1535-9778/06/$08.00⫹0 doi:10.1128/EC.5.3.507–517.2006
Copyright © 2006, American Society for Microbiology. All Rights Reserved.
The RIM101 Pathway Contributes to Yeast Cell Wall Assembly
and Its Function Becomes Essential in the Absence
of Mitogen-Activated Protein Kinase Slt2p
F. Castrejon, A. Gomez, M. Sanz, A. Duran, and C. Roncero*
Instituto de Microbiologia Bioquimica and Departamento de Microbiologia y Genetica, CSIC/Universidad Salamanca, Salamanca, Spain
Received 7 December 2005/Accepted 27 December 2005
The Saccharomyces cerevisiae ynl294c⌬ (rim21⌬) mutant was identified in our lab owing to its moderate
resistance to calcofluor, although it also displayed all of the phenotypic traits associated with its function as the putative sensor (Rim21p) of the RIM101 pathway.rim21⌬also showed moderate hypersensitivity to sodium dodecyl sulfate, caffeine, and zymolyase, and the cell wall compensatory response in this mutant was very poor, as indicated by the almost complete absence of Slt2 phosphorylation and the modest increase in chitin synthesis after calcofluor treatment. However, the cell integrity pathway appeared functional after caffeine treatment or thermal stress. rim21⌬ and rim101⌬ mutant strains shared all of the cell-wall-associated phenotypes, which were reverted by the expression of Rim101-531p, the constitutively active form of this transcription factor. Therefore, the absence of a functional RIM101 pathway leads to cell wall defects.rim21⌬, as well asrim101⌬, was synthetic lethal withslt2⌬, a synthetic defect alleviated by osmotic stabilization of the media. The double mutants grown in osmotically stabilized media were extremely hypersensitive to zymolyase and showed thicker cell walls, with poorly defined mannoprotein layers. In contrast, rim21⌬ rlm1⌬ and
rim101⌬rlm1⌬double mutants were fully viable. Taken together, these results show that the RIM101 pathway
participates directly in cell wall assembly and that it acts in parallel with the protein kinase C pathway (PKC) in this process independently of the transcriptional effect of the compensatory response mediated by this route. In addition, these results provide new experimental evidence of the direct involvement of the PKC signal transduction pathway through the Sltp2 kinase in the construction of yeast cell walls.
Cell walls surround yeast cells acting as an exoskeleton that confers cell shape, while protecting them from harsh environ-ments (8). They are formed by different types of molecules, including structural components, such as glucans and chitin, as well as other molecules, such as different types of mannopro-teins. These components are interconnected, although the na-ture of such interconnections is only partially known (5, 40). Despite the wealth of information collected about yeast cell walls, the specific role of many of the cell-wall-associated pro-teins remains unknown.
The importance of the cell wall in yeast biology is high-lighted by the fact that inhibition of the synthesis of any of its structural components leads to cell death, making the yeast cell wall an attractive target for antifungal therapy (45). Yeast cells contain a dedicated mechanism, the cell integrity or protein kinase C (PKC) signal transduction pathway, which allows the cells to respond to cell wall damage (24). The response signal is executed by means of the transcriptional induction of several genes, mainly mediated by the Rlm1p transcription factor (13, 21). This transcriptional response is known as the compensa-tory response since it compensates for the damage produced in cell walls. Cell wall damage is sensed by a functional family of plasma membrane proteins whose two most important mem-bers are Wsc1p and Mid2p (19, 32, 44). Although their exact roles are not fully understood, they are partially redundant,
and the simultaneous absence of both is lethal (16). After damage has occurred, the signal is transmitted through a ki-nase cascade whose distal mitogen-activated protein (MAP) kinase is Slt2p, which in turn phosphorylates Rlm1p, leading to its activation (for a review, see reference 24). According to its biological function, the PKC route is also constitutively in-duced in most mutants affected in cell wall structure (21). However, the PKC response is also induced by other types of stressful conditions, such as heat shock or caffeine treatment, whose effects cannot be directly related to cell wall structure (10, 27). Besides its role as a defense mechanism, the PKC cascade is also involved in the coordination of cell wall expan-sion during the cell cycle. In view of its different cellular roles, it is not surprising that mutations in the different components of the cascade lead to diverse phenotypes, which have allowed the identification of several cellular targets specific for each component of the cascade. Such targets have recently been reviewed (24) and have been grouped into two very different cellular categories: nuclear and cytoplasmic. At present, only a few targets for Slt2p MAP kinase have been established, in-cluding two transcription factors, a protein phosphatase, and a cell surface Ca2⫹channel. However, genetic evidence has im-plicated Slt2p in the control of several additional hitherto-undefined substrates (24).
Although the role of PKC signaling in cell wall synthesis and assembly is well established, recent advances in our knowledge of signaling transduction processes strongly suggest that most, if not all, yeast signaling pathways also modulate this process. The HOG route (18) acts antagonistically to PKC in the con-struction of the yeast cell wall (14) and the compensatory
* Corresponding author. Mailing address: Departamento de Micro-biologia y Genetica, CSIC/Universidad de Salamanca, Edificio Departa-mental, R-219, Avda. Campo Charro s/n, 37007-Salamanca, Spain. Phone: 34-923-294733. Fax: 34-923-224876. E-mail: [email protected].
507
on September 8, 2020 by guest
http://ec.asm.org/
response depends partially on the induction of the transcrip-tion factor Crz1p by the calcineurin pathway (13, 21). In ad-dition, it has been known for some time that the sporulation signaling cascade directs spore cell wall synthesis in a timely fashion (7, 20) and that proper mating depends on the PKC response (33). Such relationships are not unexpected since the cell wall is in direct contact with the environment and therefore constitutes a major barrier against any environmental change. In addition to the signaling pathways described above, fungi, including yeasts, contain a dedicated signal transduction path-way that is activated at alkaline pH: the so-called RIM101 pathway inSaccharomyces cerevisiae(31). RIM101 signaling, as well as all fungal alkaline pH systems, is atypical since it is not based on a MAP kinase cascade but rather on proteolytic pro-cessing of a transcription factor (30, 31), known as Rim101p inS. cerevisiae(25). Although the RIM101 pathway was described inS. cerevisiaeseveral years ago in the context of its involvement in sporulation (28), most of our knowledge about this route comes from studies on filamentous fungi orCandida albicans. The sig-naling machinery appears to be evolutionarily conserved in most fungi, and the most recent hypothesis suggests that the transcrip-tion factor is proteolytically processed at the late-endosome mem-branes (for a review, see reference 30). Nevertheless, how the sensors of this route transmit the signal remains unknown. InS. cerevisiaethe RIM cascade is formed by the products of theRIM8
(25), RIM9(9), RIM13 (12), RIM20 (47), RIM21 (2, 42), and
DFG16(2) genes in addition to the transcription factor Rim101p (25), although the function of some of them is not well estab-lished.
The yeast transcriptional response mediated by Rim101p has been characterized by using DNA array-based technology (22, 23, 39) and has provided some unexpected answers: Rim101p functions as a repressor in vivo by downregulating the expression of several genes, including the transcription factorsNRG1and SMP1(22), which themselves act as tran-scriptional repressors. Therefore, the absence of Rim101p pro-duces both an increase and a decrease in the expression of different genes. How Rim101p exerts its function is not com-pletely understood, and the regulatory response differs be-tweenS. cerevisiae(22) andC. albicans(26).
Direct experimental evidence linking the RIM101 signaling pathway and cell wall construction in yeast has not yet been obtained, but a very recent report indicates that some rim
mutants show synthetic lethality with some cell wall mutations (41). In addition, some cell-wall-affected mutants display re-duced growth at alkaline pH (38). Both observations, although not conclusive, suggest a possible relationship between the RIM101 pathway and S. cerevisiaecell wall assembly. More importantly, the expression ofPHR1andPHR2, as well as that of many other cell-wall-related genes, are pH regulated by the RIM101 pathway inC. albicans(26). CaPhr1/2p are the func-tional homologues of ScGas1p andAspergillus fumigatusGel1p proteins, two well-characterized glycosyl-transferase enzymes with essential functions in the construction of fungal cell walls (29). Unfortunately, the fact that theGAS1gene is not under the control of the RIM101 pathway either inS. cerevisiaeor inA. fumigatus does not support the relationship between RIM101 and cell wall assembly.
