0095-1137/10/$12.00 doi:10.1128/JCM.02526-09
Copyright © 2010, American Society for Microbiology. All Rights Reserved.
Molecular Epidemiology of Vancomycin-Resistant
Enterococcus faecium
: a
Prospective, Multicenter Study in South American Hospitals
䌤
Diana Panesso,
1,2† Jinnethe Reyes,
1† Sandra Rinco
´n,
1Lorena Díaz,
1Jessica Galloway-Pen
˜a,
2,7Jeannete Zurita,
3Carlos Carrillo,
4‡ Altagracia Merentes,
5Manuel Guzma
´n,
5Javier A. Adachi,
6Barbara E. Murray,
2,7and Cesar A. Arias
1,2*
Molecular Genetics and Antimicrobial Resistance Unit, Universidad El Bosque, Bogota´, Colombia1; Center for the Study of Emerging and
Re-Emerging Pathogens, Division of Infectious Diseases, Department of Internal Medicine, University of Texas Medical School at Houston, Houston, Texas2; Hospital Vozandes, Quito, Ecuador3; Laboratorio Clínico Carlos Carrillo, Lima, Peru4;
Centro Me´dico Caracas, Caracas, Venezuela5; University of Texas, M. D. Anderson Cancer Center, Houston, Texas6; and
Department Microbiology and Molecular Genetics, University of Texas Medical School at Houston, Houston, Texas7
Received 29 December 2009/Returned for modification 8 February 2010/Accepted 25 February 2010
Enterococcus faeciumhas emerged as an important nosocomial pathogen worldwide, and this trend has been associated with the dissemination of a genetic lineage designated clonal cluster 17 (CC17). Entero-coccal isolates were collected prospectively (2006 to 2008) from 32 hospitals in Colombia, Ecuador, Peru´, and Venezuela and subjected to antimicrobial susceptibility testing. Genotyping was performed with all vancomycin-resistantE. faecium (VREfm) isolates by pulsed-field gel electrophoresis (PFGE) and mul-tilocus sequence typing. All VREfm isolates were evaluated for the presence of 16 putative virulence genes (14fmsgenes, theespgene of E. faecium[espEfm], and thehylgene ofE. faecium [hylEfm]) and plasmids carrying thefms20-fms21(pilA),hylEfm, andvanAgenes. Of 723 enterococcal isolates recovered,E. faecalis was the most common (78%). Vancomycin resistance was detected in 6% of the isolates (74% of which were
E. faecium). Eleven distinct PFGE types were found among the VREfm isolates, with most belonging to sequence types 412 and 18. The ebpAEfm-ebpBEfm-ebpCEfm (pilB) and fms11-fms19-fms16 clusters were detected in all VREfm isolates from the region, whereasespEfmandhylEfmwere detected in 69% and 23% of the isolates, respectively. Thefms20-fms21 (pilA) cluster, which encodes a putative pilus-like protein, was found on plasmids from almost all VREfm isolates and was sometimes found to coexist withhylEfmand thevanAgene cluster. The population genetics of VREfm in South America appear to resemble those of such strains in the United States in the early years of the CC17 epidemic. The overwhelming presence of plasmids encoding putative virulence factors and vanA genes suggests that E. faecium from the CC17 genogroup may disseminate in the region in the coming years.
Enterococci are now recognized as important nosocomial pathogens worldwide and in the United States are ranked as the second most common cause of nosocomial infections, after staphylococci (14). The two most common enterococ-cal species isolated from clinienterococ-cal samples areEnterococcus faecalisandE. faecium; however, the proportions of isolates of these two species have dramatically changed in the last decade. Whereas up until the early to mid-1990sE. faecalis
was the overwhelmingly predominant species isolated in U.S. hospitals (37), by 2008, the proportion of nosocomialE. faecalis/E. faecium strains was ca. 1.5:1, and there was an important increase in the incidence of E. faecium nosoco-mial infections (14). Moreover, more than 80% ofE. fae-cium isolates currently recovered from U.S. hospitals are resistant to vancomycin, and virtually all of them (⬎90%) exhibit ampicillin resistance (14). On the contrary, the prev-alence of vancomycin resistance in E. faecalisremains low
(⬍7% of isolates), and ampicillin resistance continues to be extremely rare. This change in the epidemiology of entero-coccal infections has been attributed to the increased ability of a genogroup ofE. faecium (designated clonal cluster 17 [CC17]) to colonize the gastrointestinal tract of humans, cause disease (37), and exhibit high levels of resistance to most antienterococcal antibiotics. Several virulence and col-onizing factors have been postulated to explain this in-creased virulence (4, 25, 28) and include the following: (i) the presence of an intactacm gene, which encodes a colla-gen adhesin and which has been associated with the patho-genesis of endocarditis in members of CC17 (25); (ii) theesp
gene ofE. faecium(espEfm), which codes for an enterococcal surface protein, which has been shown to play a role in biofilm formation (12), and which transiently aggravates experimental urinary tract infection (18); (iii) the fms (E. faeciumsurface protein-encoding) genes, which encode cell wall-anchored proteins, including subunits of the enterococ-cal pili (13, 31); and (iv) thehylgene ofE. faecium(hylEfm;
which encodes a putative glycosyl hydrolase), which is car-ried by transferable plasmids that have been shown to in-crease the ability of a laboratory strain of E. faecium to colonize the gastrointestinal tracts of mice and also enhance the virulence of a commensal strain ofE. faeciumin exper-imental peritonitis (4, 28).
* Corresponding author. Mailing address: Division of Infectious Diseases, University of Texas Medical School at Houston, 6431 Fannin St., MSB 2.112, Houston, TX 77030. Phone: (713) 500-6765. Fax: (713) 500-5495. E-mail: [email protected].
† Diana Panesso and Jinnethe Reyes contributed equally to this study.
‡ Deceased.
䌤Published ahead of print on 10 March 2010.
1562
on May 16, 2020 by guest
http://jcm.asm.org/
In South America (Brazil and Argentina), vancomycin-resis-tant (VR) enterococcal infections have been described since 1998 (9, 20). In a prospective multicenter surveillance study conducted in 2003, the prevalence of VR among enterococci in Colombia was found to be lower (9.7%) than that in the United States (1), and the isolation of similar percentages of VR enterococci has been described more recently (21). Al-though the emergence of E. faecium CC17 has been docu-mented in Brazil, Chile, and Paraguay (16, 19, 39), prospective studies have not been performed and limited data regarding the presence of the potential virulence determinants of CC17
E. faecium (including hylEfm-containing plasmids) in South America are available. Thus, we performed a multinational, multicenter prospective surveillance study with the aim of characterizing the population genetics of enterococci circulat-ing in the northern region of South America. Clinical isolates (excluding colonizing isolates) were collected prospectively from 32 hospitals in four countries (Colombia, Ecuador, Peru´, and Venezuela) and were further characterized at the molec-ular level.
