10⫺6) for strain WS8105-1C yku70⌬ rad52⌬. The rates
peared to hybridize strongly with a Y⬘ probe (Figure
3C, lane 6). Furthermore, Southern hybridization in a of formation of temperature cells per division were
found to be three orders of magnitude lower than in sample of clone 334 with a probe for chromosome VI
(whose 300-kb band is absent in the karyotype) gave a the yku70 single mutants (3.6 ⫻ 10⫺7 and 7.3 ⫻ 10⫺7
per cell division for strains SX46A yku70⌬rad52⌬and
broad band ranging from ⵑ300 to 600 kb (data not
shown), suggesting that the smear is mainly caused by WS8105-1C yku70⌬ rad52⌬, respectively). These data
show that the ability to form temperature-resistant cells heterogeneous increase in chromosome lengths and
not by DNA degradation. in yku70 mutants depends largely on a functional RAD52
gene.
Like most strains of S. cerevisiae (Louis and Haber
1990b), spore clone 1b (grown at 30⬚) does not carry We investigated the structure of telomeric and
telo-mere-associated DNA in seven independent
tempera-Y⬘elements on all chromosomal termini; rather, the Y⬘
element is absent from chromosomes VI and XI, as is ture-resistant subclones derived from strains SX46A
Figure4.—Structure of telomeric and telomere-associated DNA in temperature-resistant subclones of yku70 rad52 double mutants. Lane 1, control clone of strain WS8105-1C yku70⌬
rad52⌬grown at 30⬚. Lanes 2–5, independent temperature-resistant subclones of the same strain. Lanes 6–8, independent temperature-resistant subclones of strain SX46A yku70⌬
rad52⌬. Lane 9, control clone of the same strain grown at 30⬚. (A) Probing with (G1-3T)nreveals that the terminal telomeric
fragments in the major band (arrow) are elongated in the temperature-resistant clones as compared to the respective control samples. Horizontal bars indicate 6, 3, 2, and 1 kb
Figure5.—TRF length in subclones of spore 2b (YKU70) (from top to bottom). (B) Probing with a Y⬘ fragment and
after plating at 30⬚ (lanes 1–3) or 37⬚ (lanes 4–7). Single (C) with a CEN4 fragment show that the relative amount of
cell clones were expanded at 30⬚before genomic DNA was Y⬘DNA in the temperature-resistant subclones is not increased
isolated, digested with XhoI, separated, and probed with radio-as compared to the control samples. This wradio-as also verified
labeled (CA)10. In the samples grown at 37⬚, the length of the by quantitative evaluation. Note that the derivatives of strain
fragments in the main telomeric band (arrow) giving maxi-WS8105-1C do not carry short Y⬘elements.
mum hybridization signal is reduced. Horizontal bars indicate 6, 3, 2, 1.6, and 1 kb, from top to bottom.
