Copyright2001 by the Genetics Society of America
Multiple Functions of the Nonconserved N-Terminal Domain
of Yeast TATA-Binding Protein
Mark Lee
1and Kevin Struhl
Department of Biological Chemistry and Molecular Pharmacology, Harvard Medical School, Boston, Massachusetts 02115 Manuscript received December 15, 2000
Accepted for publication January 26, 2001
ABSTRACT
The binding protein (TBP) is composed of a highly conserved core domain sufficient for TATA-element binding and preinitiation complex formation as well as a highly divergent N-terminal region that is dispensable for yeast cell viability.In vitro, removal of the N-terminal region domain enhances TBP-TATA association and TBP dimerization. Here, we examine the effects of truncation of the N-terminal region in the context of yeast TBP mutants with specific defects in DNA binding and in interactions with various proteins. For a subset of mutations that disrupt DNA binding and the response to transcriptional activators, removal of the N-terminal domain rescues their transcriptional defects. By contrast, deletion of the N-terminal region is lethal in combination with mutations on a limited surface of TBP. Although this surface is important for interactions with TFIIA and Brf1, TBP interactions with these two factors do not appear to be responsible for this dependence on the N-terminal region. Our results suggest that the N-terminal region of TBP has at least two distinct functionsin vivo. It inhibits the interaction of TBP with TATA elements, and it acts positively in combination with a specific region of the TBP core domain that presumably interacts with another protein(s).
T
HE TATA-binding protein (TBP) plays a central relative to full-length TBP (Horikoshiet al.1990; Lieb-ermanet al.1991), primarily by increasing the rate of role in eukaryotic transcription, being required forTBP-TATA association (Kuddus and Schmidt 1993). accurate transcriptional initiation by all three nuclear
As a consequence, DNA binding by TBP mutants defec-RNA polymerases (Hernandez 1993; Struhl 1994;
tive for the TBP-TATA interaction can be restored by Burley andRoeder 1996). TBPs from a wide variety
removal of the N-terminal domain (Lee et al. 1992). of eukaryotes and archea have a highly conserved
180-Second, the yeast TBP core domain readily forms tran-residue C-terminal core domain (80% identical between
scriptionally inert dimers and higher order oligomers yeast and human) that is sufficient for TATA-element
in vitro(Katoet al.1994;Colemanet al.1995). Although binding and preinitiation complex formation. TBPs also
the physiological relevance of TBP dimerization is con-contain an N-terminal region that is very divergent
troversial (Jackson-Fisher et al. 1999; Geisberg and across species with respect to both length and sequence
Struhl 2000), the N-terminal domain inhibits oligo-and whose function is poorly understood. Human TBP
merization of full-length TBP, potentially enhancing the does not support yeast cell growth, but species-specific
probability that TBP is monomeric under physiologic difference maps to the C-terminal core domain, not the
conditions (Campbellet al.2000). Third, a peptide or N-terminal region (Cormacket al.1991;GillandTjian
a monoclonal antibody corresponding to a portion of 1991). Indeed, a single amino acid change within the
the N-terminal region selectively blocks TATA-depen-core domain enables human TBP to substitute for all
dent transcription by RNA polymerases II and III while of the essential activities of yeast TBPin vivo(Cormack
sparing RNA polymerase I and TATA-less promoter et al.1994).
transcription (Lescureet al. 1994). Fourth, yeast and The TBP core domain and full-length TBP behave
human NC2 are most effective in repressing transcrip-similarly in many biochemical assays, but there are
sev-tion in reacsev-tions reconstituted with TBP from the same eral differences that provide insight into the function of
species, and maximal repression requires the N-terminal the N-terminal domain. First, deletion of the N-terminal
region (Goppelt and Meisterernst 1996). Fifth, re-region results in enhanced binding and bending of DNA
moval of the N-terminal region of yeast TBP increases interactions with human TBP-associated factors (TAFs) and the formation of a hybrid TFIID complex (Zhou Corresponding author:Kevin Struhl, Department of Biological Chem- andBerk1995). Sixth, the N-terminal region of human istry and Molecular Pharmacology, Harvard Medical School, Boston,
TBP mediates cooperative binding with the small nu-MA 02115. E-mail: [email protected]
clear RNA activating protein (SNAP) complex to the
1Present address:Department of Medicine, Rm. S-101, Stanford
Uni-versity Medical Center, Stanford, CA 94305. U6 promoter and enhances U6 transcription (Mittal
and Hernandez 1997). Taken together, these results suggest a regulatory role for the N-terminal domain in governing the interactions of TBP with the TATA element, with other transcription factors, and with TBP itself. However, the physiological relevance of these bio-chemical properties of the TBP N-terminal region re-mains to be established.