(This study was presented in part at the 48th Annual Inter-science Conference on Antimicrobial Agents and Chemother-apy/Infectious Diseases Society of America 46th Annual Meet-ing, Washington, DC, 25 to 28 October 2008, abstr. C2-1998.)
MATERIALS AND METHODS
Specimen collection.A multicenter study was performed from February 2006 through February 2008 to evaluate the molecular epidemiology of enterococci in four South American countries. Each participating tertiary-care hospital (a total of 32 hospitals, from the Latin American Network of Antimicrobial Resistance) in Colombia (22 hospitals in six cities), Ecuador (5 hospitals in one city), Peru´ (3 hospitals in one city), and Venezuela (2 hospitals in one city) collected consec-utive enterococcal isolates (duplicate organisms from the same patient were excluded). The samples were collected by the clinical laboratory from each participating hospital and corresponded to samples from hospitalized patients in Colombia, Peru´, and Venezuela. In Ecuador, the clinical laboratory also col-lected samples from ambulatory services and outpatient clinics. The isolates originated from the following clinical specimens: blood, urine, secretions from surgical wounds, peritoneal fluid, abdominal abscesses, joint aspirates, osteomy-elitis aspirates, pleural fluid, pericardial effusion, cerebral abscesses, and cere-brospinal fluid (CSF). In an attempt to avoid isolates that likely represented colonization, enterococci recovered from sputum, rectal swabs, catheters, or skin (unless they originated in an infected surgical wound) were excluded. Each hospital identified the microorganisms by using either automated methods (per-formed with the Vitek or MicroScan system) or manual methods, and once the corresponding isolate was included in the study, it was sent to the reference laboratory (located in Bogota´, Colombia) via courier in transport medium (Amies; BBL). Upon arrival, the reference laboratory confirmed the purity of the isolate and confirmed the identification by molecular methods using multiplex PCR for enterococci (see below).
Susceptibility testing and molecular methods for species-specific identifica-tion andvangenotype determination.Susceptibility tests for enterococci were performed by an agar dilution method with ampicillin, ciprofloxacin, chloram-phenicol, linezolid, vancomycin, and teicoplanin. The isolates were also evalu-ated for high-level resistance (HLR) to streptomycin (2,000g/ml) and genta-micin (500g/ml). All isolates from blood and CSF identified asE. faecaliswere screened for the presence of the-lactamase enzyme by use of a nitrocefin test, and all susceptibility tests were conducted by the methodology suggested by the CLSI (8). MIC determinations were carried out with the inclusion of a control reference strain (E. faecalisATCC 29212). Species-specific identification of the enterococci and determination of thevangenotype of the vancomycin-resistant isolates were performed by PCR, as described previously (2, 10) (primers for the detection of the recently describedvanLgenotype [6] were not included).E. faeciumBM4147 (vanA),E. faecalisV583 (vanB), andE. gallinarumBM4174 (vanC1) were used as control strains for MIC determinations and PCR assays.
Molecular typing.Pulsed-field gel electrophoresis (PFGE) of the vancomycin-resistantE. faecium(VREfm) isolates was performed by use of some
modifica-tions of a previously described method (22), and the banding patterns were initially interpreted by visual inspection, according to the criteria specified by Tenover et al. (35). Subsequently, cluster analysis was performed by the method of Dice and the unweighted-pair group method using average linkages (UPGMA). The band tolerance was set at 1.5%, and the threshold cutoff value was set at 85%. Representative isolates of each PFGE subtype of VREfm were further genotyped by multilocus sequence typing (MLST) by a standard protocol described previously (15, 29). Fragments of seven housekeeping genes (atpA,ddl,
gdh,purK,gyd,pstS, andadk) were sequenced, the allelic profiles were obtained, and the sequence type (ST) for each unique allelic profile was designated on the basis of the information at the MLST website (http://efaecium.mlst.net).
Detection of plasmids and putative virulence genes.The preparation of colony lysates on nylon membranes and hybridization under high-stringency conditions were performed as described previously (33). DNA probes for thehylEfm,espEfm, and 14fmsgenes were obtained by using previously published primers (11, 27, 32); the probes were radiolabeled by use of the RadPrime DNA labeling system (Invitrogen, Carlsbad, CA). Plasmid detection in representative isolates of VREfm belonging to CC17 was performed by the use of S1 nuclease and PFGE, according to a previously described protocol (4, 5). This methodology allows the detection and estimation of the size of large bacterial plasmids in the presence of genomic DNA by PFGE (5). Plasmid bands were subsequently hybridized with probes targeting thehylEfm,fms20,fms21(pilA), andvanAgenes to determine if these genes were colocated in the same plasmid, as was reported previously (17).
E. faeciumTX0016 (strain DO) (23) and ERV-99 (4) were used as controls for colony hybridizations and the PFGE-S1 nuclease experiments.
RESULTS
Phenotypic characteristics of enterococcal isolates from the northern region of South America.A total of 760 consecutive
Enterococcussp. isolates was collected in the four countries. Thirty-seven isolates were not included in the study due to protocol violations (contamination, isolates were from the same patient, the source was not included in the protocol, or misidentification). Among the isolates included (a total of 723), Colombian hospitals contributed 309 (43%) of the iso-lates, Peruvian hospitals contributed 164 (23%), Ecuadorian hospitals contributed 148 (20%), and Venezuelan hospitals contributed 102 (14%). The majority (78%) wereE. faecalis
isolates mostly recovered from urine (54%) and blood (14%) (no-lactamase-producing isolate was detected).E. faecium
comprised 15% (n⫽111) of the enterococcal isolates (Table 1). The most common clinical sources ofE. faeciumincluded urine (38%), blood (21%), and surgical wound infections (13%). Other enterococcal species also identified by PCR in-cludedE. avium,E. hirae,E. casseliflavus,E. gallinarum, andE. durans. We were unable to determine the enterococcal species for 5% (n⫽35) of the isolates.
