COMPLEMENTATION ANALYSIS O F THE TRYPTOPHAN PATHWAY IN ASPERGILLUS N I D U L A N S
C. F. ROBERTS
Department of Biology, Yale Uniuersity, New Hauen, Connecticut 0651 I
Received August 30, 1966
OLLOWING the extensive analysis of the gene-enzyme relationships of the Ftryptophan pathway in bacteria (YANOFSKY and LENNOX 1959; BLUME and BALBINDER 1966) in Saccharomyces and in Neurospora (BONNER, DE Moss and MILLS 1965), it has become O € interest to examine the pathway in the homo- thallic ascomycete Aspergillus nidulans (Eidam) Wint. This paper describes the complementation behaviour of tryptophan mutants, and the following paper by HUTTER and DE Moss (1967) the biochemical lesions and some of the proper- ties of the tryptophan enzymes. At this stage the system in Aspergillus appears very similar, perhaps identical, to that in Neurospora.
MATERIALS A N D METHODS
Isolation of mutants: Two strains of A. nidulans (Table 1) were obtained from crosses between translocation-free stocks supplied by DR. ETTA KAFER. Conidia were harvested from independ- ently isolated sub-clones, exposed to ultraviolet (UV) irradiation until about 90% were killed, and the proportion of mutants among the survivors was enriched by the filtration-concentration technic (WOODWARD, DE ZEEUW and SRB 1954). The filtration medium included an ammonium salt to eliminate mutants deficient in nitrate utilization. The final suspensions were plated in suitably supplemented minimal medium plus L-tryptophan and the tryptophan mutants detected by replica plating (ROBERTS 1959).
The mutants described in this paper were recovered from preliminary filtration enrichment experiments in which yields were irregular and often rather low. These, and subsequent, filtra- tion experiments have indicated three important factors in obtaining the mutants: the period over which the suspensions are filtered, the density at which the survivors are plated and the concentration of tryptophan in the plating medium.
The viable count was reduced from 104 to 105 per ml initially to 100 to 300 per ml at 24 hours. However, at 48 hours the viable count was reduced to about 10 per ml when a high pro- portion (about 10%) of mutants are recovered. The plating density should not be greater than 30 colonies per plate and the tryptophan concentration not less than 200 pg/ml. The presence of L-phenylalanine (50 to 200 ,ug/ml) and L-tyrosine (250 to 1,250 pg/ml) did not affect recovery of the tryptophan mutants. (The experiments were part of a screening program for multiply deficient aromatic mutants).
M e d i a Minimal medium (MM) has been described previously (FOLEY, GILE~ and ROBERTS 1965). Strains were maintained on a malt-extract agar (malt extract 20g, glucose 20g, Bacto- peptone 1 g, L-tryptophan 0.lg and agar 20g per litre) which promotes heavy conidiation.
Incubation was at 37°C.
This work was supported by Public Health Service training grant GM 397 and National Science Foundation grants
Present address: Department of Genetics, T h e University, Leicester, U.K.
G-11228 and GB 2176.
234 C . F. ROBERTS
Accumulation studies: Mutants were grown in 150 x 25 mm tubes containing 15 ml liquid M M supplemented with 15 pg/ml L-tryptophan. Maximum accumulations were observed at 5 days in gently shaken cultures or at 10 days in static cultures. Samples of the culture fluid were tested for indole by use of the indole reagent (YANOFSKY 1955) and for indole-glycerol by the ferric chloride reagent (YANOFSKY 1 x 6 ) . Fluorescent compounds, presumed to be mainly anthranilic acid were detected under a UV scanning lamp either directly in the culture fluid or after acidification and extraction into ethyl acetate. Accumulation was also evident on solid media by fluorescence under UV.