not find indications for increased amounts of Y⬘DNA
in these subclones after hybridizing with probes for Y⬘ Our data suggest that temperature-induced lethality
and CEN4 (Figure 4, B and C). Hybridization with the in yku70 mutants is related to some alteration in
telo-(G1-3T)noligonucleotide, however, revealed that in these mere length, structure, or functionality at elevated
tem-subclones the main telomeric fragments are on average perature, which can be circumvented by amplification
50 bp longer than in the parental strains (Figure 4A). of subtelomeric DNA or processes leading to telomere
When cells derived from these temperature-resistant elongation. To test whether this telomere alteration is
yku70 rad52 clones were replated at 37⬚, the plating simply a consequence of the already reduced telomere
efficiency was found to be high (close to 100%), indicat- length in Ku-deficient mutants, we included a tel1
mu-ing that the temperature-resistant phenotype in these tant strain in our studies. A fresh tel1 disruption mutant
clones is stable. Most probably, temperature resistance (strain background SX46A) was serially subcultivated
in the yku70 rad52 double mutant strains is conferred by to allow analysis of its phenotype at regular intervals
extragenic suppressor mutations that increase telomere between generations 45 and 120. In line with data
re-length or stability, but the exact mechanism remains to ported by others (LustigandPetes 1986;Rungeand
be elucidated. Zakian1996;BoultonandJackson1998), we found
Wild-type andtel1mutant clones grown at 37ⴗdo not that the tel1 mutants acquire a new equilibrium TRF show Yⴕamplification:When we analyzed the phenotype length ofⵑ1080 bp afterⵑ100 generations of growth,
of temperature-resistant yku70 mutant clones, we in- and that the plating efficiency is similar at 30⬚and 37⬚
cluded wild-type clones as controls. A total of 56 colonies (data not shown). We analyzed 20 subclones obtained
derived from YKU70 spores were picked from plates after colony formation at 37⬚, which were derived from
exposed to 37⬚and expanded in liquid medium at 30⬚ cells that had undergone 120 generations since
inactiva-before DNA isolation. To our surprise, we observed telo- tion of TEL1. Similar to what was seen in wild-type cells,
mere shortening in these clones, i.e., as compared to we observed that the terminal telomeric fragments are
samples that were exposed to a maximal temperature slightly shorter in these clones than in clones exposed
of 30⬚; the length of the terminal telomeric fragments to a maximal temperature of 30⬚, but we did not find
giving maximum hybridization signal was reduced by any indications for Y⬘ amplification (data not shown).
30–80 bp (Figure 5). This slight reduction in size, which Since the equilibrium length of the terminal telomeric
is reproducibly detectable, also occurred in strains of fragment in strain SX46A tel1⌬is shorter than in strain
different genetic background (data not shown), sug- SX46A yku70⌬, a short telomere alone appears not to
gesting thermolability of some aspect of telomere me- be sufficient for rendering the cells sensitive to elevated
tabolism. However, no indication for an amplification temperature, nor is the combination of short telomere
of Y⬘ elements was found in these YKU70 subclones, and elevated temperature sufficient for evoking
ampli-nor did we detect any chromosomal alteration in PFGE fication of Y⬘elements.
yku70mutant cells do not lose their telomeres upon
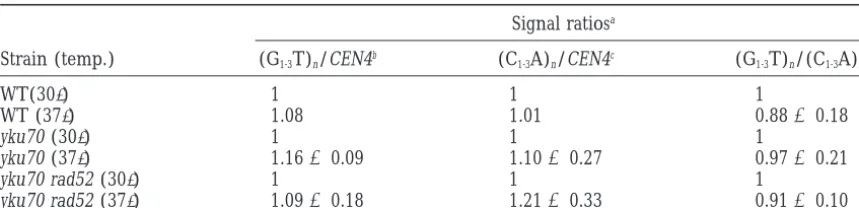
metabolism. Figure 6 and Table 4 also show that the ratio between the signals in the main telomeric band obtained after probing for the C-rich strand and the G-rich strand, respectively, is comparable at both incu-bation temperatures in wild-type, yku70 mutant, and yku70 rad52 double mutant cells. This argues against a further loss of the C-rich strand in Ku-deficient strains at elevated temperature.
To test whether, apart from the slight reduction in size, a substantial loss of telomeric DNA occurs in our strains upon incubation at 37⬚, some of the blots were also hybridized with a control sequence (CEN4) to allow a quantitation of the amount of DNA loaded. Normaliza-tion of the telomere hybridizaNormaliza-tion signal intensities with respect to the signal intensity of the internal control did not give any indication for a substantial loss of telomeric
Figure 6.—Quantitation of telomere DNA by determina- DNA in the Ku-deficient mutants (SX46A yku70⌬ and tion of the hybridization signal intensity after shifting liquid SX46A yku70⌬
rad52⌬) at elevated temperature (Figure cultures to 37⬚. Early logarithmic cultures of wild-type SX46A
6 and Table 4). The apparent discrepancy between the
(lanes 1 and 2) and SX46A yku70⌬(lanes 3 and 4) pregrown
at 30⬚were split and grown for 24 hr at 30⬚(lanes 1 and 3) data reported by us andGraveland co-workers (1998)
or 37⬚(lanes 2 and 4), respectively, before genomic DNA was on the one hand andBoultonandJackson(1998) on isolated, digested with XhoI, separated, and probed with (C1-3 the other hand may be attributable to different genetic A)n(A), (G1-3T)n(B), and CEN4 (C). The arrow indicates the
backgrounds or experimental designs. It is, however,
major telomeric band; horizontal bars indicate 6, 3, 2, and
unlikely that our treatment was not sufficient to induce
1 kb (from top to bottom).