The resistance rates of the enterococci from the Andean region are shown in Table 1. AllE. faecalisisolates were sus-ceptible to ampicillin and linezolid, whereas the rates of HLR to gentamicin and streptomycin were 28% and 30%, respec-tively; ciprofloxacin resistance was found in 29% of the E. faecalisisolates (Table 1). In contrast, the rates of resistance to ampicillin and vancomycin and HLR to gentamicin and strep-tomycin inE. faeciumwere much higher (76%, 31%, 36%, and 44%, respectively). The highest rates of resistance to ampicillin and vancomycin and HLR to gentamicin and streptomycin in
E. faeciumisolates were found in Peru´ (90%, 48%, 55%, and 65%, respectively), and the lowest were found in Venezuela (50%, 25%, 8%, and 21%, respectively). Among the ampicil-lin-resistant E. faecium isolates from the Andean region, a total of 51 isolates (46%) had MICs of⬎64g/ml. Linezolid was active against all theE. faeciumisolates tested, and only
on May 16, 2020 by guest
http://jcm.asm.org/
4% of the E. faeciumisolates tested were resistant to chlor-amphenicol.
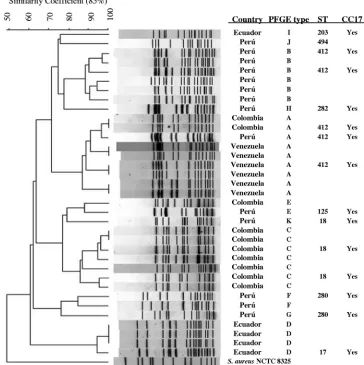
E. faeciumCC17 is also predominant in the northern region of South America.Thirty-five VREfm isolates were found, and all of them exhibited the VanA phenotype (high levels of resistance to both vancomycin and teicoplanin) and harbored thevanAgene cluster. Typing by PFGE revealed 11 different banding patterns (patterns A to K) (Fig. 1). A single PFGE banding pattern was predominant in each country (referred to as type A in Venezuela, type B in Peru´, type C in Colombia, and type D in Ecuador) (Fig. 1), although isolates from differ-ent countries (e.g., types A and E) could be found to have related PFGE types. MLST analysis of representative isolates of each PFGE type indicated that the most frequent ST was ST412; other STs included ST17, ST18, ST125, ST203, ST280, and ST282 (all of which were associated with the CC17 genetic lineage). A representative of PFGE pattern J from a Peruvian hospital was found to belong to a novel ST (ST494) which had not been reported previously.
Only three vancomycin-resistantE. faecaliswere isolated in the 2 years of the study; all three isolates were recovered in Colombia, harbored the vanB gene cluster, and were previ-ously characterized as belonging to the same unique clonal cluster that has been circulating in that country since 2001 (ST2 by trilocus sequence typing and MLST) (3, 7).
CC17 vancomycin-resistantE. faeciumisolates from South America are enriched in thefmsgenes.It had previously been shown that the members of CC17 of E. faecium harbor a number of genes encoding sortase-anchoring motif
(LPXTG)-containing cell wall proteins (13, 31); at least 15 of these genes encode predicted microbial surface components recognizing adhesive matrix molecule (MSCRAMMs) proteins in a U.S. strain (strain TX0016 [strain DO]) that was sequenced (32). It has been proposed that these genes function as important virulence determinants mediating adhesion to mammalian tis-sues, and several are enriched in the CC17 genogroup (11, 13, 31). Thus, we set out to investigate if these genes were also frequently found in CC17 isolates ofE. faecium from Latin America. Figure 2 shows the results of colony hybridizations of isolates with several putative virulence genes (including 14 of thefmsgenes;acm, which may be present as an intact gene or pseudogene, was not tested since it has been detected in almost all isolates studied previously [26]). Indeed, the four clusters of genes encoding MSCRAMM proteins (31) were highly repre-sented among the South AmericanE. faeciumisolates tested. TheebpAEfm-ebpBEfm-ebpCEfm(pilB) andfms11-fms19-fms16
clusters, which harbor genes encoding putative proteins with conserved pilin motifs and E-boxes of pilus proteins of Gram-positive bacteria, were present in all our South American VR isolates (Fig. 2). Interestingly, thescm gene, which has been shown to encode a collagen adhesin (the second collagen ad-hesin fromE. faecium), was present in more than 90% of the isolates. The otherfmsgenes were also frequently present at rates ranging from 67% to 97% in the VREfm isolates. The
[image:3.585.43.540.82.359.2]espEfmandhylEfmgenes, which have previously been associated with CC17 (28, 38), were detected in South American VRE. faeciumisolates (69% and 23% of isolates, respectively).
TABLE 1. Resistance profiles of enterococci from the northern region of South America
Organism (no. of isolates)
% (no.) of isolates resistant to the following antimicrobial agentsa:
VAN TEC CIP CHL LZD AMP HLR-GEN HLR-STR
Enterococci
Colombia (309) 5 (16) 3 (10) 30 (93) 12 (38) 0 (0) 10 (31) 19 (60) 29 (89)
Perú (164) 9 (15) 8 (14) 55 (91) 17 (28) 0 (0) 16 (27) 53 (87) 47 (77)
Ecuador (148) 5 (7) 3 (5) 25 (37) 17 (26) 0 (0) 10 (15) 28 (42) 28 (42)
Venezuela (102) 9 (9) 6 (6) 28 (29) 9 (9) 0 (0) 12 (12) 15 (15) 17 (17)
Total (723) 6 (47) 5 (35) 35 (250) 14 (101) 0 (0) 12 (85) 28 (204) 31 (225)
Enterococcus faecalis
Colombia (251) 1 (3) 0 (0) 26 (65) 13 (33) 0 (0) 0 (0) 17 (44) 27 (67)
Perú (128) 0 (0) 0 (0) 48 (61) 20 (26) 0 (0) 0 (0) 54 (69) 44 (56)
Ecuador (122) 0 (0) 0 (0) 22 (27) 20 (25) 0 (0) 0 (0) 26 (32) 27 (33)
Venezuela (59) 0 (0) 0 (0) 19 (11) 13 (8) 0 (0) 0 (0) 22 (13) 17 (11)
Total (560) 0.5 (3) 0 (0) 29 (164) 16 (92) 0 (0) 0 (0) 28 (158) 30 (167)
Enterococcus faecium
Colombia (41) 24 (10) 24 (10) 61 (25) 7 (3) 0 (0) 76 (31) 34 (14) 44 (18)
Perú (29) 48 (14) 48 (14) 93 (27) 3 (1) 0 (0) 90 (26) 55 (16) 65 (19)
Ecuador (17) 29 (5) 29 (5) 59 (10) 0 (0) 0 (0) 88 (15) 47 (8) 41 (7)
Venezuela (24) 25 (6) 25 (6) 58 (14) 4 (1) 0 (0) 50 (12) 8 (2) 21 (5)
Total (111) 31 (35) 31 (35) 68 (76) 4 (5) 0 (0) 76 (84) 36 (40) 44 (49)
Other enterococcal species
Colombia (17) 18 (3) 0 (0) 18 (3) 12 (2) 0 (0) 0 (0) 12 (2) 23 (4)
Perú (7) 14 (1) 0 (0) 43 (3) 14 (1) 0 (0) 14 (1) 28 (2) 28 (2)
Ecuador (9) 22 (2) 0 (0) 0 (0) 11 (1) 0 (0) 0 (0) 22 (2) 22 (2)
Venezuela (19) 16 (3) 0 (0) 21 (4) 0 (0) 0 (0) 0 (0) 0 (0) 5 (1)
Total (52) 17 (9) 0 (0) 19 (10) 8 (4) 0 (0) 2 (1) 11 (6) 17 (9)
aVAN, vancomycin; TEC, teicoplanin; CIP, ciprofloxacin; CHL, chloramphenicol; LZD, linezolid; AMP, ampicillin; HLR-GEN, high-level resistance to gentamicin; HLR-STR, high-level resistance to streptomycin.