In yeast cells, expression of the TBP core domain at physiological levels has minimal phenotypic conse-quence, indicating that the 63-residue N-terminal re-gion of TBP is dispensable for viability and the response to transcriptional activators (Cormacket al. 1991;Reddy andHahn1991). However, overexpression of the TBP
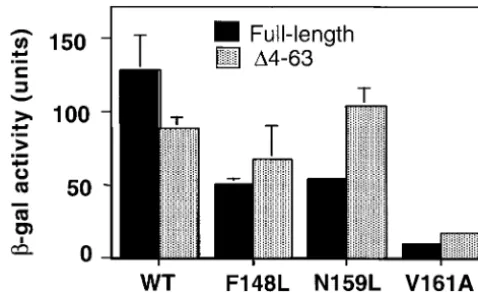
Figure1.—N-terminal truncation can rescue the transcrip-core domain inhibits cell growth, particularly on
nonfer-tional phenotypes of a subset of activation-deficient DNA-bind-mentable carbon sources (GillandTjian1991;Zhou ing mutants of yeast TBP.-Galactosidase activity of a Gal4-et al. 1991), and this toxicity is suppressed by concurrent dependentLacZreporter in wild-type and mutant TBP strains grown with galactose as the sole carbon source (inducing overexpression of Std1(Msn3) (Ganster et al. 1993).
conditions) is shown. Standard errors are given as error bars. Std1 directly associates with TBP, and deletion of the
Solid bars, full-length TBP derivatives; stippled bars, N-termi-N-terminal region qualitatively alters this interaction
nally truncated derivatives (⌬4-63). (Tillmanet al. 1995). Apart from these studies,
how-ever, relatively little is known about the function of the
TBP N-terminal regionin vivo. RESULTS
To examine the physiological role of the N-terminal
Deletion of the N-terminal region rescues the activa-region of yeast TBP, we deleted this activa-region in the
con-tion-deficient phenotype of a subset of DNA-binding text of a large number of yeast TBP mutants. Our results
mutants:Certain TBP mutants deficient in the response suggest that the N-terminal region of TBP has at least
to transcriptional activators contain mutations on the two distinct functionsin vivo. It inhibits the interaction
DNA-binding surface that dramatically reduce TATA-of TBP with TATA elements, and it functions in concert
element bindingin vitro(Kimet al. 1994;Arndtet al. with a specific surface of TBP that presumably interacts
1995; Leeand Struhl 1995). As removal of the N-ter-with another transcriptional regulatory protein.
minal region can restore TBP-TATA associationin vitro for some TBP mutants with DNA-binding defects (Lee et al. 1992), we examined the genetic properties of N-ter-MATERIALS AND METHODS
minally deleted versions of three TBP mutants deficient DNA molecules:TBP mutants used in this study have been for DNA binding and transcriptional activation (Lee described previously (CormackandStruhl 1993;Leeand
andStruhl1995). All three N-terminally deleted
deriv-Struhl1995, 1997). The N-terminally deleted derivatives in
atives support yeast cell growth at levels comparable to Table 1 derive from pBC83, aTRP1-marked centromeric
plas-mid with the 2.4-kb EcoRI-BamHI fragment containing the their full-length counterparts (Table 1). As previously yeastTBPgene with a deletion of residues 4–63 (Cormacket reported (Leeand Struhl1995), the F148L, N159L, al. 1991). The N-terminally deleted derivatives in Figure 2
and V161A derivatives are compromised for Gal4-depen-derive from pML3095, a derivative of pBC83 in which a FLAG
dent activation when compared to wild-type TBP (39, epitope was engineered at the N terminus of the TBP core
42, and 8% of wild-type activity, respectively; Figure 1). domain by subcloning of annealed oligonucleotides. Serial
deletions of the N-terminal region were generated by PCR Removal of the N-terminal region from wild-type TBP using the wild-typeTBPgene as template. results in a 40% decrease in Gal4-dependent activation,