temperature-dependent lethality, since ⵑ55% of the
yku70 cells were unable to form a colony at 30⬚ after
having been kept at 37⬚for 24 hr.
a shift to elevated temperature:Recently it was reported
It should be noted that we did not find indications that Ku-deficient cells exhibit a considerable loss of
ter-for an increase in the amount of Y⬘DNA in samples of
minal telomeric DNA when shifted to the restrictive
yku70 cells shifted to 37⬚when analyzing blots like that
temperature (BoultonandJackson 1998), and such
depicted in Figure 6. This indicates that Y⬘amplification
a loss could easily explain the necessity for alternative
does not occur unspecifically in all Ku-deficient cells, means of chromosome end stabilization in yku70
mu-but rather that it is a feature specific for those rare cells tants growing at 37⬚. Such a loss was, however, not
evi-that escape lethality at the restrictive temperature. dent from a second report on a temperature-shift
exper-iment (Gravelet al. 1998). Since Boulton and Jackson
used a probe detecting the C-rich strand, while Gravel DISCUSSION
and co-workers used a probe detecting the G-rich strand,
In this work, we aimed at further characterizing the it seemed possible that these controversial results were
role of Yku70p in telomere metabolism. We show that, due to a specific further degradation of the C-rich strand
after inactivation of the YKU70 function, shortening
in Ku-deficient mutants shifted to 37⬚. To test this possi- of the terminal telomeric repeat tract proceeds rather
bility, early-logarithmic cultures in liquid medium were
rapidly and that a new steady-state telomere length is
shifted to 37⬚for 24 hr. Under these conditions, wild- established, which isⵑ200 bp shorter than in congenic
type cells (strain SX46A) divide about eight times, and YKU70 strains. Several recent investigations show that
they continue to grow when kept at 37⬚for another 24 the absence of a functional Ku protein leads to a variety
hr. In contrast, yku70 mutant cells (strain SX46A of substantial alterations in telomere structure and
func-yku70⌬) double about four times in 24 hr before cessa- tion (see Introduction). One might expect that these
tion of further growth, and they arrest in a large-budded alterations had some negative influence on viability or
state. XhoI-digested DNA isolated from the shifted sam- growth rate. However, a detailed analysis of these
param-ples and from parallel samparam-ples kept at 30⬚was electro- eters over 120 generations after sporulation revealed
phoresed, blotted, and hybridized successively with (C1-3 that yku70 mutant spores and their YKU70 siblings did
A)nand (G1-3T)n. Figure 6 shows that in wild-type and not differ in these regards when grown at 30⬚. In
addi-yku70 mutants incubation at 37⬚ results in a slight tion, we never found indications for reduced growth at
(ⵑ20–30 bp) reduction of the mean length of the termi- 30⬚in any of our various yku70 mutant strains. Hence,
nal telomeric fragments. Comparable size reductions it appears that the telomeres in Ku-deficient cells are
were also observed in a tel1 mutant and in a yku70 rad52 still sufficiently stable and functional to allow normal
strain (data not shown). These data hint again at a growth at 30⬚. It should be noted, however, that in one
TABLE 4
Hybridization signal intensity in the major telomeric band after temperature shift to 37ⴗ
Signal ratiosa
Strain (temp.) (G1-3T)n/CEN4b (C1-3A)n/CEN4c (G1-3T)n/(C1-3A)nd
WT(30⬚) 1 1 1
WT (37⬚) 1.08 1.01 0.88⫾0.18
yku70 (30⬚) 1 1 1
yku70 (37⬚) 1.16⫾0.09 1.10⫾0.27 0.97⫾0.21
yku70 rad52 (30⬚) 1 1 1
yku70 rad52 (37⬚) 1.09⫾0.18 1.21⫾0.33 0.91⫾0.10
WT, wild type.