We present here direct experimental evidence linking the RIM101 and PKC pathways in the construction ofS. cerevisiae
cell walls. Both routes appear to act in parallel in the process, and the genetic interactions between them could open new alternatives for the therapeutic inhibition of fungal growth.
MATERIALS AND METHODS
Strains, plasmids, and yeast genetic methods.Standard procedures were used for yeast genetic manipulations (35) and DNA manipulations (36). Plasmid pWL86, a pRS314 vector containing aRIM101gene version truncated at the codon 531 (Rim101-531p), was kindly provided by A. P. Mitchell (25). TheS. cerevisiaestrains used in the present study and their origins are listed in Table 1. Single mutants were made by the one-step gene replacement technique, while double mutants were obtained after tetrad dissection of the double heterozygous mutants obtained after conjugation of the appropriate single mutant strains.
SinglekanMX4mutants were obtained from EUROSCARF, and the deletion cassettes were amplified from the genomic DNA of these strains by PCR with primers placed 200 nucleotides up- and downstream from each open reading frame. Themid2::kanMX4deletion cassette was kindly provided by H. Bussey. The deletion cassettes were transformed into the appropriate strains, and correct transformants were verified by PCR and phenotypic analysis. Thechs3::URA3,
fks1::URA3,pbs2::LEU2,rlm1::LEU2, andslt2::URA3deletion cassettes have been described elsewhere.
Therim21::URA3deletion cassette was constructed as follows. P1 and P2 DNA fragments about 250 bp up- and downstream from theRIM21open reading frame, respectively, were PCR amplified by using oligonucleotides 5⬘-GTACTACAGATT CGGTGTCC-3⬘and 5⬘-CTAAGCTTCTGCATATACTGCCG-3⬘(P1) and oligo-nucleotides 5⬘-TCAAGCTTCCAAACAGAGAAAGGCCA-3⬘and 5⬘-TATGGCC TAGGTCGC-3⬘ (P2). Both fragments were cloned into the pGEMT vector (Promega), and the resulting plasmid was linearized with HindIII, where a HindIII-fragment containing theURA3was inserted between the P1 and P2 fragments (pFMC1). The fragment containing the deletion cassette was transformed into different strains as described above to obtain therim21⌬::URA3strains.
Culture conditions.YEPD (1% yeast extract, 2% peptone, and 2% glucose) broth or agar was the complete medium used for growing yeast strains. Synthetic minimal medium (SC; 0. 7% yeast nitrogen base without amino acids and 2% glucose) was routinely used after supplementation with appropriate amino acids and nucleic acid bases.
Growth in the presence of NaCl, sorbitol, caffeine, and sodium dodecyl sulfate (SDS) was always carried out on YEPD medium supplemented with different concentrations of the compounds, while resistance to calcofluor was assayed on SC medium buffered with 50 mM potassium hydrogen phathalate (pH 6.2). Sensitivity to alkaline pH was tested on YEPD medium, buffered with 100 mM HEPES adjusted with NaOH to the desired pH. For sensitivity tests, cells were
TABLE 1. Yeast strains
Strain Relevent genotype Source
W303 1A MATacan1-100 ade2-1 his3-11,15 Ieu2-3,12 trp1-1 ura3-1
Lab collection
W303 1B MAT␣can1-100 ade2-1 his3-11,15 Ieu2-3,12 trp1-1 ura3-1
Lab collection
FCM25 W303MATarim21::URA3 This study FCM28 W303MATarim21::kanMX4 This study FCM314 W303MATarim101::kanMX4 This study FCM157 W303MAT␣chs3::URA3 Lab collection FCM111 W303MAT␣fks1::URA3 Lab collection FCM182 W303MAT␣pbs2::LEU2 Lab collection FCM165 W303MAT␣mid2::kanMX4 This study FCM319 W303MAT␣wsc1::kanMX4 This study FCM110 W303MAT␣slt2::URA3 This study FCM323 W303MAT␣rlm1::LEU2 Lab collection FCM159 W303rim21::kanMX4 chs3::URA3 This study FCM153 W303rim21::kanMX4 fks1::URA3 This study FCM161 W303rim21::kanMX4 pbs2::LEU2 This study FCM167 W303rim21::URA3 mid2::kanMX4 This study FCM382 W303rim21::URA3 wsc1::kanMX4 This study FCM236 W303rim21::kanMX4 slt2::URA3 This study FCM386 W303rim21::kanMX4 rlm1::LEU2 This study FCM328 W303rim101::kanMX4 slt2::URA3 This study FCM388 W303rim101::kanMX4 rlm1::LEU2 This study BY4741 MATahis3⌬1 leu2⌬0 met15⌬0 ura3⌬0 Euroscarff FCM384 BY4741smp1::kanMX4 Euroscarff FCM399 BY4741rim21::URA3 This study FCM400 BY4741smp1::kanMX4 rim21::URA3 This study
on September 8, 2020 by guest
http://ec.asm.org/
grown overnight in the appropriate medium and diluted to an optical density at 600 nm (OD600) of approximately 0.2 to 0.3. Then, 5l of 1/10 serial diluted
samples were spotted onto the indicated media. Growth was recorded after 48 h of incubation at 28°C.
The calcofluor effect was also tested in cells grown for 3 h in liquid YEPD supplemented with 0.075 mg of calcofluor/ml. The effect of this drug was tested by observation of septa enlargement generated under a Leica DRM500 fluores-cence microscope or by the determination of the cell wall chitin levels. Fluores-cence images were acquired with a Sensys digital camera and further processed with the Adobe Photoshop 5.5 software. All images were exposed and processed identically for accurate analysis.
Preparation of yeast extracts and immunoblot analyses.Yeast cells were grown overnight at 24°C to mid-log phase in the appropriate medium, diluted to an OD600of 0.2, and grown for 1 h. Then, cells were treated with the required
compound or shifted to 39°C for the indicated times. Cells were collected on ice-chilled water in a refrigerated centrifuge, transferred to a microcentrifuge tube, immediately pelleted, and frozen in dry ice. Cells were lysed in 120l of cold lysis buffer (50 mM Tris-HCI [pH 8], 1% Triton X-100, 150 mM NaCl, 10 mM sodium pyrophosphate, 5 mM EDTA, 5 mM EGTA, 1 mM sodium or-thovanadate, 1 mM phenylmethylsulfonyl fluoride, and the protease inhibitors TPCK [tolylsulfonyl phenylalanyl chloromethyl ketone], tosyllysine chloromethyl ketone, leupeptin, pepstatin A, antipain, and aprotinin [each at 25g/ml]) by vigorous shaking with 0.45-mm glass beads in a Fast Prep cell breaker (Bio 101). Cell extracts were separated from the glass beads and cell debris and further clarified by a 16,000⫻gspin for 15 min at 4°C. The protein concentration of the extract was determined at 280 nm, and samples were diluted in 2⫻loading buffer. Samples containing 100 mg of proteins were boiled for 5 min and frac-tionated in 10% SDS-polyacrylamide gels.
Western blots were carried out as described previously (27) by using an anti-phospho-p44/42 MAP kinase antibody (New England Biolabs) at a 1:500 dilution or an anti-Mpk1p (yN-19) goat polyclonal antibody to detect dually phosphorylated Slt2p or total Slt2p, respectively. Blots were developed by using the corresponding horseradish peroxidase-conjugated secondary antibody to-gether with the ECL (Amersham Biosciences) detection system.