on May 16, 2020 by guest
http://jcm.asm.org/
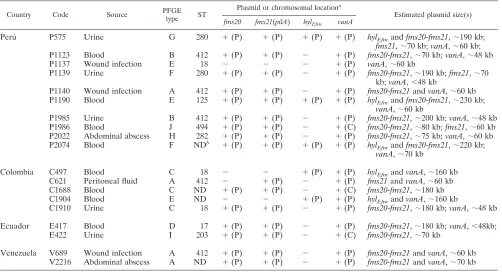
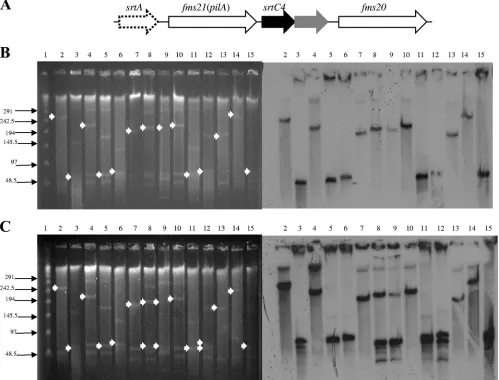
The fms20-fms21 (pilA) cluster and hylEfm are carried on plasmids associated with thevanA gene in South American VREfm. It was previously shown that hospital-associated E. faecium isolates from diverse geographical locations carry large, transferable plasmids containing thehylEfmgene (which
encodes a putative family 84 glycosyl hydrolase enzyme) that, additionally, are associated with vancomycin and aminoglyco-side resistance genes (4). These hylEfm-containing plasmids have emerged as important virulence and colonization deter-minants of CC17 ofE. faecium(4, 28). More recently, one of thefmsclusters (fms20-fms21[pilA]) encoding pilus-like pro-teins was also found to be located in a largehylEfm-carrying plasmid in U.S. strain TX0016 (strain DO) (17). Table 2 and Fig. 3 show the results of S1 nuclease digestion; PFGE; and the hybridization of representatives of South American VR E.
faeciumisolates with probes targeting thefms20-fms21(pilA) cluster, hylEfm, and vanA. When it was present, the fms20
-fms21(pilA) cluster was always encoded on plasmids ranging from ca. 60 kb to 242 kb. Of note, in some strains, fms21 -containing plasmids were found not to harbor the other mem-ber of the cluster (fms20), and in three strains (all from Peru´)
an additional copy offms21was detected in a different plasmid (strains P575, P1139, and P1986; Table 2 and Fig. 3). Further-more, the fms20-fms21cluster was found to be in the same plasmid ashylEfmin three strains (estimated plasmid sizes, 190 and 230 kb) and on the samevanA-containing plasmid in three strains (Table 2). No plasmid was found to carry thefms20
-fms21cluster,hylEfm, andvanAtogether.
DISCUSSION
In the work described here, we performed the first prospec-tive, multinational study evaluating the molecular epidemiol-ogy of VR enterococci in the northern region of South Amer-ica. Our results indicate that the prevalence of vancomycin resistance is relatively low in the participating countries (only 6% of isolates included in this study) compared to that in the United States. In our previous multicenter study performed with isolates from 15 hospitals in Colombia in 2001 and 2002, the rate of vancomycin resistance among enterococcal isolates was 9.7%. Also, unlike in the United States,E. faecalis contin-ues to be, by far, the most frequent enterococcal species
iso-FIG. 1. Molecular typing of VRE. faeciumfrom the Andean region of South America. The phylogenetic tree was constructed by use of the Dice coefficient and UPGMA clustering; the band tolerance was set at 1.5%, and the threshold cutoff value was set at 85%. ST, sequence type; CC17, clonal cluster 17.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:4.585.108.472.73.438.2]lated in this region (ratio of E. faecalis to E. faecium, 5:1). Vancomycin resistance inE. faecaliswas found sporadically in only three isolates from one city of the participating countries, and ampicillin was active against allE. faecalisisolates, under-scoring the fact that-lactams and glycopeptides continue to be useful for the treatment of enterococcal infections in the northern region of South America. On the other hand, the
[image:5.585.133.451.71.272.2]rates of HLR to aminoglycosides inE. faecaliswere ca. 30%, and there were important regional variations; for example, in the participating hospitals from Peru´, more than 50% of theE. faecalisisolates exhibited HLR to gentamicin and 44% exhib-ited HLR to streptomycin, indicating that the treatment of endovascular infections with the combination of a cell wall agent and an aminoglycoside in some areas of South America
FIG. 2. Frequency of detection of putative virulence genes in 35 VRE. faeciumisolates from the Andean region of South America. The numbers on the left-hand side indicate the four clusters of genes encoding characterized putative proteins of the enterococcal pili (30).