aHybridization signal ratios were normalized with respect to the 30⬚sample.
bRatio between the signal intensity of the major telomeric band after hybridization with (G
1-3T)nand the
signal intensity of the CEN4 fragment. Indicated are mean and SD from two blots (one experiment), except for WT (one blot).
cRatio between the signal intensity of the major telomeric band after hybridization with (C
1-3A)nand the
signal intensity of the CEN4 fragment. Indicated are mean and SD from two blots (one experiment), except for WT (one blot).
dRatio of the signal intensity in the major telomeric band after hybridization with (G
1-3T)n and (C1-3A)n,
respectively. Indicated are mean and SD from five blots (three independent experiments).
on growth at 30⬚was described (BarnesandRio1997), these bands were not seen in samples of est1 survivors
(Lundbladand Blackburn 1993). While the reason which may be explained by differing genetic
back-grounds. for this difference remains to be elucidated, it may hint
at a differential behavior of Y⬘-containing ends versus Ku-deficient mutants display a severe growth defect
at 37⬚(FeldmannandWinnacker1993;Boultonand Y⬘-lacking ends in Ku-deficient cells.
In this and previous (Friedl et al. 1998) work we
Jackson1996b;Feldmannet al. 1996;Siedeet al. 1996;
BarnesandRio1997), but we observed in yku70 mutant observed that Y⬘ amplification or fuzzy chromosomal banding occurs, with a low frequency, also in yku70 strains of various genetic backgrounds that a small
pro-portion of cells is able to escape temperature-induced mutant clones grown at 30⬚, while these events have
never been detected in wild-type cells so far. The high lethality and to form colonies at this temperature. In
this study, we analyzed 42 subclones derived from colo- variability in the frequency of temperature-resistant
yku70 cells seen in the fluctuation experiments suggests nies that appeared after plating yku70 mutant spore
products at 37⬚. As compared to control samples grown that events leading to temperature resistance can occur
spontaneously during colony growth at the permissive at 30⬚, all of the temperature-resistant subclones
exhib-ited a strong increase in the number of XhoI-fragments temperature, although we cannot exclude at the
mo-ment that additional events (which may be responsible
containing unit Y⬘elements. A similar phenomenon has
so far been observed only in postsenescence survivors for late-appearing colonies) are induced by the selective
treatment. Whether selected for or induced by incuba-derived from strains mutated for TLC1, EST1, EST2,
EST3, or EST4/CDC13 (Lundblad and Blackburn tion at 37⬚, we propose that the Y⬘amplification process enables growth at the elevated temperature in yku70
1993;Lendvayet al. 1996;Nugentet al. 1996). For the
sake of simplicity we refer to this phenomenon as Y⬘ cells. This assumption is based on the observation that
all temperature-resistant yku70 single mutant subclones amplification, but it should be borne in mind that
cur-rent models predict a simultaneous amplification and investigated so far exhibited amplification of
subtelo-meric DNA, whereas this amplification is not evident in redistribution of internal and/or distal telomeric
se-quences (LundbladandBlackburn1993;Boscoand liquid cultures of yku70 cells that were shifted to and
held at 37⬚for 24 hr (after which most cells are dead).
Haber1998;Leet al. 1999).
Postsenescence survivors and temperature-resistant Hence, Y⬘amplification correlates with temperature
re-sistance and does not occur unspecifically in all cells at yku70 clones share similarity with regard to the
instabil-ity of the phenotype after replating, the RAD52-depen- elevated temperature.