Zymolyase sensitivity assays.Yeast sensitivity to zymolyase was assayed as growth inhibition, as described previously (1). Briefly, cells were pregrown, di-luted to an OD600of 0.005, and further incubated for 24 h at 28°C in YEPD or
1 M sorbitol-supplemented YEPD in the presence of different Zymolyase 100T (Seikagaku Corp.) concentrations. Growth was then determined by measure-ment of the OD600of each culture. The data are expressed as the percentage of
growth relative to the culture grown without zymolyase.
Chitin determinations.Chitin measurements were performed with chitinase fromSerratia marcescens (Sigma-Aldrich) and colorimetric determination of GlcNAc as described previously (43). Total amounts of chitin are expressed as millimoles of GlcNAc in 100 mg of cells (wet weight).
Electron microscopy.Transmission electron microscopy (TEM) was performed essentially as described previously (46). Briefly,S. cerevisiaecultures were grown in 1 M sorbitol-supplemented YEPD to an OD600of 0.5 to 1.0. Then, 12.5 OD600units
of cells were collected by centrifugation, rapidly washed, and fixed in 2% potassium permanganate for 1 h at room temperature. Excess potassium permanganate was removed by exhaustive washing, and the stained cells were finally dehydrated by incubation in increasing concentrations of ethanol. Samples were processed in the embedding medium (Spurr Resin Embedding Kit; TAAB) by successive 2-h incu-bations in 1:1 and 1:3 ethanol-embedding medium mixtures and, finally, in fresh embedding medium. Cells were concentrated in 500l of resin, and samples were allowed to polymerize overnight at 60°C. Ultrathin sections were obtained by using an LKB Ultratome III microtome, and samples were visualized under a Zeiss EM900 transmission electron microscope. Images were processed with the Adobe Photoshop software, preserving relative magnifications. Cell wall widths were deter-mined in several cells of each strain, and the individual value for each cell is the average of eight different measurements along its surface.
RESULTS
Identification of YNL294 as a cell wall-related gene. The participation of our laboratory in the EUROFAN project al-lowed us the possibility of screening multiple nonessential mu-tants for cell-wall-associated phenotypes. Among them, the
ynl294⌬mutant showed calcofluor resistance and sensitivity to zymolyase, two phenotypic traits also associated with the lack of CSIII activity (17). However,ynl294⌬appeared to contain
normal amounts of CSIII, which prompted us to investigate the reason for the calcofluor resistance in this mutant.
During the course of the present study the Ynl294 protein was shown to have modest but significant identity withA. nidulans
PalHp (42). The functional characterization of this gene placed it in the ambient pH signaling pathway, known as the RIM101 pathway inS. cerevisiae. This gene was namedRIM21in all major databases, and it has been assumed to be the functional homo-logue ofA. nidulans PalHp. Therefore, we shall refer to it as
RIM21/Rim21p throughout the present study.
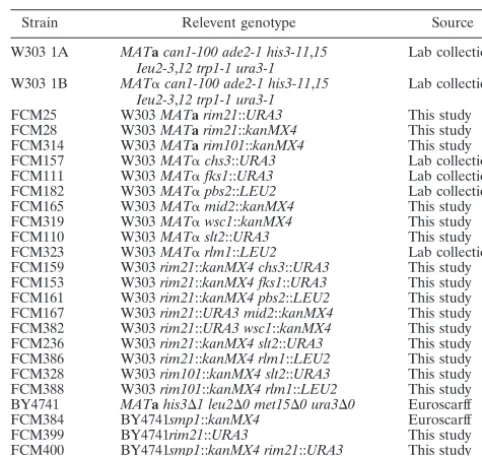
rim21⌬ cells were resistant to moderate concentrations of calcofluor (Fig. 1A) and, in accordance with the reported CSIII level (17), they contained similar amounts of chitin as the wild type (Table 2). After calcofluor treatment, chitin was localized at the septum region as in the wild type, but the intensity of the staining was significantly reduced (Fig. 1B), a clear indication that chitin levels should be reduced in these conditions. This turned out to be the case, asrim21⌬cells contained only 44% of the chitin contained in wild-type cells after calcofluor treat-ment (Table 2). This reduction was due to the lower increase in chitin biosynthesis occurring in the mutant after calcofluor treatment. Interestingly, this situation was very similar to that observed for theslt2⌬ andrlm1⌬ mutants in terms of chitin synthesis (Table 2) or calcofluor staining (not shown). Since bothslt2⌬andrlm1⌬mutants are defective in the cell integrity signaling (PKC) response, it was necessary to address the phe-notypic traits associated with the absence of this route.
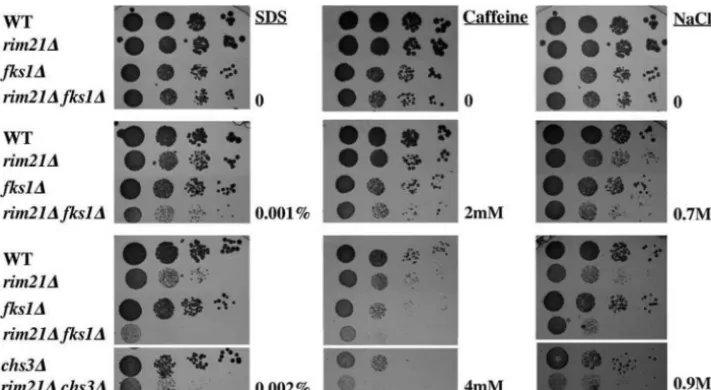
rim21⌬ growth was significantly reduced at pH above 7.6 (Fig. 1C). In addition, the mutant grew poorly in 0.9 M NaCl (Fig. 1C), but no significant defect in sporulation was observed. These phenotypes clearly agree with the proposed model of Rim21p as part of the alkaline pH signaling response (see Discussion for further analysis).
rim21⌬ growth was also reduced in the presence of 4 mM caffeine and 0.002% SDS (Fig. 1C), but it was not affected by 1.5 M sorbitol (Fig. 1C) or high temperature (not shown). Appar-ently, this mutant is moderately hypersensitive to SDS and caf-feine but not to high osmolarity. These phenotypes have been traditionally associated with defects in the cell wall structure.
Therim21⌬mutant fails to activate the PKC response after
cell wall damage.The results obtained showed that, although to a different extent, therim21⌬andslt2⌬mutants shared multiple phenotypes, pointing to Rim21p as a part of the cell integrity signaling response (PKC). In order to confirm this point, we determined the degree of the PKC response in therim21⌬mutant after calcofluor treatment. In the wild-type cells, calcofluor in-duced a rapid and long-lasting phosphorylation of Slt2p in re-sponse to the cell wall damage produced (Fig. 2A). In contrast, Slt2p phosphorylation was virtually absent in therim21⌬mutant (Fig. 2A) under similar growth conditions. The absence of this response is not a consequence of secondary mutations in this strain since it is also absent in an independent deletion strain (FMC28) and in the original FY1679⌬ynl294::kanMX4mutant (data not shown). In conclusion, deletion of RIM21 abolished PKC response after calcofluor treatment. Thefks1⌬mutant has an altered cell wall synthesis, and hence it shows a constitutive activation of the PKC pathway as determined by the degree of Slt2p phosphorylation (Fig. 2B) and the increase in chitin synthe-sis (Table 2). However, Slt2p phosphorylation was not observed in the doublefks1⌬ rim21⌬mutant and thus chitin synthesis was
on September 8, 2020 by guest
http://ec.asm.org/
reduced (Table 2). Taken together, these results indicated that the PKC response was not elicited in therim21⌬mutant after cell wall damage produced by two completely different mechanisms. In contrast, Slt2p phosphorylation was induced in the rim21⌬
mutant by incubation at high temperatures or after caffeine treat-ment (Fig. 2C), another two stressful situations in which the PKC response is activated (Fig. 2C) (27). Apparently, Rim21p is spe-cifically involved in the activation of the PKC response after direct cell wall damage.