TABLE 2. Plasmid or chromosomal location offms20,fms21(pilA),hylEfm, andvanAin vancomycin-resistantEnterococcus faeciumisolates
from the Andean region of South America
Country Code Source PFGE
type ST
Plasmid or chromosomal locationa
Estimated plasmid size(s)
fms20 fms21(pilA) hylEfm vanA
Perú P575 Urine G 280 ⫹(P) ⫹(P) ⫹(P) ⫹(P) hylEfmandfms20-fms21,⬃190 kb;
fms21,⬃70 kb;vanA,⬃60 kb; P1123 Blood B 412 ⫹(P) ⫹(P) ⫺ ⫹(P) fms20-fms21,⬃70 kb;vanA,⬃48 kb
P1137 Wound infection E 18 ⫺ ⫺ ⫺ ⫹(P) vanA,⬃60 kb
P1139 Urine F 280 ⫹(P) ⫹(P) ⫺ ⫹(P) fms20-fms21,⬃190 kb;fms21,⬃70 kb;vanA,⬍48 kb
P1140 Wound infection A 412 ⫹(P) ⫹(P) ⫺ ⫹(P) fms20-fms21andvanA,⬃60 kb P1190 Blood E 125 ⫹(P) ⫹(P) ⫹(P) ⫹(P) hylEfmandfms20-fms21,⬃230 kb;
vanA,⬃60 kb
P1985 Urine B 412 ⫹(P) ⫹(P) ⫺ ⫹(P) fms20-fms21,⬃200 kb;vanA,⬃48 kb P1986 Blood J 494 ⫹(P) ⫹(P) ⫺ ⫹(C) fms20-fms21,⬃80 kb;fms21,⬃60 kb P2022 Abdominal abscess H 282 ⫹(P) ⫹(P) ⫺ ⫹(P) fms20-fms21,⬃75 kb;vanA,⬃60 kb
P2074 Blood F NDb ⫹(P) ⫹(P) ⫹(P) ⫹(P) hyl
Efmandfms20-fms21,⬃220 kb;
vanA,⬃70 kb
Colombia C497 Blood C 18 ⫺ ⫺ ⫹(P) ⫹(P) hylEfmandvanA,⬃160 kb
C621 Peritoneal fluid A 412 ⫺ ⫹(P) ⫺ ⫹(P) fms21andvanA,⬃60 kb
C1688 Blood C ND ⫹(P) ⫹(P) ⫺ ⫹(C) fms20-fms21,⬃180 kb
C1904 Blood E ND ⫺ ⫺ ⫹(P) ⫹(P) hylEfmandvanA,⬃160 kb
C1910 Urine C 18 ⫹(P) ⫹(P) ⫺ ⫹(P) fms20-fms21,⬃180 kb;vanA,⬃48 kb
Ecuador E417 Blood D 17 ⫹(P) ⫹(P) ⫺ ⫹(P) fms20-fms21,⬃180 kb;vanA,⬍48kb;
E422 Urine I 203 ⫹(P) ⫹(P) ⫺ ⫹(C) fms20-fms21,⬃70 kb
Venezuela V689 Wound infection A 412 ⫹(P) ⫹(P) ⫺ ⫹(P) fms20-fms21andvanA,⬃60 kb V2216 Abdominal abscess A ND ⫹(P) ⫹(P) ⫺ ⫹(P) fms20-fms21andvanA,⬃70 kb
a
P, plasmid; C, chromosomal. b
ND, not done; MLST was not performed because the isolate had a PFGE pattern closely related to that of another isolate whose MLST profile was determined.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:5.585.43.545.437.709.2]may be challenging and alternative bactericidal therapies should be sought. Also, we did not find important differences in phenotypic or genotypic characteristics between VREfm isolates from urine and blood (except that the blood isolates had higher rates of resistance to chloramphenicol [20%] than those VREfm isolates that originated in urine [0%]).
Although our findings indicate that the proportion of E. faecalis/E. faecium is considerably higher in South American hospitals than in U.S. hospitals (14), the South American VREfm isolates have genotypic and phenotypic characteristics similar to those of their U.S. counterparts: (i) South American
VREfm isolates mostly belong to ST412 and ST18 (the CC17 lineage), indicating that a hospital-associated genetic lineage is present in the region and supporting previous findings of stud-ies performed in other countrstud-ies of South America (16, 19, 39); (ii) the South American CC17 VREfm isolates also harbor an important number of MSCRAMM genes, including the four gene clusters encoding predicted components of theE. faecium
[image:6.585.43.542.72.452.2]pili (particularly theebpEfm and thefms11-fms19-fms16 clus-ters) (13, 31, 32), a finding that is consistent with the fact that the presence of these genes appears to be more reliably asso-ciated with members of the CC17 genetic lineage (11) than the
FIG. 3. Plasmids containing thefms20gene or thefms21gene (pilA), or both genes, in representativeE. faeciumisolates from the Andean region of South America. (A) Genetic organization offms20-fms21(pilA) inE. faeciumTX0016 (strain DO). This locus also contains a predicted class C sortase-encoding gene (srtC4) and a class A sortase-encoding gene (srtA) (30). (B) S1 nuclease digestion of total DNA ofE. faeciumstrains, followed by PFGE (left panel) and hybridization with anfms20probe (right panel). Lane 1, bacteriophage lambda ladder (molecular sizes [in kilobases] are shown to the left); lanes 2 and 14, TX0016 (strain DO); lanes 3, ERV-99 (4); lanes 4, P1985 (Peru´); lanes 5, P1123 (Peru´); lanes 6, P2022 (Peru´); lanes 7, E417 strain (Ecuador); lanes 8, P1139 (Peru´); lanes 9, P575 (Peru´); lanes 10, P2074 (Peru´); lanes 11, E422 (Ecuador); lanes 12, P1986 (Peru´); lanes 13, C1688 (Colombia); lanes 15, V2216 (Venezuela). Plasmid bands are shown as linearized fragments on the gel; the white arrows indicate the plasmid bands hybridizing with thefms20probe. (C) S1 nuclease digestion of total DNA ofE. faeciumstrains, followed by PFGE (left panel) and hybridization with anfms21(pilA) probe (right panel). Lane 1, bacteriophage lambda ladder (molecular sizes [in kilobases] are shown to the left); lanes 2 and 14, TX0016 (strain DO); lanes 3, ERV-99 (4); lanes 4, P1985 (Peru´); lanes 5, P1123 (Peru´); lanes 6, P2022 (Peru´); lanes 7, E417 (Ecuador); lanes 8, P1139 (Peru´); lanes 9, P575 (Peru´); lanes 10, P2074 (Peru´); lanes 11, E422 (Ecuador); lanes 12, P1986 (Peru´); lanes 13, C1688 (Colombia); lanes 15, V2216 (Venezuela). The white arrows indicate the plasmid bands hybridizing with thefms21
(pilA) probe.
on May 16, 2020 by guest
http://jcm.asm.org/
presence ofespEfmandhylEfm; and (iii) high-level of resistance to ampicillin is commonly found inE. faeciumisolates from the region, with the majority of isolates exhibiting MICs of⬎64
g/ml.
