Received April 26, 1957
comparison of ring chromosomes in maize and Drosophila shows a striking
A
dissimilarity in their behavior. Whereas all the rings described to date in maize are unstable, owing to the formation of dicentrics, the Drosophila rings are by comparison extremely stable. Recent studies on the unstable w v c chromo- some in Drosophila have been interpreted as indicating that the observed insta- bility of this chromosome is independent of the ring structure(HINTON
1957). I n contrast to Drosophila, two unstable ring chromosomes showing a high fre- quency of discentric double-sized rings at anaphase have been reported in human tumor tissue (LEVAN 1956). Studies of ring chromosome behavior are of im- portance in cytogenetic research because of their application to such problems as chromosome replication and recombination. This paper describes a stable ring chromosome in maize.A chromosome 9 ring with the dominant markers C and
Wx
was produced by X-irradiation of pollen carrying a normal chromosome complement (Figure 1 ).
This ring showed the typical unstable behavior giving rise to cytologically ob- served dicentric rings at anaphase, and in the proper genotype to kernels varie- gated for both C-c and Wz-wx. Two smaller rings were isolated from it, one carrying C but not W z , and the other W z but not C. Both of these were unstable. Variegated kernels carrying the C ring with the recessive c allele on the two normal homologues were planted and crossed to c pollen. All the resultant ears except one gave the expected colorless and C-c variegated kernels. The odd ear gave the low transmission of colored kernels expected with a trisomic fragment ( 16 percent) ; however, all these were uniformly colored. The absence of varie- gation was indicative of a change in the ring chromosome resulting in a stable fragment. This stability has been maintained through four generations of backcrossing.Without an appreciable change in size, the stabilization of a ring chromosome could result as a consequence of one of two events: the opening out of the ring to form a rod chromosome fragment in which sister-strand exchanges do not give rise to dicentrics, or the maintenance of the ring structure with a modifica- tion in its behavior such that dicentric chromosomes are not formed, as appears to be the case with the rings in Drosophila. Although the small size of the ring makes the cytological studies difficult, the configurations observed favor the latter. The stable ring is of about the same size as the small unstable ring from which it arose (Figure 2 ) . At meiosis, the ring is most frequently collapsed upon itself,
I.
FIGURE 1 .-Original unstahle large ring-Pachytene. FIGURE 2.--Small unstahle C ring-Pachytene.
of root tip and glume mitoses and of the meiotic divisions were examined for dicentric double-sized rings. None were found. In all cases the rings were seen to disjoin freely from each other.
The differential behavior of Drosophila and maize ring chromosomes as re- lated to instability is very perplexing. The possibility that this is a reflection of different mechanisms of chromosome replication in plants and animals is in- consistent with LEVAN’S observations of high ring instability in man. The case against this is made even stronger by the finding of the two ring types in maize. Experiments have been conducted indicating that in maize ring behavior is de- pendent on the structure of the chromosome itself and not on the genotype or cytoplasm. Reciprocal crosses were made between plants homozygous for c and w x on the normal chromosomes carrying the stable C ring, and similarly marked plants carrying instead the unstable
Wx
ring. Both these rings are products of the larger ring, which carried both genes. By the use of these markers, those kernels that contained both the stable and unstable rings (the starchy colored kernels) could be recognized. If ring stability were under the genetic control of the nucleus or the cytoplasm, both rings would be expected to show identical behavior when present in the same cells. This was not the case. These kernels were nonvariegated for C and variegated forWx.
It seems clear therefore that ring stability is dependent on the structure of the ring chromosome itself.The variegation studies are based on the classification of the phenotype, whereas the ring is recognized cytologically. To determine whether the color factor being studied is actually carried by the ring chromosome, root tips from colored and colorless sib kernels were sectioned and scored for the presence or absence of the ring chromosome in metaphase. Eighteen kernels of each class were scored. All the colorless were without the ring. Of the colored class, 15 showed the ring. The other three probably represent cases where the ring was present in the endosperm but lost to the embryo.
TABLE I
Frequency of reversion to variegation in endosperms from reciprocal crosses
Cross
Colored Percentage
Colorless Variegated Nonvariegated --- Percentage colored variegation in colored class
c / c / C nng X c/c 8 13,440 26 3117 18.9 0.8
c / c X c/c/C ring 8 11,958 39 1410 10.8 2.7
Total 25,398 65 4527 . . . 1.4
in the female gametophyte. The colored-colorless variegated kernels can result from meiotic or premeiotic reversions or from a reversion in the first division of the gametophyte with an associated loss of the stable ring from the sister nucleus. That the reversions are post meiotic in origin is indicated by the lack of correlation between the endosperm and embryo in the variegated kernels. Difficulty in the detection of the variegation in those endosperms which had received both a stable and unstable ring could account for the low frequency of female reversions.