In order to further confirm the relationship between Rim21p and cell wall construction, we analyzed the hypothetical
syn-FIG. 1. Phenotypic characterization of therim21⌬mutant. (A) Resistance to calcofluor on SC medium. Plates were incubated for 48 h at 28°C. (B) Chitin distribution after calcofluor (0.075 mg/ml) treatment for 3 h. Note the lower intensity of the staining in the mutant. (C) Comparative growth of wild-type andrim21⌬strains on the indicated media. 1/10 serial dilutions of overnight cultures were spotted and incubated for 48 h at 28°C. WT, wild type.
FIG. 2. Activation of the PKC signaling pathway under different conditions. (A) Time course of Slt2p phosphorylation after calcofluor treatment. Samples were collected at the indicated times after the addition of calcofluor. The levels of total Slt2p are shown as a loading control. (B) Slt2p phosphorylation in wild-type andfks1⌬strains grow-ing logarithmically or after calcofluor treatment. (C) Comparative levels of Slt2p phosphorylation in wild-type andrim21⌬strains after caffeine, calcofluor, or thermal stress. WT, wild type.
TABLE 2. Chitin levels in selected strains
Strain
Chitin level (mmol of GlcNAc/100 mg
of cells)a(%) Increase
(fold)b Calcofluor
resistancec
Control Calcofluor
WTd
mutant 0.24 (100) 1.15 (100) 4.8 ⫹
rim21⌬mutant 0.24 (100) 0.51 (44.3) 2.1 ⫹⫹
rim101⌬mutant 0.30 (125) 0.68 (59.1) 2.3 ⫹⫹
slt2⌬mutant 0.26 (108) 0.60 (52.2) 2.3 ⫺
rlm1⌬mutant 0.32 (133) 0.78 (67.8) 2.4 ⫹⫹
fks1⌬mutant 1.11 (463) 1.01 (87.8) NAe ⫺
rim21⌬fks1⌬mutant 0.84 (350) 0.85 (73.9) NA NA
aChitin was determined in cells growing in YEPD or in YEPD supplemented
with 0.075 mg/ml calcofluor media, as indicated. Detailed protocols are described in Materials and Methods.
bThat is, the relative increase in chitin levels after calcofluor treatment. cResistance to calcofluor is with reference to the WT (⫹). Therefore, “⫺”
stands for hypersensitive and “⫹⫹” represents moderately resistant. On this scale, mutants lacking chitin would be “⫹⫹⫹⫹.”
dWT, wild type. eNA, not applicable.
on September 8, 2020 by guest
http://ec.asm.org/
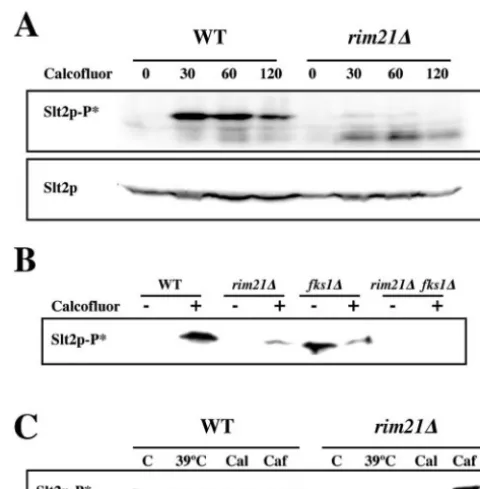
thetic lethality betweenrim21⌬and several other mutants pre-viously shown to have altered cell walls.chs3⌬rim21⌬,pbs2⌬ rim21⌬,fks1⌬rim21⌬,mid2⌬rim21⌬, andwsc1⌬rim21⌬ dou-ble mutants were viadou-ble and did not show any apparent defect under normal growth conditions (data not shown). However, we were unable to obtain the double slt2⌬ rim21⌬ mutant, suggesting a synthetic lethal interaction between both muta-tions (see below). Further characterization of the double mu-tants indicated that the phenotypic defects detected were ad-ditive (Fig. 3 and data not shown). However, thefks1⌬rim21⌬
mutant showed an extreme hypersensitivity to SDS (Fig. 3) that could not be explained by the additive effect of both mutations.
We have previously stated thatslt2⌬rim21⌬double mutants appeared to be nonviable. In order to confirm this lethality, we performed a statistical analysis ofrim21⌬ and slt2⌬ segrega-tion. As expected, noslt2⌬ rim21⌬double mutants were ob-tained after the micromanipulation of 34 tetrads (Table 3), albeit the normal segregation of the two characters occurred individually, a clear indication of the synthetic lethality be-tween both mutations. In order to analyze whether or not such lethality might be directly related to a cell wall defect, we performed the segregation analysis on plates supplemented with 1 M sorbitol. As shown in Table 3, from the spores analyzed we were able to recover a significant number of dou-ble mutants, which accounted for ca. 60% of those expected based on the appearance of single mutations. Apparently,
rim21⌬slt2⌬synthetic lethality can be suppressed by osmotic stabilization of the growth media, suggesting a severe defect in cell wall assembly in this double mutant.
The RIM pathway is involved in cell wall construction.All of the results presented thus far link the phenotypes of the
rim21⌬mutation to the construction of the yeast cell wall through the participation of Rim21p in the cell integrity signaling response. In addition, according to its structure and functional similarity to AnPalHp (2, 42), Rim21p ap-pears to be a membrane protein that acts as a sensor. We were therefore prompted to place Rim21p as an additional
sensor of the PKC pathway. If this were the case, we reasoned that we would not find a relationship between RIM101 and PKC pathways but rather an involvement of Rim21p in both routes. In order to test this point, we characterized the cell-wall-associated phenotypes of therim101⌬mutant, which lacks the transcription factor associated with the RIM101 response. Therim101⌬mutant was partially resistant to calcofluor and hypersensitive to SDS and caffeine (results not shown). It con-tained normal amounts of chitin, but calcofluor treatment in-creased chitin synthesis only by a factor of 2 (Table 2), in a way similar to what occurred for therim21⌬mutant. In addition, we were unable to obtain the doublerim101⌬slt2⌬mutant in the W303 genetic background (Table 3), a clear indication of
FIG. 3. Phenotypic analysis of selected double mutants. Early logarithmic growing cells were spotted onto the indicated media at 1/10 serial dilutions. Growth was scored after 48 h at 28°C. Note the extreme hypersensitivity to SDS of the doublefks1⌬rim21⌬mutant. WT, wild type.
TABLE 3. Spore viabilityain different crosses
Cross Medium
Spore viability with genotypea:
SLT2 RIM21
(101)
slt2⌬ RIM21
(101)
SLT2 rim21⌬
(101)
slt2 rim21⌬
(101)
slt2⌬ ⫻rim21⌬ YEPD 27 30 28 0
YEPD⫹S 28 29 30 17
slt2⌬ ⫻rim101⌬ YEPD 29 49 44 0
YEPD⫹S 28 47 46 15
slt2⌬ ⫻rim21⌬
(pWL86)b
YEPD 27 28 26 12
rlm1⌬ ⫻rim21(101)⌬ YEPD 4 (6) 4 (10) 6 (8) 8 (7)
aThe spore viability is the number of spores of each genotype recovered from
each medium after tetrad dissection. The relevant genotype of the diploid is indicated. The spore genotype relevant for the study is indicated, but note the alternativeRIM101/rim101⌬loci in the corresponding cross in parentheses.
bPlasmid shuffling in viable spores of theslt2⌬ ⫻rim21⌬(pWL86) cross was
as follow (genotype, plasmid loss):SLT2 RIM21, 46.6% (n⫽88);slt2⌬RIM21, 80.2% (n⫽86);SLT2 rim21⌬, 52.3% (n⫽88); andslt2⌬rim21⌬, 0.0% (n⫽
434). Several viable spores of each genotype were grown for 30 generations in nonselective medium and plated, and isolated colonies were tested for the pro-totrophy associated with the plasmid. The results are expressed as the percentage of cells that have lost the plasmid. The total number of colonies tested is indicated for each strain.