Complementation tests: A further improvement of the technic described previously ( FOLEY,
GILFS and ROBERTS 1965) produced very efficient synthesis of forced heterokaryons. A dense conidial suspension of a mutant in one of the two original strains was prepared in dilute liquid malt-extract medium and one small drop (<.05 ml) placed on each of a series of plates of MM
+
L-tryptophan (250 pg/ml). A loop of conidia from one of a series of mutants in the other original strain was mixed with each drop and the conidia were dug into the surface of the agar. The plates were incubated f o r 1 to 2 days and pieces of the resulting mycelium trans- ferred to the same medium before conidiation had started. Heterokaryons were observed at a high frequency as vigorous outgrowths from these transfers after 5 to 7 days further incubation. They were tested for tryptophan independence by transferring adjacent pieces of mycelium toTABLE 1
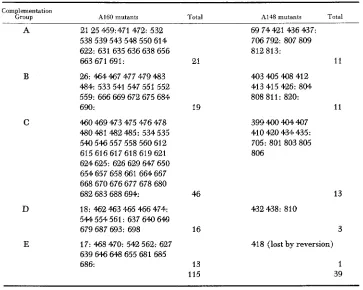
Functional analysis of the tryptophan mutants
~~~ ~~
Complementation
Group A160 mutants Total A148 mutants Total
A 21 25 459:471 472: 532
538 539 543 548 550 614 622: 631 635 636 638 656
663 671 691: 21
D
E
26: 464 467 477 479 483 484: 533541547551552 559: 666669672675684
690: 19
460 469 473 475 476 478 480 481 482 485: 534 535 540546557558560612 615616617618619621 624 625: 626 629 647 650 654 657 658 661 664 667 668670676677678680
682 683 688 694: 46
18: 462 463 465 466 474: 544 554 561: 637 640 649
679 687 693: 698 16
17: 468 470: 542 562: 627 639 646 648 655 681 685
686: 13
115
69 74 421 436 437: 706 792: 807 809 812 813:
11 403 4Q5 408 412
413 415 426: 804 808 811: 820:
11 399 400 404 4Q7 410 420 434 435: 705: 801 803 805 806
13 432 438: 810
3
418 (lost by reversion)
1 39
The colon (:) separates mutants isolated in different mutation experiments. Strain A160 is of genotype b i l . Acri w 3 . nicX. Strain A148 is of genotype pabal y .
The mutant alleles are re&esented: ’ bil =biotin; A c r l z dominant resistance to acriflavine; w3 =white conidia; nicX = nicotinic acid; pabal = p-aminobenzoic acid; y = yellow conidia; rib02 = riboflavine; facB = acetate re-
T R Y P T O P H A N PATHWAY IN ASPERGILLUS 235 MM and to MM
+
L-tryptophan. Where necessary, diploids were isolated from the hetero- karyons by the usual procedure (ROPER 1952).RESULTS A N D DISCUSSION
Characterization and complementation grouping of the tryptophan mutants: The mutants were tested for growth on anthranilic acid (25 pg/ml), indole
(IO pg/ml) or tryptophan (100 pg/ml) and separated into three classes: those responding to all three supplements, those responding to indole or tryptophan and those responding only to tryptophan. Mutants within each of these classes were then tested for complementation in heterokaryons, and most of them (137)
found to fall into four complementation groups A, B, C and D (Tables 1, 2). Representative mutants from each group were assigned to linkage groups by mitotic analysis ( MCCULLY and FORBES 1965).
The remaining 1 7 mutants all responded to anthranilic acid or indole but were complementary to mutants in group A. Of these mutants 14 were found to fall into a fifth complementation group (E) which is in a different chromosome from group A (Table 2). The remaining three mutants (420,535,664) complemented both A and E mutants. However, they were assigned to the same linkage group as group C mutants by mitotic haploidisation and were subsequently found non- complementary to trypC testers (see below).
T h e tryptophan loci: The main features of the tryptophan loci are outlined below. The enzyme defects mentioned were determined by R. HUTTER and J. A. DE Moss (see the following paper).
t r y p A (Linkage group 11). Mutants respond to anthranilic acid or indole; no accumulation was detected. These mutants lack anthranilic acid synthetase and are unable to synthesize anthranilic acid from chorismic acid. No interallelic complementation was observed in 50 heterokaryon tests.
trypB (Linkage group I). Mutants fail to respond to anthranilic acid or indole and require higher coilcentrations of L-tryptophan (100 pg/ml compared to 25 pg/ml for t r y p A ) for good growth on MM. Strong accumulation of indole- glycerol (io a lesser extent an tlxanilic acid) was observed in the culture medium.
NO indole was detected either in the medium o r in the cells. The indicated defect
TABLE 2
Characterization of the tryptophan loci
Locus
trrpD
Group E*
No. of mutants Growth response
32 Anth+ Ind+
30 Anth- Ind-
3 Anth+ Ind+
56 Anth- Ind+
19 Anth- Ind+
14 Anth+ Ind+
Accumulation Linkage group
None I1
Anth IG I
None VI11
+/-Anth
Anth I1
None VI
Anth = anthranilic acid, Iud = indole, IG = indole-glycerol.
23 6 C. F. ROBERTS
was found in tryptophan synthetase. No indole-utilizing mutants at this locus were recovered, paralleling their low frequency in Neurospora. One case of com- plementation between alleles was observed in 33 heterokaryon tests.
trypC (Linkage group VIII)
.
This complex locus maps 7 units from rib02 and 12 units from facB. Mutants are heterogenous; a few (420, 535, 664) respond to anthranilic acid and are thus apparently only defective in anthranilic acid synthetase. The bulk, however, do not respond to anthranilic acid and are varied with respect to accumulation of this intermediate. One of the anthranilate uti- lising mutants (535) was closely linked (approximately le4 recombination) to trypC-803.Complementation analysis reveals a class of noncomplementing mutants (A160485 and 30 others; A148-801 and six others). These mutants do not complement the anthranilate utilising mutants (420, 535, 664), and one (485) was found to lack three enzyme activities: anthranilate synthetase, phosphorib- osyl anthranilate (PRA) isomerase and indole-%glycerol phosphate (InGP) synthetase. However other mutants (473 and 612) apparently lacking all three enzymes are able to complement indicating very low levels of activity and perhaps instability of the enzyme (s) produced.