Phenotypic analyses:TBP derivatives were assayed for their
in accord with previous results (Cormack et al. 1991; ability to support cell growth by spotting 104and 105cells on
ReddyandHahn1991). By contrast, N-terminal dele-glucose medium supplemented with casamino acids and
5-flu-oroorotic acid, using the plasmid shuffle assay and strain BY⌬2 tion confers increased transcriptional activation by the (Cormacket al. 1991); growth was monitored after 2–3 days N159L, V161A, and perhaps the F148L derivatives. incubation at 30⬚. S1 nuclease protection assays for measuring Deletion of the N-terminal region of TBP does not RNA levels and-galactosidase assays for determining
Gal4-affectHIS3TATA-element utilization:The results above dependent activation were performed as described elsewhere
and previous biochemical experiments indicate that the (Cormack and Struhl 1992; Lee and Struhl1995; Iyer
andStruhl1996). Intracellular levels of N-terminally deleted N-terminal region can act as an inhibitor of TBP func-TBPs were determined by Western blotting using monoclonal tion. For this reason, we examined whether the N-termi-antibody mAb58C9 (Santa Cruz Biotechnologies) against the nal region of TBP affects TATA-element utilization at FLAG epitope. To avoid the confounding effects of
homeo-theHIS3promoter in a manner similar to that of other static mechanisms on TBP levels, mutant TBP levels were
mea-general inhibitors of TBP function such as Mot1 (Col-sured in a strain containing hemagglutinin-tagged TBP (Lee
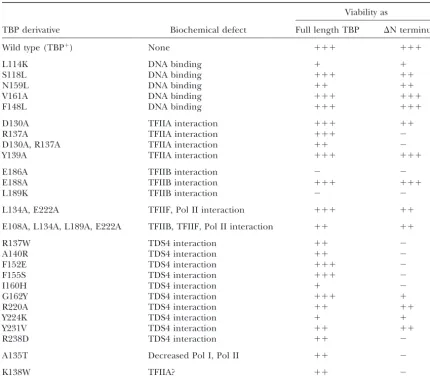
TABLE 1
Viability of defined TBP mutants with and without the N-terminal domain
Viability as
TBP derivative Biochemical defect Full length TBP ⌬N terminus
Wild type (TBP⫹) None ⫹⫹⫹ ⫹⫹⫹
L114K DNA binding ⫹ ⫹
S118L DNA binding ⫹⫹⫹ ⫹⫹
N159L DNA binding ⫹⫹ ⫹⫹
V161A DNA binding ⫹⫹⫹ ⫹⫹⫹
F148L DNA binding ⫹⫹⫹ ⫹⫹⫹
D130A TFIIA interaction ⫹⫹⫹ ⫹⫹
R137A TFIIA interaction ⫹⫹⫹ ⫺
D130A, R137A TFIIA interaction ⫹⫹ ⫺
Y139A TFIIA interaction ⫹⫹⫹ ⫹⫹⫹
E186A TFIIB interaction ⫺ ⫺
E188A TFIIB interaction ⫹⫹⫹ ⫹⫹⫹
L189K TFIIB interaction ⫺ ⫺
L134A, E222A TFIIF, Pol II interaction ⫹⫹⫹ ⫹⫹
E108A, L134A, L189A, E222A TFIIB, TFIIF, Pol II interaction ⫹⫹ ⫹⫹
R137W TDS4 interaction ⫹⫹ ⫺
A140R TDS4 interaction ⫹⫹ ⫺
F152E TDS4 interaction ⫹⫹⫹ ⫺
F155S TDS4 interaction ⫹⫹⫹ ⫺
I160H TDS4 interaction ⫹ ⫺
G162Y TDS4 interaction ⫹⫹⫹ ⫹
R220A TDS4 interaction ⫹⫹ ⫹⫹
Y224K TDS4 interaction ⫹ ⫹
Y231V TDS4 interaction ⫹⫹ ⫹⫹
R238D TDS4 interaction ⫹⫹ ⫺
A135T Decreased Pol I, Pol II ⫹⫹ ⫺
K138W TFIIA? ⫹⫹ ⫺
(Moqtaderiet al. 1996). TheHIS3promoter contains tein-protein interactions with other components of the transcription apparatus (Lescureet al. 1994;Zhouand a noncanonical TATA-like element (TC) that is
responsi-ble for initiation from the⫹1 site and a consensus TATA Berk1995; Goppelt and Meisterernst 1996). Dele-tion of the N-terminal region has minimal effects on element (TR) that is responsible for initiation from the
⫹13 site (IyerandStruhl1995). Loss of Mot1, NC2, or growth and on transcription by all three nuclear RNA polymerases. Thus, if the N-terminal region mediates TAF130 results in a dramatic decrease in TC-dependent
transcription from the⫹1 site, but does not affect TR- physiologically important functions, such functions
must be also performed by another region of TBP or dependent transcription from the⫹13 site. In contrast,
deletion of the N-terminal region of TBP does not affect by another protein(s).