How can Y⬘amplification confer viability to
Ku-defi-dence of the amplification process, the Y⬘element
am-plification factors, and the transfer of Y⬘elements onto cient cells growing at elevated temperature? Given that
Y⬘ amplification is not the only mechanism enabling
chromosome ends previously lacking these elements.
However, bands indicative for chromosome ends lack- growth at elevated temperatures in Ku-deficient cells
(see below), we consider it unlikely that the
viability-ing Y⬘elements were readily detectable in our samples
of temperature-resistant yku70 clones after hybridiza- conferring effect is due to Y⬘-specific features, such as
ARS consensus sequences (ChanandTye1980) or the
helicase-like protein encoded by part of the naturally chromatin in a way that its telomere-defining activity is lost. This may be a consequence of the further reduction
occurring Y⬘elements (Yamadaet al. 1998). We rather
assume that the rescuing effect of the mechanism that of telomere length that may further destabilize
Ku-lack-ing telomeric chromatin because of a reduction of the
manifests itself as Y⬘amplification is conferred by
recom-binative amplification and redistribution of telomeric number of other telomere-associated proteins.
Alterna-tively, some protein or its interaction with DNA or other sequences, similar to what has been proposed to explain
the suppression of senescence in yeast mutants deficient proteins may be less stable at 37⬚, and Ku may have
a role in ensuring sufficent stability of the telomeric
in the telomerase pathway (McEachern and
Black-burn1996). chromatin under these conditions.
In this scenario, the Y⬘ amplification mechanism
What kind of alteration occurs at telomeres of
Ku-deficient cells upon incubation at elevated temperature? could rescue the cells by compensating for the
tempera-ture-induced telomere shortening and maintaining a In mutants deficient for the telomerase pathway, loss
of viability is thought to occur once a critical telomere length similar to that seen at 30⬚(Figure 2), thus
ensur-ing the necessary number of protein bindensur-ing sites. In size limit is reached after replication-associated
shorten-ing. It seemed possible that a critical telomere size limit a similar way telomere elongation, as seen in the very
rare temperature-resistant yku70 rad52 clones, could fa-could be reached in Ku-deficient cells incubated at 37⬚
because of a temperature-dependent further reduction cilitate proper chromatin formation. The observation
that the severity of the TPE deficiency in Ku⫺mutants
of the telomere length. Indeed, in yku70 mutant cells,
the average TRF length, as determined by the location increases with temperature (Evanset al. 1998;Nugent
et al. 1998) also supports our assumption that telomeric of the maximum hybridization signal, is decreased after
incubation at the elevated temperature. The extent of chromatin is thermolabile in the absence of Ku.
Overex-pression of EST1, EST2, or TLC1 in Ku-deficient mutants the size reduction, however, is comparable to that seen
in wild-type and tel1 mutant cells. Since the average TRF not only suppresses temperature sensitivity (Nugentet
al. 1998) but also alleviates the TPE defect at elevated length after the temperature shift in the tel1 mutant is
slightly shorter than in the yku70 mutant, it appears that temperature (Evanset al. 1998). So far, the telomere
lengths resulting from overexpression of individual the reduced average telomere length per se cannot be
responsible for temperature-induced lethality. components of the telomerase pathway have not been
investigated; recent data, however, are consistent with Although we cannot exclude that our methods to
determine telomere length and amount of telomeric an assumed role for these proteins in formation of
telo-meric chromatin that is independent of increased te-DNA are not sensitive enough to allow detection of
small but biologically significant differences between lomerase activity (Evanset al. 1998). Finally, we found
that temperature resistance of yku70 mutant cells is in-yku70 mutants and tel1 mutants, we do not think that
the absence of Ku leads to a more drastic temperature- creased when they are incubated in media containing
1 m sorbitol (B. Feller-Hoff, unpublished results).