Complementation between trypC mutants is generally vigorous in both hetero- karyons and diploids, however, a few very weak positive reactions were observed in heterokaryons and in these cases the diploid complementation was far stronger. All mutants at the locus complement two trypA mutants tested, yielding normal vigorous heterokaryotic growth in the absence of tryptophan.
It is evident that the try$ locus is analagous to the tryp-1 locus in Neurospora (DE Moss and WEGMAN 1965). It deserves thorough analysis and also may prove of some interest as a system for testing the efficiency of heterokaryon and diploid complementation (compare FOLEY, GILES and ROBERTS 1965; APIRION 1966).
trypD (Linkage group 11; 15 units from trypA)
.
Mutants at this locus respond to indole but not anthranilic acid. They all accumulated much fluorescent mate- rial, most probably anthanilic acid, indicating deficieny in phosphoribosyl anthranilate transferase which proved to be the case. No instance of allelic complementation was found in 19 heterokaryons tested.Group E (Linkage group VI). The mutants respond to anthranilic acid, indi- cating a deficiency before this intermediate. When it 'was found that the mutants had anthranilic acid synthetase activity (and all the other activities in the tryp-
tophan path), it became evident that they may be blocked before chorismic acid. It was found that the tryptophan requirement is replaced by quinic acid (500 pg/ml) in the presence of phenylalanine ( 100 pg/ml) and tyrosine (500 pg/ml), and that quinic and shikimic acid have a sparing effect a t very low tryptophan levels 1 pg/ml). The mutants did not respond to high levels (200 M/ml) of nicotinic acid or nicotinamide, nor to phenylalanine and tyrosine alone.
T R Y P T O P H A N PATHWAY I N ASPERGILLUS 23 7 The isolation of group E mutants only in the A160 strain (biotin and nicotinic acid requiring) was suspicious, and further genetic analysis reveals that the E
phenotype is only expressed in the presence of the nic8 marker (nicotinic acid, linkage group VII)
.
It is thus possible that the group E defect is post-rather than pre-tryptophan. However further work is required to determine if the effect of the nic8 marker is physiological or chromosomal, and to explore the relationship between the group E mutants and the nidO marker which is in the same chro- mosome.Gene enzyme relationships in Ascomycetes: The present results are of interest since the close similarity of the tryptophan systems in Aspergillus and Neuro- spora are consistent with a close phylogenetic relationship between the two organisms, and their divergence lrom Saccharomyces ( BONNER, DE MOSS and MILLS 1965). The apparently identical pattern in the genetic control by trypA of anthranilate synthetase (tryp-2 in Neurospora) and by trypC of anthranilate synthetase, PRA isomerase and InGP synthetase (tryp-2 in Neurospora), and the formation of an enzyme aggregate with all three activities is most striking
(HUTTER and DE Moss 1967). The two loci are unlinked in both organisms. It is now clear that the structure of three of the enzymes in the histidine pathway are determined by a cluster of three genes in Neurospora (AHMED, CASE and GILES 1964), in Aspergillus (BERLYN 1966) and in Saccharomyces
(FINK 1965, 1966). Similarly five enzymes concerned in aromatic biosynthesis are determined by a cluster of five genes in Neurospora (GILES, CASE and PAR-
TRIDGE 1965). Preliminary results with Aspergillus indicate similarity of the aro- matic biosynthetic system with that in Neurospora as all multiply requiring mutants fall into only two complementation groups.
Attempts to separate the three histidine enzymes determined by the hist-3 rtgion in Neurospora (AHMED 1966) and the hi-4 region in Saccharomyces
238 C. F. ROBERTS
aggregate does not necessarily contradict this idea since two of the three enzymatic functions (PRA isomerase and IGP synthetase) are associated with a single polypeptide chain in E . coli (CREIGHTON and YANOFSKY 1966), but it remains to be determined if the activities are separable or not in the moulds.
I wish to thank PROFESSOR N. H. GILES for this support and interest in this work. It has been a pleasure to collaborate with DR. R. HUTTER and PROFESSOR J. A. DE Moss. I t is also a pleasure to acknowledge the excellent help of MR. SAMIR TAKLAN who did much of the experimental work with great care and diligence.