To search for such potential redundant functions, we the pattern of transcriptional initiation, and hence
TATA-element utilization, at theHIS3promoter region constructed N-terminally deleted versions of a panel of TBP mutants with specific defects for interactions with (Figure 2). In addition, the N-terminally deleted version
of TBP has minimal effects on transcription by RNA TFIIA (Stargell andStruhl1995; Leeand Struhl 1997), TFIIB (Kimet al. 1994;LeeandStruhl1997), polymerases I and III. Thus, by the criterion of
TATA-element utilization at theHIS3promoter, the N-termi- TFIIF, RNA polymerase II (LeeandStruhl1997), and the RNA polymerase III factor Brf1 (Cormack and nal region of TBP behaves differently from other
pro-teins that can function as inhibitors of TBP function. Struhl1993). Deletion of the N-terminal region from TBP mutants deficient for interaction with DNA, TFIIB, Mutations within the core domain of TBP render the
N-terminal domain essential for viability:In addition to TFIIF, and RNA polymerase II produces only modest changes in growth (Table 1). In contrast, N-terminal its negative effect on DNA binding, the N-terminal
TFIIA-TABLE 2
Viability of N-terminally truncated TBP derivatives with and without an N-terminal FLAG epitope
Viability as
TBP Full-length ⌬N FLAG-⌬N
derivative TBP terminus terminus
Wild type (TBP⫹) ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹
A135T ⫹⫹ ⫺ ⫹
K138W ⫹⫹ ⫺ ⫹⫹
R238D ⫹⫹ ⫺ ⫹
R137A ⫹⫹⫹ ⫺ ⫺
A140R ⫹⫹ ⫺ ⫺
F152E ⫹⫹⫹ ⫺ ⫺
F155S ⫹⫹ ⫺ ⫺
I160H ⫹ ⫺ ⫺
level achieved by N-terminally deleted wild-type TBP (Figure 3).
N-terminal-dependent mutations map to a limited sur-face of TBP and define a new TBP function:The muta-tions that confer dependence on the N-terminal domain lie in close proximity within the three-dimensional struc-ture of the TBP core domain (Figure 4). Specifically, the five positions define a limited surface on the convex face of TBP that lines the inner portion of the groove formed by helix H2, helix H2⬘, and the segments preced-ing strand S1 and connectpreced-ing helix H2 and strand S1⬘. Amino acid substitutions at neighboring positions
Figure 2.—Deletion of the N terminus of TBP does not
significantly alter Pol I, II, or III transcription in vivo. S1 where the side chains project away from this putative nuclease protection assays for determination ofHIS3, DED1, interaction surface do not yield this phenotype. For rRNA, and tRNAw transcription levels in yeast strains
sup-example, N159, I160, and V161 are adjacent residues; ported by either full-length or N-terminally deleted (⌬4-63)
but, unlike I160, the side chains of N159 and V161 TBP are shown. All bands in the right lane are⬍50% higher
project, away from the convex surface of TBP, and muta-in muta-intensity than muta-in the left lane and likely represent a
differ-ence in total RNA used for each assay. tions at these positions do not confer this phenotype. Despite the overlap of this surface with the regions previously implicated in interactions with TFIIA and and Brf1-interaction surfaces results in lethality. Al- Brf1, disruption of interaction with neither of these two though removal of the N-terminal domain does not factors appears to be responsible for this N-terminal destabilize wild-type TBP (Cormacket al. 1991;Reddy dependence. First, only a subset of the mutations that and Hahn1991;Ganster et al. 1993), it might cause disrupt either TFIIA or Brf1 interaction yield this pheno-protein instability of TBP derivatives with mutations
within the core domain. To address this possibility, FLAG-tagged versions of these inviable N-terminally de-leted TBP mutants were generated to facilitate detection of their protein levels in vivo. Unexpectedly, three of these mutants (A135T, K138W, and R238D) are viable in the context of this N-terminal FLAG epitope tag (Ta-ble 2). The ability of an unrelated peptide (i.e.,FLAG) to functionally substitute for the N terminus in these three mutants suggests that these mutants, in the ab-sence of the N terminus, are unstable or misfolded. The
Figure3.—Western blot analysis of inviable N-terminally remainder of the inviable N-terminally truncated TBP
truncated TBP mutants. Whole-cell extracts from yeast strains mutants, however, are inviable in the context of the supported by wild-type TBP and coexpressing the indicated FLAG epitope, and their in vivo protein levels (with TBP derivatives were analyzed with monoclonal antibody
Figure5.—Sequential truncation of the N terminus of TBP. Viability of TBP derivatives with successive deletions of the N-terminal domain was assessed by growth on medium con-taining 5-fluoroorotic acid (FOA) in a plasmid shuffle assay. R137A and A140R respectively affect the TFIIA- and Brf1-interaction surfaces.