induced shortening or loss of telomeric sequences than
that occurring in Ku-proficient cells (Figure 6). Rather, Sorbitol is known to stabilize proteins and
protein-pro-tein interactions in vitro (e.g.,Wimmeret al. 1997); it has
we think that in the absence of Ku the normal
tempera-ture-induced telomere shortening is no longer tolerated been proposed that sorbitol can also stabilize cellular
proteins in cells grown in its presence (Heath et al.
by the cell or that some other, not length-related,
alter-ation of the telomere conformalter-ation occurs. Ku is a telo- 1995). By whatever mechanism caused, a
temperature-induced instability of the telomeric chromatin may mere-binding protein, and it may, directly or indirectly,
constitute an important factor for the establishment of cause the loss of the privileged status of the chromosome
ends, thus making them appear as sites of chromosomal telomeric chromatin. Telomeres exhibit a special type
of chromatin organization in which the DNA is not breaks. Alternatively, a loss of defined telomere
struc-ture may affect proper chromosome segregation in mi-bound by nucleosomes, but rather by so-called
teloso-mal proteins, including Rap1p (for review seeZakian tosis. Additional investigations will be required to clarify
this point. 1996). This specific chromatin organization is known
to be responsible for the TPE; it may also have a role We gratefully acknowledge K. W. Runge for helpful comments, U.
in differentiating telomeric ends from DNA ends at sites Hamm for expert technical assistance, M. Kistler and U. Hoffmann
for spore dissection, M. Kupiec, P. Philippsen, W. Siede, and W.
of DSBs. In the absence of Ku, formation of proper
Oertel for providing strains and plasmids, and E. Fritz for valuable
telomeric chromatin seems to be impaired, as inferred
discussions. Part of this work was supported by the Commission of
from the reduction of TPE seen in Ku-deficient cells
the European Communities, grant FI4P-CT95-0010.
even at permissive temperature. Still, the telomere-defining function of telomeric chromatin appears to suffice when Ku-deficient cells grow at 30⬚, since their
LITERATURE CITED
viability is not reduced and indications for alternative
Barnes, G.,andD. Rio,1997 DNA double-strand-break sensitivity,
chromosome end stabilizations are rarely found. We
DNA replication, and cell cycle arrest phenotypes of Ku-deficient
propose that the combination of elevated temperature Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 94: 867–872.
andC. Cullin,1993 A simple and efficient method for direct Lundblad,1996 Senescence mutants of Saccharomyces cerevisiae with a defect in telomere replication identify three additional gene deletion in Saccharomyces cerevisiae. Nucleic Acids Res.
21:3329–3330. EST genes. Genetics 144: 1399–1412.
Li, B.,andA. J. Lustig,1996 A novel mechanism for telomere size
Bosco, G.,andJ. E. Haber,1998 Chromosome break-induced DNA
replication leads to nonreciprocal translocations and telomere control in Saccharomyces cerevisiae. Genes Dev. 10: 1310–1326.
Liefshitz, B., A. Parket, R. MayaandM. Kupiec,1995 The role capture. Genetics 150: 1037–1047.
Boulton, S. J.,andS. P. Jackson,1996a Saccharomyces cerevisiae Ku70 of DNA repair genes in recombination between repeated se-quences in yeast. Genetics 140: 1199–1211.
potentiates illegitimate DNA double-strand break repair and
serves as a barrier to error-prone DNA repair pathways. EMBO Ligner, J., T. R. Hughes, A. Shevchenko, M. Mann, V. Lundblad
et al., 1997a Reverse transcriptase motifs in the catalytic subunit J. 15: 5093–5103.
Boulton, S. J.,andS. P. Jackson,1996b Identification of a Saccharo- of telomerase. Science 276: 561–567.
Ligner, J., T. R. Cech, T. R. HughesandV. Lundblad,1997b Three
myces cerevisiae Ku80 homologue: roles in DNA double strand
break rejoining and in telomeric maintenance. Nucleic Acids ever shorter telomere (EST) genes are dispensable for in vitro yeast telomerase activity. Proc. Natl. Acad. Sci. USA 94: 11190– Res. 24: 4639–4648.
