Positive Rods: Development of a Diagnostic Algorithm for the Clinical
Laboratory
Bettina Schulthess, Guido V. Bloemberg, Reinhard Zbinden, Erik C. Böttger, Michael Hombach Institute of Medical Microbiology, University of Zurich, Zurich, Switzerland
Reported matrix-assisted laser desorption ionization–time of flight mass spectrometry (MALDI-TOF MS) identification rates of Gram-positive rods (GPR) are low compared to identification rates of Gram-positive cocci. In this study, three sample prepara-tion methods were compared for MALDI-TOF MS identificaprepara-tion of 190 well-characterized GPR strains: direct transfer, direct transfer-formic acid preparation, and ethanol-formic acid extraction. Using the interpretation criteria recommended by the manufacturer, identification rates were significantly higher for direct transfer-formic acid preparation and ethanol-formic acid extraction than for direct transfer. Reducing the species cutoff from 2.0 to 1.7 significantly increased species identification rates. In a subsequent prospective study, 215 clinical GPR isolates were analyzed by MALDI-TOF MS, and the results were compared to those for identification using conventional methods, with discrepancies being resolved by 16S rRNA andrpoBgene analysis. Us-ing the direct transfer-formic acid preparation and a species cutoff of 1.7, congruencies on the genus and species levels of 87.4% and 79.1%, respectively, were achieved. In addition, the rate of nonidentified isolates dropped from 12.1% to 5.6% when using an extended database, i.e., the Bruker database amended by reference spectra of the 190 GPR of the retrospective study. Our data demonstrate three ways to improve GPR identification by the Bruker MALDI Biotyper, (i) optimize sample preparation using formic acid, (ii) reduce cutoff scores for species identification, and (iii) expand the database. Based on our results, we suggest an identification algorithm for the clinical laboratory combining MALDI-TOF MS with nucleic acid sequencing.
T
raditionally, identification of Gram-positive rods (GPR) in clinical diagnostic laboratories is based on morphological and biochemical criteria (1). During the last 2 decades, molecular ap-proaches such as 16S rRNA gene sequence analysis have been im-plemented to complement or replace conventional identification algorithms (1). However, molecular techniques remain costly and are available in only a few routine clinical laboratories. Recently, matrix-assisted laser desorption ionization–time of flight mass spectrometry (MALDI-TOF MS) emerged as a novel and cost-saving approach in diagnostic microbiology for the identification of bacteria and fungi (2,3). Different MALDI-TOF MS systems for microbial identification have been established, including the MALDI Biotyper (Bruker Daltonik GmbH, Bremen, Germany), Vitek MS (bioMérieux, Marcy l’Etoile, France), and the Andro-mas system (AndroAndro-mas SAS, Paris, France). MALDI-TOF MS sys-tems are based on the acquisition of a mass spectrum of an un-known organism and its comparison with reference databases for identification (4–6). The systems differ mainly in the sample prep-aration procedures, the species coverage of the reference database, and the identification algorithm of the software (7–10).Most studies addressing the use of MALDI-TOF MS for the identification of bacteria in the diagnostic laboratory have focused on the analysis of Gram-negative bacteria and Gram-positive cocci and included only a few GPR (2,3,7,9–12). Identification by MALDI-TOF MS has been analyzed for single genera of GPR, such asCorynebacterium(13–15),Actinomyces(16),Nocardia(17), Lis-teria(18), anaerobic GPR (19), and difficult-to-identify GPR (20, 21). These studies were mostly done using the Bruker MALDI Biotyper system and indicated that sample preparation by the di-rect transfer procedure without additional modification is not suf-ficient for accurate identification of GPR but that a chemical ex-traction method is required. This exex-traction method, however,
involves additional preparatory steps, significantly increasing pro-cessing time (22), and seems, therefore, not suitable for high-throughput applications as required in clinical diagnostic labora-tories. An adapted direct transfer method with on-target formic acid treatment leading toin situcell lysis holds promise for in-creased identification rates without time-consuming extraction (23). Similar identification rates were reported for Corynebacte-rium spp. and anaerobic bacteria when on-target formic acid preparation and extraction were compared (19,24). An alterna-tive preparation method with on-target ethanol treatment was described by Farfour et al. (25), who analyzed a large collection of GPR using the Andromas system and reported accurate species identification. Few studies have systematically compared different sample preparation methods for the identification of GPR with MALDI-TOF MS (13,24,26).
In this study, we evaluated the MALDI Biotyper (Bruker Dal-tonik) for identification of clinical GPR strains. In the first, retro-spective part of the study, we analyzed 190 well-characterized GPR strains from our strain collection, including 64 species from 21 genera. We compared identification rates of three sample
prepa-Received1 September 2013Returned for modification12 October 2013
Accepted14 January 2014
Published ahead of print22 January 2014
Editor:R. Patel
Address correspondence to Bettina Schulthess, [email protected].
Supplemental material for this article may be found athttp://dx.doi.org/10.1128
/JCM.02399-13.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JCM.02399-13
on May 16, 2020 by guest
http://jcm.asm.org/
ration methods (direct transfer, direct transfer-formic acid prep-aration with on-target formic acid treatment, and ethanol-formic acid extraction) using standard interpretation criteria of the man-ufacturer and individual species and genus identification cutoff values. Based on the data from the retrospective study, we selected the direct transfer-formic acid preparation for the second, pro-spective part of the study, where we compared MALDI-TOF MS-based identification with the current identification algorithm used in our laboratory (n⫽215 isolates) (1). We defined reliable spe-cies and genus identifications for GPR, and we propose a practical algorithm of GPR identification for the routine laboratory inte-grating MALDI-TOF MS-based identification.