the absence of a particular interaction(s) normally me-diated by the core domain of TBP, at least one function
Figure4.—TBP mutants with N-terminal dependence map
of the N-terminal region becomes essential for yeast cell to a discrete surface of TBP. Two views of an X-ray
crystallo-graphic structure of yeast TBP (based on coordinates from growth. To localize the region within the N terminus NCBI Entrez MMDB file 1TBP; Chasman et al. 1993) are that performs this function, we analyzed a series of suc-shown. Structure renditions were generated by Cn3D version cessive deletions within the N-terminal region in the 3.0 (NCBI). Residues given in green represent positions tested
context of two TBP mutants that require the N-terminal in this study in which full-length and N-terminally truncated
domain: R137A (TFIIA-interaction mutant) and A140R derivatives exhibited identical phenotypes. Residues given in
red indicate positions at which mutations conferred N-termi- (Brf1-interaction mutant). Remarkably, all but the final nal dependence. Top, view of TBP molecular saddle perpen- 10 amino acids of the N-terminal domain could be re-dicular to the intramolecular dyad axis. Bottom, view of the moved without affecting the viability of these two TBP convex surface of TBP molecular saddle along the
intramolec-mutants (Figure 5). This 10-amino-acid segment likely ular dyad axis, looking into the groove formed by helix H2,
encodes a specific function, as an unrelated sequence helix H2⬘, and the segments preceding strand S1 and
connect-ing helix H2 and strand S1⬘. (FLAG) could not be functionally substituted (Table 2).
DISCUSSION type (Table 1). Second, Y139A, which is 20–30 times
the N-terminal region from these DNA-binding mutants region of TBP performs a positive function. In one model, the N-terminal domain stabilizes the interaction of TBP can partially restore their activation competency.
This result provides evidence that the inhibitory effect between TBP and the factor that interacts with the sur-face of the core domain identified by virtue of the syn-of the TBP N-terminal domain on DNA binding, long
recognizedin vitro(Horikoshiet al. 1990;Lieberman thetic lethal effects described above. The role of the N-terminal region of human TBP in mediating coopera-et al. 1991;Leeet al. 1992;KuddusandSchmidt1993),
has physiological relevance. tive binding with the SNAP complex at the U6 promoter (MittalandHernandez1997) is a precedent for such More generally, our results indicate that the
N-termi-nal region of TBP has an autoinhibitory role in vivo. a model, although the SNAP complex and its target site (PSE) do not exist in yeast cells. In a second model, the In otherwise wild-type cells, loss of this autoinhibitory
function does not significantly affect cell growth or gen- apparent positive role of the N-terminal region might actually be a negative function that is redundant with eral transcriptional functions of TBP. In part, the
mini-mal phenotype caused by loss of the TBP N-terminal a distinct negative regulatory function that is mediated by the limited surface of the TBP core domain. In this region might be due to functional redundancy with
other negative regulators of TBP function such as NC2 view, synthetic lethality results from the simultaneous elimination of multiple negative regulators of TBP func-(Gadbois et al. 1997; Kim et al. 1997, 2000; Prelich
1997;Lemaireet al. 2000;Xieet al. 2000), Mot1 (Auble tion. In accord with this model, the region immediately adjacent to yeast TBP core domain is required both for et al. 1994, 1997;Collart1996;MadisonandWinston
1997), the N terminus of TAF145 (Kokuboet al. 1998), the synthetic lethal interactions observed here (Figure 5) and for the toxicity observed upon overexpression and the NOT complex (Collart and Struhl 1994;
Collart 1996). Nevertheless, we suspect that loss of of the core domain (Zhouet al. 1991). By either model, our results and previous observations (ZhouandBerk this autoregulatory function might result in the
genera-tion of a hyperfuncgenera-tional TBP, thereby explaining why 1995; Goppelt andMeisterernst 1996;Mittal and Hernandez1997) suggest that the evolutionarily diver-overexpression of the TBP core domain strongly inhibits
cell growth even in the presence of wild-type TBP (Gill gent N-terminal region coevolved with other transcrip-tional regulators to mediate certain species-specific andTjian1991;Zhouet al. 1991).