Recently, a genotypic analysis of early clinical isolates ofE. faeciumin the United States (11) showed that CC17E. faecium
isolates have been circulating in the United States since at least 1982, and it was postulated that an ancestral genogroup ofE. faeciumenriched with thefmsgenes was able to subsequently acquire additional virulence and antibiotic resistance determi-nants (such asespEfm,hylEfm, HLR to ampicillin, and thevan
gene clusters) and establish itself as an epidemic hospital-associated pathogen in the United States. Our results suggest that the population genetics of VREfm in the northern region of South America resembles that of isolates responsible for early outbreaks in the United States (in about 1982) (11), and it is tempting to speculate that an increase in the frequency of
E. faeciuminfections in South America may be expected in the following decade as members of the hospital-associated lin-eage disseminate and establish themselves as nosocomial pathogens in the region. Nonetheless, it is important to point out that the number of hospitals participating in this study differed in each country; while 22 hospitals from six different cities were represented in Colombia, a lower number of hos-pitals in the other three countries participated in the study. Thus, it is difficult to make countrywide generalizations of the molecular epidemiology of enterococcal isolates in Peru´, Ven-ezuela, and Ecuador. However, our data provide important information on the type of organisms currently circulating in the region.
An important goal of our study was to determine if South American VREfm isolates also contain genes encoding puta-tive pilus proteins that are present on plasmids coexisting with
hylEfm or vancomycin resistance gene clusters, as previously shown for other strains (4, 17). Enterococcal pili are important cell surface structures that have been implicated in the patho-genesis of experimental endocarditis and urinary tract infec-tions (24, 34). These trimeric units are composed of a major backbone subunit and two minor subunits (36) with an N-terminal signal sequence and a C-N-terminal cell wall-sorting signal (30) encoded by different genes. Indeed, we found that one of these clusters (fms20-fms21[pilA]) is carried by large plasmids in the majority of isolates studied. Moreover, in some strains, the fms20-fms21 (pilA) cluster coexists on the same plasmid with thehylEfmgene or with vanA, providing further evidence that the plasmid dissemination of virulence, resis-tance, and/or colonization determinants may be a strategy adopted by hospital-associated lineages ofE. faecium world-wide. This finding also supports the hypothesis that the An-dean region of South America may be in the early stages of an
E. faeciumepidemic and that as thefms20-fms21/hylEfm -carry-ing plasmids disseminate, the ability of the organisms to colo-nize the gastrointestinal tract of patients and cause disease may increase.
In summary, this is the first multicenter, multinational pro-spective study of the molecular epidemiology of VR entero-cocci in South America. Our results indicate that the preva-lence ofE. faeciumis low among clinical isolates, but genotypic analysis supports the fact that the population genetics ofE. faeciumis similar to that seen in the United States more than
25 years ago, suggesting that the dissemination of these highly adaptedE. faeciumorganisms in the hospital environment may occur in South America in the coming years through the clonal expansion of hospital-adapted isolates and with the horizontal transfer of virulence and colonization determinants.
ACKNOWLEDGMENTS
This report is dedicated to the memory of Carlos Carrillo. We thank the Universidad El Bosque for financial support and are indebted to Karen Jime´nez and German Contreras for technical as-sistance. We also thank Maria Virginia Villegas for facilitating the coordination of the participating centers in Colombia. We are grateful to Kavindra S. Singh and Jaime Moreno for providing technical ex-pertise and Jouko Sillanpaa for critical review of the manuscript.
The following personnel and hospitals participated in the collection of isolates: in Bogota´, Colombia, Claudia London˜o and Martha Her-rera (Fundacio´n Salud Bosque); Constanza Correa (Hospital Simo´n Bolívar); Norma Montoya (Clínica de Occidente); Wilson Daza and Martha Uzeta (Clínica del Nin˜o); Narda Olarte and Martha Garzo´n (Hospital El Tunal); Gloria Gallo (Hospital Santa Clara); Fernando Pen˜aloza and Nubia Escobar (Hospital Occidente de Kennedy); Mar-tha Ruiz (Clínica San Pedro Claver); Carlos A´ lvarez, Nidia Torres, and Ziomara Gonza´lez (Hospital San Ignacio); Clara Luz Rico (Fundacio´n Santa Fe de Bogota´); Giovanni Rodríguez and Deise Rojas (Clínica Infantil Colsubsidio); Juan Benavides, Maritza Pe´rez, and Esperanza Guevara (Clínica Saludcoop Jorge Pin˜eros Corpas); and Patricia Ar-royo (Instituto Nacional de Cancerología); in Cali, Colombia, María Virginia Villegas and Beatriz Vanegas (Centro Me´dico Imbanaco); María del Socorro Rojas (Clínica Saludcoop Occidente Cali); and Ernesto Martínez and Nancy Villamarín (Hospital Universitario del Valle); in Medellín, Colombia, Sergio Jaramillo and Jaime Lo´pez (Hospital Pablo Tobo´n Uribe) and Magda Ca´rdenas (Clínica Salud-coop Juan Luis London˜o); in Bucaramanga, Colombia, Adriana Pinto (Clínica La Foscal and Fundacio´n Cardiovascular); in Neiva, Colom-bia, Marino Cabrera and Luz Eneyda Quintero (Hospital Universita-rio Hernando Moncaleano Perdomo) and Carmen Elisa Llanos (Hos-pital Universitario San Jorge); in Ecuador, Hos(Hos-pital Vozandes, Hospital Eugenio Espejo, Hospital Baca Ortiz, Hospital Carlos Andre´s Andrade Marín, and Hospital General de las Fuerzas Armadas; in Peru´, Gene Martínez Medina and Susana Kuwae de Okuhama (Labo-ratorio Clínico Carlos Carrillo); Federico Yan˜ez Rojas and Liliana Alvarado (Hospital Nacional Sergio Bernales); Greenlandia Ferreyros Brandon and María Silva (Instituto Nacional de Enfermedades Neo-pla´sicas); and Rosa Avurio Usca and Gladys Patin˜o Soto (Hospital Nacional Hipo´lito Unanue); and in Venezuela, Adele Rizzi (Centro Me´dico de Caracas and Hospital Vargas de Caracas).