on September 8, 2020 by guest
http://ec.asm.org/
the synthetic lethality between both mutations. This lethality could also be partially overcome by osmotic stabilization of the media (Table 3). The similarity betweenrim21⌬andrim101⌬
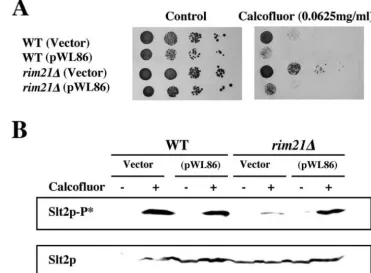
phenotypes strongly supports the relationship between the RIM101 and PKC routes rather than a direct role of Rim21p in the PKC cascade. In order to confirm this hypothesis, we analyzed the complementation behavior of a plasmid contain-ing the C-terminal deletion of Rim101p which produces its constitutive activation (25) and hence induction of the RIM pathway. The introduction of plasmid pWL86 (Rim101-531p) in the rim21⌬ mutant restored alkaline pH growth (not shown), calcofluor sensitivity (Fig. 4A), and the PKC response after calcofluor treatment (Fig. 4B), without having a direct effect per se on the induction of the PKC response (Fig. 4B). In clear accordance with these results, Rim101-531p also pro-moted the growth of the otherwise lethalrim21⌬slt2⌬mutant. However, the suppression of this lethality was only partial and quantitatively very similar to that obtained by osmotic stabili-zation (Table 3). This experiment also allowed us to reconfirm the lethality of theslt2⌬rim21⌬double mutant. It was able to grow containing the pWL86 plasmid, but we were unable to induce plasmid shuffling in this strain even after 30 generations of growth in nonselective media, where other strains lose plas-mids with a frequency close to 50% (Table 3).
In conclusion, simultaneous inactivation of the RIM101 and PKC signaling pathways leads to nonviable cells. However, it should be noticed that neither therim21⌬orrim101⌬ muta-tions showed synthetic lethality with therlm1⌬mutant (Table 3) and that the double mutants grew normally. This mutant
lacks the only transcription factor that has been experimentally implicated in the PKC-induced transcriptional response, thus pointing to additional levels of control of cell wall assembly through the PKC route.
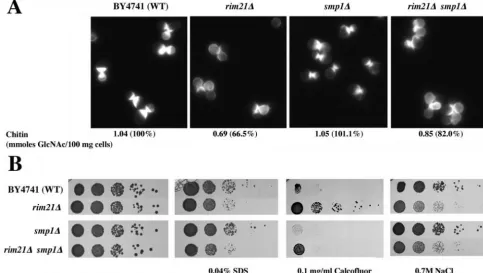
Deletion of the Smp1p transcriptional repressor has minor effects on cell wall-associatedrim21⌬defects.Recent advances in the regulatory network of RIM101 signaling inS. cerevisiae
have pointed to an important role of two transcriptional re-pressors, Nrg1p and Smp1p, in the process (22). Although there is no experimental evidence implicating either of them in cell wall assembly (http://www.yeastgenome.org/), Smp1p is the only known homologue of the Rlm1p transcription factor inS. cerevisiae(11). It was therefore tempting to associate the role of RIM signaling in cell wall assembly with a Smp1p deregu-lation. In order to test this possibility, we characterized the
smp1⌬andsmp1⌬rim21⌬phenotypes. Thesmp1⌬strains did not show any cell-wall-associated phenotype (Fig. 5) but, more interestingly, deletion of theSMP1gene did not suppress the hypersensitivity of arim21⌬mutant to SDS or NaCl (Fig. 5B). Double mutants were still moderately resistant to calcofluor (Fig. 5B), and the effect of this drug was lower than in the wild type, as determined by calcofluor staining (Fig. 5A). However, its resistance to calcofluor seemed to be lower than that of the single rim21⌬. In addition, after calcofluor treatment the amount of chitin was significantly higher than inrim21⌬but still lower than in the wild type (Fig. 5A). These results clearly ruled out the derepression ofSMP1as the only factor linking the RIM101 pathway with cell wall assembly, suggesting a
FIG. 4. Effect of the expression of the constitutive form of Rim101p. Wild-type andrim21⌬strains were transformed with pWL86 (Rim101-531p) and characterized phenotypically. (A) Resistance to calcofluor on SC medium. (B) Slt2p phosphorylation before (⫺) and after (⫹) calcofluor treatment in the indicated strains. Total Slt2p levels are shown in the lower panel. WT, wild type.
on September 8, 2020 by guest
http://ec.asm.org/
more complex mechanism in which derepression of SMP1
could be partially involved.
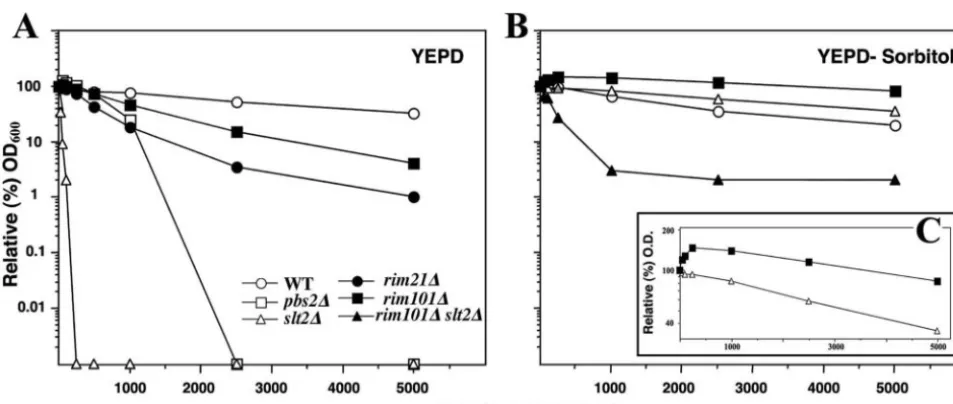
Double RIM and PKC mutants can only grow in osmotically stabilized media because of severe cell wall defects.In a fur-ther attempt to understand the role of the RIM101 pathway in cell wall construction, we analyzed the behavior of the double mutants isolated. As previously stated,slt2⌬rim21⌬orslt2⌬ rim101⌬double mutants were only isolated on YEPD media supplemented with 1 M sorbitol. When these strains were transferred to plain YEPD media, growth stopped and almost no growth was detected on solid (Fig. 6A) or liquid (not shown) medium. Although growth on sorbitol-supplemented solid medium did not indicate any apparent growth defect of
slt2⌬rim21⌬/101double mutants, after inoculation of cultures in liquid mediaslt2⌬ rim21⌬ growth was significantly slower (Fig. 6B). The generation time of the double mutant was 222 min, approximately twice as long as any of the singlerim21⌬or
slt2⌬mutants, with 112 and 110 min, respectively.
The synthetic lethality of theslt2⌬rim21⌬/101mutants was strain dependent since these double mutants in the FY1016 genetic background were able to grow on plain YEPD. How-ever, growth was very slow at 28°C, and the strain was essen-tially thermosensitive at 37°C (not shown), confirming the syn-thetic interaction between the RIM101 and PKC pathways in two different genetic backgrounds.
In order to discover whether this genetic interaction was re-lated to cell-wall-associated defects, as expected from phenotypic characterization of the individual mutants, we carried out a sen-sitivity test to zymolyase, an enzymatic cocktail that acts on fungal
cell walls (see Materials and Methods for the specific protocol). In YEPD medium,slt2⌬cells were extremely hypersensitive to zy-molyase, and these cells did not grow when zymolyase concentra-tions were as low as 25 U (Fig. 7A).pbs2⌬cells, used as control, showed a moderate hypersensitivity to zymolyase. The rim21⌬
andrim101⌬single mutants showed a modest but reproducible hypersensitivity to the enzyme since their growth was reduced in the presence of zymolyase, although they were still able to grow substantially even at the highest concentration used (Fig. 7A). Characterization of the doublerim101⌬slt2⌬mutant was neces-sarily carried out on YEPD with 1 M sorbitol. In this medium, resistance to zymolyase was significantly improved in all of the strains tested and theslt2⌬single mutant became as resistant to zymolyase as the wild type (Fig. 7B). Interestingly, rim101⌬
growth was increased reproducibly at low concentrations of zy-molyase (Fig. 7B and C). However, the doublerim101⌬ slt2⌬
mutant was very hypersensitive to zymolyase compared to the single mutants (Fig. 7B). Similar results were obtained for the
rim21⌬slt2⌬mutant (data not shown).