Functional redundancy between the N terminus and functions of TBP.
a surface of the TBP core domain: We examined the We thank Martin Schmidt for providing STD constructs, Laurie effect of N-terminal deletion in the context of TBP muta- Stargell for helpful discussions at the initial stages of this work, and Lisete Fernandes and Ada Garcia for assistance in the preparation of tions representing 25 unique positions with the core
this manuscript. We also thank Susanna Chou, Sukalyan Chatterjee, domain. Mutations at only 5 of those positions yield a
and Irene Wu for their discussions and critical reading of the manu-lethal phenotype when combined with the N-terminal
script. This work was supported by a predoctoral fellowship to M.L. deletion (excluding the mutants that are rescued by the from the Howard Hughes Medical Institute and by research grants presence of an N-terminal FLAG epitope). Strikingly, to K.S. from the National Institutes of Health (GM 30186 and GM
53720). these 5 positions define a limited surface of TBP that
lines the inner portion of the groove formed by helix H2, helix H2⬘, and the segments preceding strand S1
LITERATURE CITED and connecting helix H2 and strand S1⬘. This synthetic
lethality between TBP mutations lacking the N-terminal Arndt, K. M., S. Ricupero-HovasseandF. Winston, 1995 TBP mutants defective in activated transcriptionin vivo. EMBO J.14:
region and those affecting a specific surface in the core
1490–1497.
domain provides genetic evidence for functional redun- Auble, D. T., K. E. Hansen, C. G. F. Mueller, W. S. Lane, J. Thorner et al., 1994 Mot1, a global repressor of RNA polymerase II tran-dancy between these two regions of TBP.
scription, inhibits TBP binding to DNA by an ATP-dependent These observations provide evidence both for a
posi-mechanism. Genes Dev.8:1920–1934.
tive function for the N-terminal region of TBP and for a Auble, D. T., D. Wang, K. W. PostandS. Hahn, 1997 Molecular analysis of the SNF2/SWI2 protein family member MOT1, an previously unidentified surface of the TBP core domain
ATP-driven enzyme that dissociates TATA-binding protein from that interacts with a transcriptional regulatory
pro-DNA. Mol. Cell. Biol.17:4842–4851.
tein(s). The factor(s) that interacts with this surface Burley, S. K., andR. G. Roeder, 1996 Biochemistry and structural biology of transcription factor IID (TFIID). Annu. Rev. Biochem. of the TBP core domain is unknown, although TFIIA,
65:769–799. TFIIB, and Brf1 do not appear to be strong candidates.
Campbell, K. M., R. T. Ranallo, L. A. StargellandK. J. Lumb, A role for the TBP N terminus in interactions with other 2000 Reevaluation of transcriptional regulation by TATA-bind-ing protein oligomerization: predominance of monomers. Bio-transcription factors is consistent with observations in
chemistry39:2633–2638. vitro in which the presence of the N-terminal domain
Chasman, D. I., K. M. Flaherty, P. A. SharpandR. D. Kornberg, affects the association of TBP with NC2 (Goppeltand 1993 Crystal structure of yeast TATA-binding protein and model for interaction with DNA. Proc. Natl. Acad. Sci. USA90:
Meisterernst 1996), TAFs (Zhou and Berk 1995),
8174–8178. Std1 (Tillman et al. 1995), and the SNAP complex
Chatterjee, S., and K. Struhl, 1995 Connecting a promoter-(MittalandHernandez1997). bound protein to the TATA-binding protein overrides the need
Coleman, R. A., A. K. Taggart, L. R. BenjaminandB. F. Pugh, Kokubo, T., M. J. Swanson, J. I. Nishikawa, A. G. Hinnebuschand
Y. Nakatani, 1998 The yeast TAF145 inhibitory domain and 1995 Dimerization of the TATA binding protein. J. Biol. Chem.
270:13842–13849. TFIIA competitively bind to TATA-binding protein. Mol. Cell. Biol.18:1003–1012.