This work was funded in part by an independent research grant from Pfizer, SA. C.A.A. and B.E.M. are supported by a K99/R00 Pathway to Independence Award (award 4R00 AI72961) and R01 grant AI067861 from the National Institute of Allergy and Infectious Diseases, respec-tively. D.P. was partially funded by a graduate scholarship from the Instituto Colombiano para el Desarrollo de la Ciencia y Tecnología, Francisco Jose´ de Caldas, COLCIENCIAS; and S.R. was supported by an ASM-PAHO Infectious Disease Epidemiology and Surveillance Fellowship.
C.A.A. has received lecture fees from Pfizer, Novartis, and Merck and grant support from Pfizer. B.E.M. has had grant support from Johnson & Johnson, Astellas, Palumed, and Intercell and has served as consultant for Astellas Pharma US Inc., Theravance Inc., Cubist, Tar-ganta Therapeutics Corporation, Johnson & Johnson, Pfizer, Astra-Zeneca, and Wyeth-Ayerst. J.Z. reports the receipt of research grants from Pfizer and Wyeth. M.G. has served as consultant for Pfizer, Merck and Co., Wyeth, and Becton and Dickinson.
REFERENCES
1.Arias, C. A., J. Reyes, M. Zu´n˜iga, L. Cortes, C. Cruz, L. Rico, and D. Panesso.2003. Multicentre surveillance of antimicrobial resistance in en-terococci and staphylococci from Colombian hospitals, 2001–2002. J. Anti-microb. Chemother.51:59–68.
2.Arias, C. A., B. Robredo, K. V. Singh, C. Torres, D. Panesso, and B. E. Murray.2006. Rapid identification ofEnterococcus hiraeandEnterococcus
on May 16, 2020 by guest
http://jcm.asm.org/
duransby PCR and detection of a homologue of theE. hirae mur-2gene in
E. durans. J. Clin. Microbiol.44:1567–1570.
3.Arias, C. A., S. Rincon, S. A. Chowdhury, E. Martínez, W. Coronell, J. Reyes, S. R. Nallapareddy, and B. E. Murray.2008. MRSA USA300 clone and VREF—a U.S.-Colombian connection? N. Engl. J. Med.359:2177–2179. 4.Arias, C. A., D. Panesso, K. V. Singh, L. B. Rice, and B. E. Murray.2009.
Cotransfer of antibiotic resistance genes and ahylEfm-containing virulence
plasmid inEnterococcus faecium. Antimicrob. Agents Chemother.53:4240– 4246.
5.Barton, B. M., G. P. Harding, and A. J. Zuccarelli.1995. A general method for detecting and sizing large plasmids. Anal. Biochem.226:235–240. 6.Boyd, D. A., B. M. Willey, D. Fawcett, N. Gillani, and M. R. Mulvey.2008.
Molecular characterization ofEnterococcus faecalisN06-0364 with low-level vancomycin resistance harboring a novelD-Ala–D-Ser gene cluster,vanL. Antimicrob. Agents Chemother.52:2667–2672.
7.Chowdhury, S. A., C. A. Arias, S. R. Nallapareddy, J. Reyes, R. J. Willems, and B. E. Murray.2009. A trilocus sequence typing scheme for hospital epidemiology and subspecies differentiation of an important nosocomial pathogen,Enterococcus faecalis. J. Clin. Microbiol.47:2713–2719. 8.Clinical and Laboratory Standards Institute.2009 Performance standards
for antimicrobial susceptibility testing; 19th informational supplement M100-S19. Clinical and Laboratory Standards Institute, Wayne, PA. 9.Dalla Costa, L. M., D. C. Souza, L. T. Martins, R.C. Zanella, M. C.
Bran-dilone, S. Bokermann, H. S. Sader, and H. A. Souza.1998. Vancomycin-resistantEnterococcus faecium: first case in Brazil. Braz. J. Infect. Dis.2:160– 163.
10.Dutka-Malen, S., S. Evers, and P. Courvalin.1995. Detection of glycopep-tide resistance genotypes and identification to the species level of clinically relevant enterococci by PCR. J. Clin. Microbiol.33:24–27. (Erratum,33:
1434.)
11.Galloway-Pen˜a, J. R., S. R. Nallapareddy, C. A Arias, G. M. Eliopoulos, and B. E. Murray.2009. Analysis of clonality and antibiotic resistance among early clinical isolates ofEnterococcus faeciumin the United States. J. Infect. Dis.200:1566–1573.
12.Heikens, E., M. J. Bonten, and R. J. Willems.2007. Enterococcal surface protein Esp is important for biofilm formation of Enterococcus faecium
E1162. J. Bacteriol.189:8233–8240.
13.Hendrickx, A. P., M. J. Bonten, M. van Luit-Asbroek, C. M. Schapendonk, A. H. Kragten, and R. J. Willems.2008. Expression of two distinct types of pili by a hospital-acquiredEnterococcus faeciumisolate. Microbiology154:
3212–3223.
14.Hidron, A. I., J. R. Edwards, J. Patel T. C. Horan, D. M. Sievert, D. A. Pollock, and S. K. Fridkin.2008. Antimicrobial-resistant pathogens associ-ated with healthcare-associassoci-ated infections: annual summary of data reported to the national healthcare safety network at the Centers for Disease Control and Prevention, 2006–2007. Infect. Control. Hosp. Epidemiol.29:996–1011. (Erratum,30:107, 2009.)
15.Homan, W. L., D. Tribe, S. Poznanski, M. Li, G. Hogg, E. Spalburg, J. D. Van Embden, and R. J. Willems.2002. Multilocus sequence typing scheme forEnterococcus faecium. J. Clin. Microbiol.40:1963–1971.
16.Khan, M. A., J. B. Northwood, R. G. J. Loor, A. T. R Tholen, E. Riera, M. Falcon, A. van Westreenen, and J. P. Hays.2009. High prevalence of ST-78 infection-associated vancomycin-resistantEnterococcus faeciumfrom hospi-tals in Asuncio´n, Paraguay. Clin. Microbiol. Infect. doi:10.1111/j.1469-0691.2009.02898x.
17.Kim, D. S., K. V. Singh, S. R. Nallapareddy, X. Qin, D. Panesso, C. A. Arias, and B. E. Murray.2010. Thefms21(pilA)-fms20locus encoding one of four distinct pili ofEnterococcus faeciumis harbored on a large transferable plasmid associated with gut colonization and virulence. J. Med. Microbiol.
59:505–507.