Further characterization of these mutants was performed by TEM of cells grown in 1 M sorbitol. The overall aspect of the double mutant cells was similar to those of the single mutants and the controls (Fig. 8A), but closer scrutiny (Fig. 8B) re-vealed some differences in the double mutant. The rim21⌬ slt2⌬cell walls were 0.217⫾0.006m in width, significantly (50%) wider than those of the wild-type (0.149⫾0.01m),
rim21⌬(0.143⫾0.009m), orslt2⌬(0.136⫾0.008m) cells. In addition, the darker layers flanking the inner core of the cell
FIG. 5. Phenotypic characterization of thesmp1⌬mutants. (A) Calcofluor staining of the indicated mutants and chitin levels after calcofluor treatment measured as described in Materials and Methods. In parentheses, the relative chitin levels are indicated compared to the wild type. All pictures show identical exposure and processing times. (B) Sensitivity of the indicated strains to different drugs. WT, wild type.
on September 8, 2020 by guest
http://ec.asm.org/
wall appeared less defined, suggesting an altered assembly of the cell wall structure.
DISCUSSION
Ynl294/Rim21p forms a functional part of the alkaline pH signaling pathway but also participates in cell wall construc-tion.Yeast cells respond to damage to their cell walls almost exclusively through the induction of the PKC pathway, which
triggers a transcriptional response mediated by the Rlm1p transcription factor (13, 21). However, recent advances in our knowledge of the mechanisms of cell wall synthesis and assem-bly point to multiple inputs to the regulation of this process (for a review, see reference 24). Therefore, it is not surprising to find multiple mutations conferring cell-wall-associated de-fects through the use of massive screenings. In one of these screenings, partially carried out in our lab, the resistance of the
FIG. 6. Growth of the indicated strains in different media. (A) Complete tetrads grown on YEPD–1 M sorbitol were transferred in parallel to YEPD and YEPD–1 M sorbitol and incubated for 3 days at 28°C. Note the minor growth of the double mutants on plain YEPD. (B) Growth curve of the indicated strains. Logarithmically growing cells in YEPD–1 M sorbitol were diluted in the same medium to an OD600of 0.025, and growth
was monitored for 12 h. Note the slower growth of the doublerim21⌬slt2⌬mutant. WT, wild type.
FIG. 7. Sensitivity of different strains to zymolyase. (A) Growth of indicated strains in increasing concentrations of zymolyase in YEPD media. The data are expressed as percentages of growth compared to identical cultures grown without zymolyase. See Materials and Methods for the detailed protocol. A logarithmic scale is used. (B) Same experiment as in panel A, but the cells were grown on YEPD–1 M sorbitol. Note the extreme growth reduction of the double mutantrim101⌬slt2⌬above 1,000 mU and the minor effect of zymolyase in this medium on the rest of strains. (C) Higher magnification of panel B, comparing the growth of therim101⌬mutant and the wild type. Note the improved growth of the rim101⌬mutant in the presence of zymolyase. WT, wild type.
on September 8, 2020 by guest
http://ec.asm.org/
ynl294⌬ mutant to calcofluor was uncovered (17). However, this mutant contained normal levels of CSIII. Previously, this combination of phenotypes had been only found in different mutants of the HOG pathway (14). It was therefore tempting to associate Ynl294p with a signaling cascade involved in cell wall assembly.
During the course of this work, Ynl294p was named Rim21p as part of the alkaline pH signaling response (RIM101) inS. cerevisiae(2, 42). Our results confirmed this participation since
rim21⌬mutants grew poorly at pH 7.6 and showed a significant hypersensitivity to NaCl (Fig. 1), both phenotypes expected from the alteration of the regulatory network associated with the RIM101 response. In addition, the ⌬rim21 mutant also showed a strong defect in mating (data not shown), whose characterization will be presented elsewhere. Interestingly, our results also highlight the cell-wall-associated defects displayed by therim21⌬mutant. Therefore, our main goal was to know whether Rim21p, or alternatively the RIM101 pathway, was involved in cell wall assembly.
TheS. cerevisiaeRIM101 pathway acts on cell wall structure.
The many cell-wall-associated phenotypes of therimmutants described here can be simply explained in terms of the absence of a functional PKC response after cell wall damage (Fig. 2A and B), even though PKC is activated by other stimuli (Fig. 2C). However, if this model is correct then therim21⌬mutant should have a synthetic lethality pattern similar to that of the
slt2⌬ mutant, which is not the case:chs3⌬ rim21⌬ orfks1⌬ rim21⌬mutants were viable and, more importantly, theslt2⌬ rim21⌬double mutant turned out to be lethal. The synthetic lethality was relieved by osmotic stabilization of the media, suggesting that Rim21p and Slt2p act in parallel in the con-struction of the yeast cell wall. These results also argue against a role of Rim21p as a sensor in the PKC cascade, although is
still possible that it could be involved in the process as part of another signaling cascade, as could be speculated from its biological role as a PM sensor (2, 42).
In order to test this possibility, we refocused our attention on Rim101p, the transcription factor involved in the RIM101 response. rim101⌬ cells showed essentially the same pheno-types asrim21⌬cells. In addition, therim101⌬andslt2⌬ mu-tations were also synthetically lethal. More important, the phe-notypes shown by the rim21⌬ mutant were reversed by the expression of the constitutively activated form of Rim101p (25) (Fig. 4). This reversal included not only growth at alkaline pH but also calcofluor sensitivity, activation of the PKC response after cell wall damage, and synthetic lethality with the slt2⌬
mutation. These results allow us to confirm that the RIM101 pathway, and not only Rim21p, acts on the formation of yeast cell wall. This relationship has also been recently uncovered in massive synthetic lethality analyses which show thatslt2⌬displays synthetic lethality with therim20⌬,rim13⌬, andrim8⌬mutations (41). Surprisingly, neither therim101⌬nor therim21⌬mutations showed synthetic lethality with theslt2⌬mutation in this screen-ing, a finding very likely due to methodological problems associ-ated with massive screenings.
Once the relationship between RIM101 and the cell wall has been established, the next step would be to determine the exact role of the RIM101 pathway in this process. However, this is very difficult, owing to the complex nature of the cell wall components and their interactions. The mild phenotypes found for the⌬rimmutants, as well as their modest hypersensitivity to zymolyase, suggested minor changes in cell wall structure. These changes very likely affect the cell wall interaction with calcofluor, leading to drug resistance and the virtual absence of the PKC response. These minor defects became lethal in the absence ofslt2⌬, but not in therlm1⌬mutant, which lacks the
FIG. 8. TEM images of the indicated strains grown in YEPD–1 M sorbitol. Cells are presented at two different magnification levels to show details. Note the less-defined outer layer, corresponding to the mannoprotein layer (B). WT, wild type. Bars, 0.4m.
on September 8, 2020 by guest
http://ec.asm.org/
transcriptional response mediated by PKC activation after cell wall damage (13, 21). These results strongly support a model in which the RIM and PKC pathways act in parallel in a group of complementary functions required for proper cell wall assem-bly. The specific functions of these genes are very hard to define because Slt2p appears to have multiple intracellular targets not yet defined (24). One such target is the Ca2⫹ chan-nel Cch1p/Mid2p (4). However, the genetic interaction de-scribed here appears to be independent of Ca2⫹signaling since the absence of the calcineurin proteins was not synthetic lethal withrim21⌬ and neither did overexpression of the constitu-tively active calcineurin regulatory subunit (15) suppress the synthetic lethality of therim21⌬slt2⌬mutant (data not shown). Unfortunately, transcriptional insight into the RIM101 path-way is also very complex. The yeast RIM101 pathpath-way has been implicated in transcriptional repression (3, 22) and transcrip-tional induction (3, 23, 39), but it is also required for the expression of several genes (22). The three categories include genes that potentially participate in the construction of yeast cell walls and hence the three regulatory mechanisms could interact in the production of the phenotypes described. In agreement with this hypothesis, deletion of the transcriptional repressorsmp1⌬only partially suppressed the cell wall-associ-ated phenotypes of therim21⌬mutant. The massive synthetic lethality screening recently reported (41) also argues against a single gene being responsible for the phenomena discussed here since none of the multiple genes regulated by the RIM101 pathway showed synthetic lethality with slt2⌬ except for the
RIMgenes.