Collart, M. A., 1996 TheNOT,SPT3, andMOT1genes functionally
interact to regulate transcription at core promoters. Mol. Cell. Kuddus, R., andM. C. Schmidt, 1993 Effect of the non-conserved N-terminus on the DNA binding activity of the yeast TATA bind-Biol.16:6668–6676.
Collart, M. A., andK. Struhl, 1994 NOT1(CDC39),NOT2(CDC36), ing protein. Nucleic Acids Res.21:1789–1796.
Kuras, L., andK. Struhl, 1999 Binding of TBP to promotersin NOT3, andNOT4encode a global negative regulator of
transcrip-tion that differentially affects TATA-element utilizatranscrip-tion. Genes vivois stimulated by activators and requires Pol II holoenzyme. Nature389:609–612.
Dev.8:525–537.
Cormack, B. P., andK. Struhl, 1992 The TATA-binding protein Lee, D. K., J. Dejong, S. Hashimoto, M. HorikoshiandR. G. Roeder, 1992 TFIIA induces conformational changes in TFIID via inter-is required for transcription by all three nuclear RNA polymerases
in yeast cells. Cell69:685–696. actions with the basic repeat. Mol. Cell. Biol.12:5189–5196.
Lee, M., andK. Struhl, 1995 Mutations on the DNA-binding surface
Cormack, B. P., andK. Struhl, 1993 Regional codon
randomiza-tion: defining a TATA-binding protein surface required for RNA of TBP can specifically impair the response to acidic activators in vivo. Mol. Cell. Biol.15:5461–5469.
polymerase III transcription. Science262:244–248.
Cormack, B. P., M. Strubin, A. S. PonticelliandK. Struhl, 1991 Lee, M., andK. Struhl, 1997 A severely defective TATA-binding protein-TFIIB interaction does not preclude transcriptional acti-Functional differences between yeast and human TFIID are
local-ized to the highly conserved region. Cell65:341–348. vationin vivo. Mol. Cell. Biol.17:1336–1345.
Lemaire, M., J. Xie, M. Meisterernstand M. A. Collart, 2000
Cormack, B. P., M. Strubin, L. A. StargellandK. Struhl, 1994
Conserved and nonconserved functions of yeast and human The NC2 repressor is dispensable in yeast mutated for the Sin4p component of the holoenzyme and plays roles similar to Mot1p TATA-binding proteins. Genes Dev.8:1335–1343.
Gadbois, E. L., D. M. Chao, J. C. Reese, M. R. Greenand R. A. in vivo. Mol. Microbiol.36:163–173.
Lescure, A., Y. Lutz, D. Eberhard, X. Jacq, A. Krol et al., 1994
Young, 1997 Functional antagonism between RNA polymerase
II holoenzyme and global negative regulator NC2in vivo. Proc. The N-terminal domain of the human TATA-binding protein plays a role in transcription from TATA-containing RNA polymer-Natl. Acad. Sci. USA94:3145–3150.
Ganster, R. W., W. ShenandM. C. Schmidt, 1993 Isolation of ase II and III promoters. EMBO J.13:1166–1175.
Li, X.-L., A.Virbasius, X.Zhuand M. R.Green, 1999 Enhancement STD1, a high-copy-number suppressor of a dominant negative
mutation in the yeast TATA-binding protein. Mol. Cell. Biol.13: of TBP binding by activators and general transcription factors. Nature389:605–609.
3650–3659.
Geisberg, J. V., andK. Struhl, 2000 TATA-binding protein mutants Lieberman, P. M., M. C. Schmidt, C. C. KaoandA. J. Berk, 1991 Two distinct domains in the yeast transcription factor IID and that increase transcription from enhancerless and repressed
pro-motersin vivo. Mol. Cell. Biol.20:1478–1488. evidence for a TATA box-induced conformational change. Mol. Cell. Biol.11:63–74.
Gill, G., andR. Tjian, 1991 A highly conserved domain of TFIID
displays species specificity in vivo. Cell65:333–340. Madison, J. M., andF. Winston, 1997 Evidence that Spt3 function-ally interacts with Mot1, TFIIA, and TATA-binding protein to
Goppelt, A., andM. Meisterernst, 1996 Characterization of the
basal inhibitor of class II transcription NC2 fromSaccharomyces confer promoter-specific transcriptional control in Saccharo-myces cerevisiae. Mol. Cell. Biol.17:287–295.
cerevisiae. Nucleic Acids Res.24:4450–4455.