S U M M A R Y
Analysis of tryptophan requiring mutants in Aspergillus nidutans has revealed four unlinked loci controlling the ability to synthesize tryptophan from chorismic acid. One of the loci (try$) is complex, determining the production of a multi- functional enzyme aggregate analogous to the tryp-1 locus in Neurospora. It is noted that the close similarities in a number of gene-enzyme systems in Ascomy- cetes is possibly correlated with the production of multifunctional enzyme ag- gregates.
LITERATURE CITED
AHMED, A., 1966 A multi-enzyme complex specified by the histidine-3 region in Neurospora. (Abstr.) Genetics 54: 316.
AHMED, A., M. E. CASE, and N. H. GILES, 1964 The nature of complementation among mutants in the histidine-3 region of Neurospora crassa. Brookhaven Symp. Biol. 17: 53-65. APIRION, D., 1966 Recessive mutants a t unlinked loci which complement in diploids but not
in heterokaryons of Aspergillus nidulans. Genetics 53 : 935-941.
BERLYN, M., 1966 Gene-enzyme relationships in histidine biosynthesis in Aspergillus nidulans.
Ph.D. thesis, Yale University, New Haven, Connecticut.
BLUME, A. J., and E. BALBINDER, 1966 The tryptophan operon of Salmonella fyphimurium.
Fine structure analysis by deletion mapping and abortive transduction. Genetics 53 : 577-592.
BONNER, D. A., J. A. DE Moss, and S. E. MILLS, 1965 The evolution of an enzyme. pp. 305-318.
Evolving Genes and Proteins. Edited by V. BRYSON and H. J. VOGEL. Academic Press, New York.
Indole-3-glycerol phosphate synthetase of Escherichia coli, an enzyme of the tryptophan operon. J. Biol. Chem. 241 : 46164624.
Biochemical diversity in the tryptophan pathway. Biochem. Biophys. Res. C o m u n . 18: 850-857.
An enzyme aggregate in the tryptophan pathway of
Neurospora crassa. Proc. Natl. Acad. Sci. U.S. 54: 241-247.
Gene-enzyme relationships in histidine biosynthesis in yeast. Science 14.6:
525-527. - 1966 A cluster of genes controlling three enzymes in histidine biosynthesis in Saccharomyces cerevisiae. Genetics 53 : 4454-59.
Complementation at the adenylosuc- cinase locus in Aspergillus nidulans. Genetics 52: 1247-1263.
Evidence for an aromatic (arom) operon in Neurospora crassa. (Abstr.) Genetics 52: 444-446.
CREIGHMN, T. E., and C. YANOFSKY, 1966
DE Moss, J. A., 1965
DE Moss, J. A., and J. WEGMAN, 1965
FINK, G. R., 1964
FOLEY, JEAN M., N. H. GI-, and C. F. ROBERTS, 1965
T R Y P T O P H A N P A T H W A Y I N ASPERGILLUS 239
Symposium on structure and function of genes and chromosomes: A sum- mation. Nat. Cancer Inst. Monograph 18: 341-354.
Enzyme analysis of the tryptophan pathway in Asper- gillus nidulans. Genetics 5 5 : 241-247.
Genes and proteins involved i n histidine biosynthesis in Salmonella. Brookhaven Symp. Biol. 17 :
15-50.
MCCULLY, K. S., and E. FORBES, 1965 The use of p-fluorophenylalanine with ‘master strains’ of Aspergillus nidulans for assigning genes to linkage groups. Genet. Res. 6: 352-359. PIWARD, J., and B. J. WALLACE, 1966 Distribution and function of genes concerned with aro-
matic biosynthesis in Escherichia coli. J. Bacteriol. 91 : 1494-1508.
ROBERTS, C. F., 1959 A replica plating technique f o r the isolation of nutritionally exacting mutants of a filamentous fungus (AspergiZlus nidulans)
.
J. Gen. Microbiol. 20: 540-548. ROPER, J. A., 1952 Production of heterozygous diploids in filamentous fungi. Experentia 8:1 4 1 5 .
WOODWARD, V. W., J. R. DE ZEEUW, and A. M. SRB, 1954 The separation and isolation of partic- ular biochemical mutants of Neurospora by differential germination of conidia followed by filtration and selective plating. Proc. Natl. Acad. Sci. US. 40: 192-200.
Tryptophan synthetase from Neurospora. pp. 233-238. Methods in En- zymology, Vol. 2. Edited by S. P. COLOWICK and N. 0. KAPLAN. Academic Press, New York.
-
The enzymatic conversion of anthranilic acid to indole. J. Biol. Chem. 223:1 71 -1 84.
Transduction and recombination study of linkage rela- tionships among the genes controlling tryptophan synthesis in Escherichia coli. Virology
8: 425-447. GILES, N. H., 1965
HU~TTER, R., and J. A. DE MOSS, 1967
LOPER, J. C., M. GRABNER, R. C. STAHL, Z. HARTMAN, and P. E. IFARTMAN, 1964
YANOFSKY, C., 1955
1956