Interestingly, several of the genes regulated by the RIM101 pathway inS. cerevisiaecode for cell wall remodeling activities, such asCRH1,CRH2(34), orCTS1(6). It is thus very likely that a combinatorial effect of several remodeling activities would account for the phenotypes found in therim⌬mutants. This hypothetical defect in cell wall remodeling rather than in structural activities would provide a more suitable explanation for the mild structural defects observed in the cell walls of the double mutants.
The functional relationship between the RIM101 and PKC routes is also becoming apparent from other evidence. TheC. albicans PHR1andPHR2genes are regulated by pH (37) and, like other fungal homologues such as theS. cerevisiae GAS1
and theA. fumigatus GEL1, they play essential roles in cell wall construction (29, 37). Although there is no evidence about pH regulation of theGAS1orGEL1genes, thegas1⌬and theslt2⌬
and bck1⌬ mutants have recently been shown to display a moderate hypersensitivity to alkaline pH (38). Therefore, yeast cells appear to require cell wall remodeling for growth at high pH. If this hypothesis is correct, then very likely alkaline pH would trigger the PKC response. This is the case since the growth ofS. cerevisiaeat pH 7.6 induced a transient phosphor-ylation of Slt2p depending on the presence of Rim21p (data not shown). In conclusion, the RIM101 and PKC pathways cooperate in cell wall remodeling after alkaline shock.
The main conclusion from the present study is that this is the first direct experimental evidence assigning a role to the RIM101 pathway in the assembly of theS. cerevisiaecell walls. This rela-tionship should help to define the additional intracellular targets of Slt2p that have remained elusive thus far. To date, virtually all of the signal transduction pathways described in yeasts act on cell
wall assembly, highlighting the dynamic nature of this structure that allows cellular adaptation in response to many different types of environmental changes.
ACKNOWLEDGMENTS
We thank the members of the laboratory of A. Duran for critical comments on the manuscript and N. Skinner for language revision. Special thanks are due to A. P. Mitchell and M. S. Cyert for strains and plasmids and to all the members of the Eurocellwall project for useful comments.
M.S. acknowledges the financial support from the CSIC through the I3P-BPG2003 program. This research was supported by CICYT grant BIO2004-00280 and EU grants QLK3-CT-2000-01537 and LSHB-CT-2004-511952.
REFERENCES
1.Alonso-Monge, R., l. E. Rea, I. Wojda, J. P. Bebelman, W. H. Mager, and M. Siderius.2001. Hyperosmotic stress response and regulation of cell wall integrity inSaccharomyces cerevisiaeshare common functional aspects. Mol. Microbiol.41:717–730.
2.Barwell, K. J., J. H. Boysen, W. Xu, and A. P. Mitchell.2005. Relationship ofDFG16to the Rim101p pH response pathway inSaccharomyces cerevisiae
andCandida albicans. Eucaryot. Cell4:890–899.
3.Bensen, E. S., S. J. Martin, M. Li, J. Berman, and D. A. Davis.2004. Transcriptional profiling inCandida albicansreveals new adaptive responses to extracellular pH and functions for Rim101p. Mol. Microbiol.54:1335– 1351.
4.Bonilla, M., and K. W. Cunningham.2003. Mitogen-activated protein kinase stimulation of Ca2⫹signaling is required for survival of endoplasmic
retic-ulum stress in yeast. Mol. Biol. Cell14:4296–4395.
5.Cabib, E., and A. Duran.2005. Synthase III-dependent chitin is bound to different acceptors depending on location on the cell wall of budding yeast. J. Biol. Chem.280:9170–9179.
6.Cabib, E., S. J. Silverman, and J. A. Shaw.1992. Chitinase and chitin synthase 1: counterbalancing activities in cell separation ofSaccharomyces cerevisiae. J. Gen. Microbiol.138:97–102.
7.Chu, S., J. DeRisi, M. Eisen, J. Mulholland, D. Botstein, P. O. Brown, and I. Herskowitz.1998. The transcriptional program of sporulation in budding yeast. Science282:699–705.
8.Cid, V. J., A. Duran, F. del Rey, M. P. Snyder, C. Nombela, and M. Sanchez.
1995. Molecular basis of cell integrity and morphogenesis inSaccharomyces cerevisiae. Microbiol. Rev.59:345–386.
9.Denison, S. H., S. Negrete-Urtasun, J. M. Mingot, J. Tilburn, W. A. Mayer, A. Goel, E. A. Espeso, M. A. Penalva, and H. N. J. Arst.1998. Putative membrane components of signal transduction pathways for ambient pH regulation inAspergillusand meiosis inSaccharomycesare homologous. Mol. Microbiol.30:259–264.
10.de Nobel, H., C. Ruiz, H. Martin, W. Morris, S. Brul, M. Molina, and F. M. Klis.2000. Cell wall perturbation in yeast results in dual phosphorylation of the Slt2/Mpk1 MAP kinase and in an Slt2-mediated increase inFKS2-lacZ
expression, glucanase resistance, and thermotolerance. Microbiology146:
2121–2132.
11.Dodou, E., and R. Treisman.1997. TheSaccharomyces cerevisiaeMADS-box transcription factor Rlm1 is a target for the Mpk1 mitogen-activated protein kinase pathway. Mol. Cell. Biol.17:1848–1859.
12.Futai, E., T. Maeda, H. Sorimachi, K. Kitamoto, S. Ishiura, and K. Suzuki.
1999. The protease activity of a calpain-like cysteine protease in Saccha-romyces cerevisiaeis required for alkaline adaptation and sporulation. Mol. Gen. Genet.260:559–568.
13.Garcia, R., C. Bermejo, C. Grau, R. Perez, J. M. Rodriguez-Pena, J. Francois, C. Nombela, and J. Arroyo.2004. The global transcriptional response to transient cell wall damage inSaccharomyces cerevisiaeand its regulation by the cell integrity signaling pathway. J. Biol. Chem.279:15183–15195. 14.Garcia-Rodriguez, L. J., A. Duran, and C. Roncero.2000. Calcofluor
anti-fungal action depends on chitin and a functional high-osmolarity glycerol response (HOG) pathway: evidence for a physiological role of the Saccha-romyces cerevisiaeHOG pathway under noninducing conditions. J. Bacteriol.
182:2428–2437.
15.Garrett-Engele, P., B. Moilanen, and M. S. Cyert.1995. Calcineurin, the Ca2⫹/calmodulin-dependent protein phosphatase, is essential in yeast
mu-tants with cell integrity defects and in mumu-tants that lack a functional vacuolar H⫹-ATPase. Mol. Cell. Biol.15:4103–4114.
16.Green, R., G. Lesage, A. M. Sdicu, P. Menard, and H. Bussey.2003. A synthetic analysis of theSaccharomyces cerevisiaestress sensor Mid2p, and identification of a Mid2p-interacting protein, Zeo1p, that modulates the PKC1-MPK1 cell integrity pathway. Microbiology149:2487–2499. 17.Groot, P. W. J. D., C. Ruiz, C. R. Vazquez de Aldana, and F. M. Klis.2001.