Hernandez, N., 1993 TBP, a universal eukaryotic transcription fac- Mittal, V., andN. Hernandez, 1997 Role for the amino-terminal region of human TBP in U6 snRNA transcription. Science275:
tor? Genes Dev.7:1291–1308.
Horikoshi, M., T. Yamamoto, Y. Ohkuma, P. A. WeilandR. G. 1136–1140.
Moqtaderi, Z., Y. Bai, D. Poon, P.A. WeilandK. Struhl, 1996
Roeder, 1990 Analysis of structure-function relationships of
yeast TATA box binding factor TFIID. Cell61:1171–1178. TBP-associated factors are not generally required for transcrip-tional activation in yeast. Nature382:188–191.
Iyer, V., andK. Struhl, 1995 Mechanism of differential utilization
of thehis3TRand TCTATA elements. Mol. Cell. Biol15:7059– Prelich, G., 1997 Saccharomyces cerevisiae BUR6encodes a DRAP1/
NC2-alpha homolog that has both positive and negative roles in 7066.
Iyer, V., andK. Struhl, 1996 Absolute mRNA levels and transcrip- transcriptionin vivo. Mol. Cell. Biol.17:2057–2065.
Reddy, P., andS. Hahn, 1991 Dominant negative mutations in yeast tional initiation rates inSaccharomyces cerevisiae. Proc. Natl. Acad.
Sci. USA93:5208–5212. TFIID define a bipartite DNA-binding region. Cell65:349–357.
Stargell, L. A., andK. Struhl, 1995 The TBP-TFIIA interaction
Jackson-Fisher, A. J., C. Chitikila, M. MitraandB. F. Pugh, 1999
A role for TBP dimerization in preventing unregulated gene in the response to acidic activatorsin vivo. Science269:75–78.
Struhl, K., 1994 Duality of TBP, the universal transcription factor. expression. Mol. Cell3:717–727.
Kato, K., Y. Makino, T. Kishimoto, J. Yamauchi, S. Katoet al., Science263:1103–1104.
Tillman, T. S., R. W. Ganster, R. Jiang, M. Carlson andM. C.
1994 Multimerization of the mouse TATA-binding protein
(TBP) driven by its C-terminal conserved domain. Nucleic Acids Schmidt, 1995 STD1 (MSN3) interacts directly with the TATA-binding protein and modulates transcription of the SUC2 gene Res.22:1179–1185.
Kim, S., J. G.Na, M.Hampseyand D.Reinberg, 1997 The Dr1/ of Saccharomyces cerevisiae. Nucleic Acids Res.23:3174–3180.
Xiao, H., J. D. FriesenandJ. T. Lis, 1995 Recruiting TATA-binding DRAP1 heterodimer is a global repressor of transcription in vivo.
Proc. Natl. Acad. Sci. USA94:820–825. protein to a promoter: transcriptional activation without an up-stream activator. Mol. Cell. Biol.15:5757–5761.
Kim, S., K. Cabane, M. HampseyandD. Reinberg, 2000 Genetic
analysis of the Ydr1-Bur6 repressor complex reveals an intricate Xie, J., M. Collart, M. Lemaire, G. StelzerandM. Meisterernst, balance among transcriptional regulatory proteins in yeast. Mol. 2000 A single point mutation in TFIIA suppresses NC2 require-Cell. Biol.20:2455–2465. mentin vivo. EMBO J.19:672–682.
Kim, T. K., S. Hashimoto, R. J. I. Kelleher, P. M. Flanagan, R. D. Zhou, Q., andA. J. Berk, 1995 The yeast TATA-binding protein
Kornberget al., 1994 Effects of activation-defective TBP muta- (TBP) core domain assembles with human TBP-associated factors tions on transcription initiation in yeast. Nature369:252–255. into a functional TFIID complex. Mol. Cell. Biol.15:534–539.
Klages, N., andM. Strubin, 1995 Stimulation of RNA polymerase Zhou, Q., M. C. Schmidt andA. J. Berk, 1991 Requirement for II transcription initiation by recruitment of TBP in vivo. Nature acidic amino acid residues immediately N-terminal to the
con-374:822–823. served domain of Saccharomyces cerevisiae TFIID. EMBO J. 10:
Klein, C., andK. Struhl, 1994 Increased recruitment of TATA- 1843–1852. binding protein to the promoter by transcriptional activation