Boulton, S. J.,andS. P. Jackson,1998 Components of the Ku- 11195.
Louis, E. J.,andJ. E. Haber,1990a Mitotic recombination among dependent non-homologous end-joining pathway are involved in
telomeric length maintenance and telomeric silencing. EMBO subtelomeric Y⬘repeats in Saccharomyces cerevisiae. Genetics 124: 547–559.
J. 17: 1819–1828.
Chan, C. S. M.,andB.-K. Tye,1980 Autonomously replicating se- Louis, E. J.,andJ. E. Haber,1990b The subtelomeric Y⬘element family in Saccharomyces cerevisiae: an experimental system for re-quences in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 77:
6329–6333. peated sequence evolution. Genetics 124: 533–545.
Lundblad, V.,andE. H. Blackburn,1993 An alternative pathway
Chan, C. S. M.,andB.-K. Tye,1983 Organization of DNA sequences
and replication origins at yeast telomeres. Cell 33: 563–573. for yeast telomere maintenance rescues est1⫺senescence. Cell
73:347–360.
Conrad, M. N., J. H. Wright, A. J. WolfandV. A. Zakian,1990
RAP1 protein interacts with yeast telomeres in vivo: overproduc- Lundblad, V., andJ. W. Szostak,1989 A mutant with a defect in telomere elongation leads to senescence in yeast. Cell 57: tion alters telomere structure and decreases chromosome
stabil-ity. Cell 63: 739–750. 633–643.
Lustig, A. J.,andD. T. Petes,1986 Identification of yeast mutants
Counter, C. M., M. Meyerson, E. N. EatonandR. A. Weinberg,
1997 The catalytic subunit of yeast telomerase. Proc. Natl. Acad. with altered telomere structure. Proc. Natl. Acad. Sci. USA 83: 1398–1402.
Sci. USA 94: 9202–9207.
Evans, S. K., M. L. Sistrunk, C. I. NugentandV. Lundblad,1998 McEachern, M. J.,andE. H. Blackburn,1996 Cap-prevented re-combination between terminal telomeric repeat arrays (telomere Telomerase, Ku, and telomeric silencing in Saccharomyces
cerevis-iae. Chromosoma 107: 352–358. CPR) maintains telomeres in Kluyveromyces lactis lacking te-lomerase. Genes Dev. 10: 1822–1834.
Feinberg, A.,andB. Vogelstein, 1983 A technique for
radiola-belling DNA restriction fragments to high specific activity. Anal. Milne, G. T., S. Jin, K. B. ShannonandD. T. Weaver,1996 Muta-tions at two Ku homologs define a DNA end-joining repair path-Biochem. 132: 6–13.
Feldmann, H.,andE. L. Winnacker,1993 A putative homologue way in Saccharomyces cerevisiae. Mol. Cell. Biol. 16: 4189–4198.
Nugent, C. I.,andV. Lundblad,1998 The telomerase reverse tran-of the human autoantigen Ku from Saccharomyces cerevisiae. J. Biol.
Chem. 268: 12895–12900. scriptase: components and regulation. Genes Dev. 12: 1073–1085.
Nugent, C. I., T. R. Hughes, N. F. Lue and V. Lundblad, Feldmann, H., L. Driller, B. Meier, G. Mages, J. Kellermannet
al., 1996 HDF2, the second subunit of the Ku homologue from 1996 Cdc13p: a single-strand telomeric DNA-binding protein with a dual role in yeast telomere maintenance. Science 274:
Saccharomyces cerevisae. J. Biol. Chem. 271: 27765–27769.
Friedl, A. A., A. KraxenbergerandF. Eckardt-Schupp,1995 Use 249–252.
Nugent, C. I., G. Bosco, L. O. Ross, S. K. Evans, A. P. Salinger
of pulsed-field gel electrophoresis for studies of DNA
double-strand break repair in the yeast Saccharomyces cerevisiae. Methods et al., 1998 Telomere maintenance is dependent on activities required for end repair of double-strand breaks. Curr. Biol. 8:
7:205–218.