A genomic approach for the identification and classification of genes
on September 8, 2020 by guest
http://ec.asm.org/
volved in cell wall formation and its regulation inSaccharomyces cerevisiae. Comp. Funct. Genom.2:124–142.
18.Hohmann, S.2002. Osmotic stress signaling and osmoadaptation in yeasts. Microbiol. Mol. Biol. Rev.66:300–372.
19.Ketela, T., R. Green, and H. Bussey.1999.Saccharomyces cerevisiaeMid2p is a potential cell wall stress sensor and upstream activator of thePKC1-MPK1
cell integrity pathway. J. Bacteriol.181:3330–3340.
20.Krisak, L., R. Strich, R. S. Winters, J. P. Hall, M. J. Mallory, D. Kreitzer, R. S. Tuan, and E. Winter.1994.SMK1, a developmentally regulated MAP kinase, is required for spore wall assembly inSaccharomyces cerevisiae. Genes Dev.8:2151–2161.
21.Lagorce, A., N. C. Hauser, D. Labourdette, C. Rodriguez, H. Martin-Yken, J. Arroyo, J. D. Hoheisel, and J. Francois.2003. Genome-wide analysis of the response to cell wall mutations in the yeastSaccharomyces cerevisiae. J. Biol. Chem.278:20345–20357.
22.Lamb, T. M., and A. P. Mitchell.2003. The transcription factor Rim101p governs ion tolerance and cell differentiation by direct repression of the regulatory genesNRG1andSMP1inSaccharomyces cerevisiae. Mol. Cell. Biol.23:677–686.
23.Lamb, T. M., W. Xu, A. Diamond, and A. P. Mitchell.2001. Alkaline re-sponse genes of Saccharomyces cerevisiae and their relationship to the
RIM101pathway. J. Biol. Chem.276:1850–1856.
24.Levin, D. E.2005. Cell wall integrity signaling inSaccharomyces cerevisiae. Microbiol. Mol. Biol. Rev.69:262–291.
25.Li, W., and A. P. Mitchell.1997. Proteolytic activation of Rim1p, a positive regulator of yeast sporulation and invasive growth. Genetics145:63–73. 26.Lotz, H., K. Sohn, H. Brunner, F. A. Muhlschlegel, and S. Rupp.2004.
RBR1, a novel pH-regulated cell wall gene ofCandida albicans, is repressed by RIM101 and activated by NRG1. Eukaryot. Cell3:776–784.
27.Martin, H., J. M. Rodriguez-Pachon, C. Ruiz, C. Nombela, and M. Molina.
2000. Regulatory mechanisms for modulation of signaling through the cell integrity Slt2-mediated pathway inSaccharomyces cerevisiae. J. Biol. Chem.
275:1511–1519.
28.Mitchell, A. P.1994. Control of meiotic gene expression inSaccharomyces cerevisiae. Microbiol. Rev.58:56–70.
29.Mouyna, I., T. Fontaine, M. Vai, M. Monod, W. A. Fonzi, M. Diaquin, L. Popolo, R. P. Hartland, and J. P. Latge.2000. Glycosylphosphatidylinositol-anchored glucanosyltransferases play an active role in the biosynthesis of the fungal cell wall. J. Biol. Chem.275:14882–14889.
30.Penalva, M. A., and H. N. J. Arst.2004. Recent advances in the character-ization of ambient pH regulation of gene expression in filamentous fungi and yeasts. Annu. Rev. Microbiol.58:425–451.
31.Penalva, M. A., and H. N. J. Arst.2002. Regulation of gene expression by ambient pH in filamentous fungi and yeasts. Microbiol. Mol. Biol. Rev.
66:426–446.
32.Philip, B., and D. E. Levin.2001. Wsc1 and Mid2 are cell surface sensors for cell wall integrity signaling that act through Rom2, a guanine nucleotide exchange factor for Rho1. Mol. Cell. Biol.21:271–280.
33.Philips, J., and I. Herskowitz.1997. Osmotic balance regulates cell fusion during mating inSaccharomyces cerevisiae. J. Cell Biol.138:961–974.
34.Rodriguez-Pena, J. M., V. J. Cid, J. Arroyo, and C. Nombela.2000. A novel family of cell wall-related proteins regulated differently during the yeast life cycle. Mol. Cell. Biol.20:3245–3255.
35.Rose, M. D., F. Winston, and P. Hieter.1990. Methods in yeast genetics: a laboratory course manual. Cold Spring Harbor Laboratory Press, New York, N.Y.
36.Sambrook, J., E. F. Fritsch, and T. Maniatis.1989. Molecular cloning: a laboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
37.Saporito-Irwin, S. M., C. E. Birse, P. S. Sypherd, and W. A. Fonzi.1995.
PHR1, a pH-regulated gene ofCandida albicans, is required for morpho-genesis. Mol. Cell. Biol.15:601–613.
38.Serrano, R., D. Bernal, E. Simon, and J. Arino.2004. Copper and iron are the limiting factors for growth of the yeastSaccharomyces cerevisiaein an alkaline environment. J. Biol. Chem.279:19698–19704.
39.Serrano, R., A. Ruiz, D. Bernal, J. R. Chambers, and J. Arino.2002. The transcriptional response to alkaline pH inSaccharomyces cerevisiae: evidence for calcium-mediated signalling. Mol. Microbiol.46:1319–1333.
40.Smits, G. J., J. C. Kapteyn, H. van den Ende, and F. M. Klis.1999. Cell wall dynamics in yeast. Curr. Opin. Microbiol.2:348–352.
41.Tong, A. H., G. Lesage, G. D. Bader, H. Ding, H. Xu, X. Xin, J. Young, G. F. Berriz, R. L. Brost, M. Chang, Y. Chen, X. Cheng, G. Chua, H. Friesen, D. S. Goldberg, J. Haynes, C. Humphries, G. He, S. Hussein, L. Ke, N. Krogan, Z. Li, J. N. Levinson, H. Lu, P. Menard, C. Munyana, A. B. Parsons, O. Ryan, R. Tonikian, T. Roberts, A. M. Sdicu, J. Shapiro, B. Sheikh, B. Suter, S. L. Wong, L. V. Zhang, H. Zhu, C. G. Burd, S. Munro, C. Sander, J. Rine, J. Greenblatt, M. Peter, A. Bretscher, G. Bell, F. P. Roth, G. W. Brown, B. Andrews, H. Bussey, and C. Boone.2004. Global mapping of the yeast genetic interaction network. Science303:808–813.
42.Treton, B., S. Blanchin-Roland, M. Lambert, A. Lepingle, and C. Gaillardin.
2000. Ambient pH signalling in ascomycetous yeasts involves homologues of theAspergillus nidulansgenespalFandpaIH. Mol. Gen. Genet.263:505–513. 43.Trilla, J. A., A. Duran, and C. Roncero.1999. Chs7p, a new protein involved in the control of protein export from the endoplasmic reticulum that is specifically engaged in the regulation of chitin synthesis inSaccharomyces cerevisiae. J. Cell Biol.145:1153–1163.
44.Verna, J., R. A. Lodde, K. Lee, A. Vagts, and R. Ballester.1997. A family of genes required for maintenance of cell wall integrity and for the stress response inSaccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA94:13804– 13809.
45.Vicente, M. F., A. Basilio, and P. F. Cabello.2003. Microbial natural prod-ucts as a source of antifungals. Clin. Microbiol. Infect.9:15–32.
46.Wang, H., X. Tang, J. Liu, S. Trautmann, D. Balasundaram, D. McCollum, and M. K. Balasubramanian.2002. The multiprotein exocyst complex is essential for cell separation inSchizosaccharomyces pombe. Mol. Biol. Cell
13:515–529.
47.Xu, W., and A. P. Mitchell.2003. Yeast PalA/AIP1/Alix homolog Rim20p associates with a PEST-like region and is required for its proteolytic cleav-age. J. Bacteriol.183:6917–6923.