18.Leendertse, M., E. Heikens, and L. M. Wijnands.2009. Enterococcal surface protein transiently aggravatesEnterococcus faecium-induced urinary tract infection in mice. J. Infect. Dis.200:1162–1165.
19.Lopez, M., J. C. Hormazabal, A. Maldonado, G. Saavedra, F. Baquero, J. Silva, C. Torres, and R. del Campo.2009. Clonal dissemination of Entero-coccus faecalis ST201 and Enterococcus faecium CC17-ST64 containing Tn5382-vanB2among 16 hospitals in Chile. Clin. Microbiol. Infect.15:586– 588.
20.Marín, M. E., J. R. Mera, R. C. Arduino, A. P. Correa, T. M. Coque, D.
Stamboulian, and B. E. Murray.1998. First report of vancomycin-resistant
Enterococcus faeciumisolated in Argentina. Clin. Infect. Dis.26:235–236. 21.Moet, G. J., R. N. Jones, D. J. Biedenbach, M. G. Stilwell, and T. R. Fritsche.
2007. Contemporary causes of skin and soft tissue infections in North Amer-ica, Latin AmerAmer-ica, and Europe: report from the SENTRY Antimicrobial Surveillance Program (1998–2004). Diagn. Microbiol. Infect. Dis.57:7–13. 22.Murray, B. E., K. V. Singh, J. D. Heath, B. R. Sharma, and G. M. Weinstock.
1990. Comparison of genomic DNAs of different enterococcal isolates using restriction endonucleases with infrequent recognition sites. J. Clin. Micro-biol.28:2059–2063.
23.Nallapareddy, S. R., G. M. Weinstock, and B. E. Murray.2003. Clinical isolates ofEnterococcus faeciumexhibit strain-specific collagen binding me-diated by Acm, a new member of the MSCRAMM family. Mol. Microbiol.
47:1733–1747.
24.Nallapareddy, S. R., K. V. Singh, J. Sillanpa¨a¨, D. A. Garsin, M. Ho¨o¨k, S. L. Erlandsen, and B. E. Murray.2006. Endocarditis and biofilm-associated pili ofEnterococcus faecalis. J. Clin. Invest.116:2799–2807.
25.Nallapareddy, S. R., K. V. Singh, and B. E. Murray.2008. Contribution of the collagen adhesin Acm to pathogenesis ofEnterococcus faeciumin exper-imental endocarditis. Infect. Immun.76:4120–4128.
26.Nallapareddy, S. R., K. V. Singh, P. C. Okhuysen, and B. E. Murray.2008. A functional collagen adhesin gene,acm, in clinical isolates ofEnterococcus faeciumcorrelates with the recent success of this emerging nosocomial pathogen. Infect. Immun.76:4110–4119.
27.Rice, L. B., L. Carias, S. Rudin, C. Vael, H. Goossens, C. Konstabel, I. Klare S.R. Nallapareddy, W. Huang, and B. E. Murray.2003. A potential virulence gene, hylEfm, predominates inEnterococcus faeciumof clinical origin. J.
In-fect. Dis.187:508–512.
28.Rice, L. B., V. Lakticov, L. Carias, S. Rudin, R. Hutton, and S. H. Marshall.
2009. Transferable capacity for gastrointestinal colonization inEnterococcus faeciumin a mouse model. J. Infect. Dis.199:342–349.
29.Ruiz-Garbajosa, P., M. J. Bonten, D. A. Robinson, J. Top, S. R. Nallapa-reddy, C. Torres, T. M. Coque, R. Canto´n, F. Baquero, B. E. Murray, R. del Campo, and R. J. Willems.2006. Multilocus sequence typing scheme for
Enterococcus faecalisreveals hospital-adapted genetic complexes in a back-ground of high rates of recombination. J. Clin. Microbiol.44:2220–2228. 30.Schneewind, O., P. Model, and V. A. Fischetti.1992. Sorting of protein A to
the staphylococcal cell wall. Cell70:267–281.
31.Sillanpa¨a¨, J., S. R. Nallapareddy, V. P. Prakash, X. Qin, M. Ho¨o¨k, G. M. Weinstock, and B. E. Murray.2008. Identification and phenotypic charac-terization of a second collagen adhesin, Scm, and genome-based identifica-tion and analysis of 13 other predicted MSCRAMMs, including four distinct pilus loci, inEnterococcus faecium. Microbiology154:3199–3211. 32.Sillanpa¨a¨, J., V. P. Prakash, S. R. Nallapareddy, and B. E. Murray.2009.
Distribution of genes encoding MSCRAMMs and pili in clinical and natural populations ofEnterococcus faecium. J. Clin. Microbiol.47:896–901. 33.Singh, K. V., T. M. Coque, G. M. Weinstock, and B. E. Murray.1998. In vivo
testing of anEnterococcus faecalis efaAmutant and use ofefaAhomologs for species identification. FEMS Immunol. Med. Microbiol.21:323–331. 34.Singh, K. V., R. J. Lewis, and B. E. Murray.2009. Importance of theepa
locus ofEnterococcus faecalisOG1RF in a mouse model of ascending uri-nary tract infection. J. Infect. Dis.200:417–420.
35.Tenover, F. C., R. D. Arbeit, R. V. Goering, P. A. Mickelsen, B. E. Murray, D. H. Persing, and B. Swaminathan.1995. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J. Clin. Microbiol.33:2233–2239.
36.Ton-That, H., and O. Schneewind.2003. Assembly of pili on the surface of
Corynebacterium diphtheriae. Mol. Microbiol.50:1429–1438.
37.Treitman, A. N., P. R. Yarnold, J. Warren, and G. A. Noskin.2005. Emerging incidence ofEnterococcus faeciumamong hospital isolates (1993 to 2002). J. Clin. Microbiol.43:462–463.
38.Willems, R. J., W. Homan, J. Top, M. van Santen-Verheuvel, D. Tribe, X. Manzioros, C. Gaillard, C. M. Vandenbroucke-Grauls, E. M. Mascini, E. van Kregten, J. D. van Embden, and M. J. Bonten.2001. Variantespgene as a marker of a distinct genetic lineage of vancomycin-resistantEnterococcus faeciumspreading in hospitals. Lancet357:853–855.
39.Willems, R. J., J. Top, M. van Santen, D. A. Robinson, T. M. Coque, F. Baquero, H. Grundmann, and J. M. Bonten.2005. Global spread of vanco-mycin resistantEnterococcus faeciumfrom distinct nosocomial genetic com-plex. Emerg. Infect. Dis.11:821–828.