Friedl, A. A., M. Kiechle, B. FellerhoffandF. Eckardt-Schupp, 657–660.
Polotnianka, R. M., J. LiandA. J. Lustig, 1998 The yeast Ku 1998 Radiation-induced chromosome aberrations in
Saccharo-myces cerevisiae: influence of DNA repair pathways. Genetics 148: heterodimer is essential for protection of the telomere against nucleolytic and recombinational activities. Curr. Biol. 8: 831–834. 975–988.
Gravel, S., M. Larrivee, P. LabrecqueandR. J. Wellinger,1998 Porter, S. E., P. W. Greenwell, K. B. RitchieandT. D. Petes,1996 The DNA-binding protein Hdf1p (a putative Ku homologue) is Yeast Ku as a regulator of chromosomal DNA end structure.
Science 280: 741–744. required for maintaining normal telomere length in Saccharomyces
cerevisiae. Nucleic Acids Res. 24: 582–585.
Greider, C. W.,1996 Telomere length regulation. Annu. Rev.
Bio-chem. 65: 337–365. Rothstein, R.,1989 Targeting, disruption, replacement, and allelic rescue: integrative DNA transformation in yeast. Methods
Enzy-Heath, C. V., C. S. Copeland, D. C. Amberg, V. del Priore, M.
Snyderet al., 1995 Nuclear pore complex clustering and nu- mol. 194: 281–301.
Runge, K. W.,and V. A. Zakian, 1996 TEL2, an essential gene
clear accumulation of poly(A)⫹RNA associated with mutation
of the Saccharomyces cerevisiae RAT2/NUP120 gene. J. Cell Biol. required for telomere length regulation and telomere position effect in Saccharomyces cerevisiae. Mol. Cell. Biol. 16: 3094–3105.
131:1677–1697.
Hoffman, C. S.,andF. Winston,1987 A ten-minute DNA prepara- Sambrook, J., E. F. FritschandT. Maniatis,1989 Molecular Clon-ing: A Laboratory Manual. Cold Spring Harbor Laboratory Press,
tion from yeast efficiently releases autonomous plasmids for
trans-formation of Escherichia coli. Gene 57: 267–272. Cold Spring Harbor, NY.
Siede, W., A. A. Friedl, I. Dianova, F. Eckardt-SchuppandE. C. Ja¨ger, D.,andP. Philippsen,1989 Stabilization of dicentric
chro-mosomes in Saccharomyces cerevisiae by telomere addition to bro- Friedberg, 1996 The Saccharomyces cerevisiae Ku autoantigen homologue affects radiosensitivity only in the absence of homolo-ken ends or by centromere deletion. EMBO J. 8: 247–254.
Jones, M. E., S. M. ThomasandA. Rogers,1994 Luria-Delbru¨ ck gous recombination. Genetics 142: 91–102.
Singer, M. S.,andD. E. Gottschling,1994 TLC1: template RNA
fluctuation experiments: design and analysis. Genetics 136: 1209–
1216. component of Saccharomyces cerevisiae telomerase. Science 266: 404–409.
Laroche, T., S. G. Martin, M. Gotta, H. C. Gorham, F. E. Pryde
et al., 1998 Mutation of yeast Ku genes disrupts the subnuclear Tsukamoto, Y., J. KatoandH. Ikeda,1996 Hdf1, a yeast Ku-protein homologue, is involved in illegitimate recombination, but not in organization of telomeres. Curr. Biol. 8: 653–656.
Le, S., K. J. Moore, J. E. HaberandC. W. Greider,1999 RAD50 homologous recombination. Nucleic Acids Res. 24: 2067–2072.
Tsukamoto, Y., J. KatoandH. Ikeda,1997 Silencing factors partici-and RAD51 define two pathways that collaborate to maintain
telomeres in the absence of telomerase. Genetics 152: 143–152. pate in DNA repair and recombination in Saccharomyces cerevisiae. Nature 388: 900–903.