Eleven variegated kernels were available for crossing in a n attempt to de- termine if the variegation involved a change in the ring back to the unstable con- dition. Two of the variegated kernels received the color factor from the male and the other nine from the female parent. For both variegated kernels from the male, the embryos carried the color factor but in the stable condition. These plants were used as female parents in backcrosses to tester plants, and showed the low transmission of the color factor. One plant gave 108 c : 35 C, and the other 129 c : 28 C : 2 C-c variegated. I n the female, there is a definite correlation between variegation in the endosperm and loss of the sister ring chromatid to the embryo. Of the nine variegated kernels from the female, one showed the color factor in the embryo as well, with low transmission (129 c : 23 C). All the colored seeds were nonvariegated. Seven gave only colorless progeny, indicating the absence of the color gene in the embryo. Because of the dissimilarity between the endo- sperm and embryo in these ten cases it was not possible to determine the cause of the reversion to variegation.
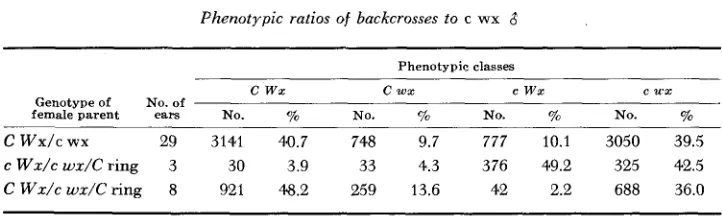
independence, and class 3 an odd distribution resulting from the transfer of c
w x and c W x kernels into the phenotypic C w x and C W x classes respectively
because of the presence of the C ring (Table 2). There is some indication from the data of a non random recovery of the ring among the crossover and non- crossover classes; however, this is still under investigation.
TABLE 2
Phenotypic ratios of backcrosses to c wx 8
Phenotypic classes
c wx c w x e wx e u x
Genotype of No. of
female parent ears No. yo No. 70 NO. To No. %
c
w x / c wx 29 3141 40.7 748 9.7 777 10.1 3050 39.5c W d c w x / C ring 3 30 3.9 33 4.3 376 49.2 325 42.5 c W d c w d C r i n g 8 921 48.2 259 13.6 42 2.2 688 36.0
A single case of reversion to an unstable ring in the embryo was found. The associated endosperm was self-colored, not variegated, again indicating a postme- iotic origin for the reversion. I n a backcross the colored progeny of this plant were all variegated (195 c : 40 C-c variegated). The ring was observed in both root tip prophases and pachynema and showed no detectable morphological differences from the parental stable ring from which it was derived. Double bridges result- ing from dicentric rings have been observed in root tip anaphases. To date no premeiotic reversions giving instability in both the endosperm and embryo have been found, but many more variegated kernels are presently being tested. Insta- bility in the embryo is detected by endosperm variegation in the next generation progeny.
S U M M A R Y
The behavior of a stablJ ring chromosome derived from an unstable ring is described. It is recognized as such by the absence of dicentric double sized rings at anaphase and of variegation for the markers that it carries. The stability is dependent on the structure of the ring itself. Unstable and stable ring chromo- somes retain their individuality even when both are present in the same cells. Apparent postmeiotic reversions to the unstable condition are recovered.
LITERATURE CITED
HINTON, C., 1957 The analysis of rod derivatives of an unstable ring chromosome of Dro- sophila melanogaster. Genetics 42 : 55-65.
LEVAN, A., 1956 Self-perpetuating ring chromosomes i n two human tumors. Hereditas 42: 366-371.
MCCLINTOCK, B., 1938 The production of homozygous deficient tissues with mutant char- acteristics by means of the aberrant mitotic behavior of ring-shaped chromosomes. Genetics 23: 315-376.
The stability of broken ends of chromosomes in Zea mays. Genetics 26: 234-282. 1941
NOVITSKI, E., 1955
SANDLER, L., 1957
SCHWARTZ, D., 1953a
Genetic measures of centromere activity in Drosophila melanogaster. J.
The meiotic behavior of reversed compound ring X chromosomes in Dro-
The behavior of an X-ray-induced ring chromosome in maize. Am. Cellular Comp. Physiol. Suppl. 2. 45: 151-169.
sophila melanogaster. Genetics 42 : 764-782.
Naturalist 87: 19-28.
1953b Evidence for sister-strand crossing over in maize. Genetics 38: 251-260.