MATERIALS AND METHODS
Bacterial strains and culture conditions.For the retrospective part of the study, we selected 190 Gram-positive rods (GPR) from the institute’s strain collection, including the clinically most relevant genera, i.e., Acti-nomyces(n⫽45 strains) andCorynebacterium(n⫽81 strains), as well as more rarely encountered genera (n⫽64 strains), such asActinobaculum,
Dermabacter,Nocardia,Rothia, andTrueperella(see Table S1 in the sup-plemental material). The strains were characterized by phenotypic meth-ods and 16S rRNA gene analysis, which was considered the gold standard for identification. ForCorynebacteriumspp., additional rpoBgene se-quencing was done when the discriminatory power of the biochemical testing and 16S rRNA gene analysis was insufficient (see the identification ofCorynebacteriumspp. below). In the prospective part of the study (Feb-ruary 2012 to September 2012), a total of 215 clinical GPR isolates were identified in parallel by (i) MALDI-TOF MS and (ii) our conventional identification algorithm (1). Per this algorithm, identification was based on phenotypic characteristics only for 144 out of 215 isolates; for 71 out of 215 isolates, additional 16S rRNA gene sequence analyses were done. Bac-teria were routinely cultivated on Columbia agar containing 5% sheep blood (bioMérieux) at 37°C with 7.5% CO2for 24 to 48 h for MALDI-TOF MS measurement.Nocardiaspp. were grown aerobically at 30°C, and
Propionibacteriumspp. were grown under anaerobic conditions.
Identification ofCorynebacteriumspecies.Species assignments by 16S rRNA gene analysis ofCorynebacteriumspp. with high homology of 16S rRNA gene sequences (27), i.e.,C.afermentans/C.coyleae/C. mucifa-ciens,C.aurimucosum/C. minutissimum/C.singulare,C.macginleyi/C. ac-colens, andC.propinquum/C. pseudodiphtheriticum, were confirmed by biochemical testing. When the biochemical reaction pattern did not allow unambiguous species assignment according toBergey’s Manual of System-atic Bacteriology(27),rpoBsequence analysis was done.rpoBanalysis con-firmed the identifications by 16S rRNA gene analysis for all isolates, except oneC.mucifaciensisolate which was more closely related toC. afermen-tansbyrpoBsequence analysis, and thus, was assigned toCorynebacterium
sp. only.
Phenotypic identification.GPR were identified by means of bio-chemical reactions according to those reported by von Graevenitz and Funke (28), including catalase; acid production from glucose, maltose, sucrose, mannitol, and xylose in semisolid cysteine-Trypticase agar me-dium; motility; nitrate reduction; hydrolysis of urea; hydrolysis of esculin; the CAMP test; and a test for lipophilicity.
16S rRNA gene analysis.Identification by partial 16S rRNA gene se-quence analysis was done according to CLSI guidelines (29) and as de-scribed by Bosshard et al. (1). A 16S rDNA fragment, corresponding to
Escherichia colipositions 10 to 806, was amplified using primers BAK11w (5=-AGTTTGATC[A/C]TGGCTCAG) and BAK2 (5=-GGACTAC[C/T/A] AGGGTATCTAAT), and sequenced with the forward primer BAK11w (1). For identification, the following criteria were used: (i) species identi-fication when the determined sequence had a similarity score ofⱖ99% with that of a reference sequence of a classified species, (ii) genus assign-ment when the similarity score was⬍99% andⱖ95%, and (iii) family assignment when the similarity score was⬍95% (1).
rpoBgene analysis.Sequence analysis of the partialrpoBgene of Co-rynebacteriumspp. was done as previously described (30). Species assign-ment requiredⱖ95% sequence similarity (31).
Sample preparation for MALDI-TOF MS.Preparations of bacterial isolates for MALDI-TOF MS measurement were done as previously de-scribed (22,23). Briefly, for the direct transfer method, fresh colony ma-terial was smeared on a polished steel MSP 96 target (Bruker Daltonik) using a toothpick, overlaid with 1l of a saturated a-cyano-4-hydroxy-cinnamic acid (HCCA) matrix solution in 50% acetonitrile-2.5% trifluo-roacetic acid (Bruker Daltonik), and air dried at room temperature. For the direct transfer-formic acid method, 1l of 70% formic acid was added to the bacterial spot and allowed to air dry before the matrix solution was added. For the ethanol-formic acid extraction procedure, a loopful of bacterial material was suspended in 300l distilled water, and 900l ethanol was added. The cell suspension was centrifuged at 17,000⫻gfor 2 min, and the supernatant was discarded. The centrifugation was re-peated, and the residual ethanol was discarded. The pellet was air dried and thoroughly resuspended in 5 to 50l formic acid-water (70:30 [vol/ vol]) depending on the size, and finally, an equal volume of acetonitrile was added. After centrifugation at 17,000⫻gfor 2 min, 1l of the supernatant was transferred to the MALDI target plate and allowed to dry at room temperature before being overlaid with 1l of matrix solution.
MALDI-TOF MS analysis.The acquisition and analysis of mass spec-tra were performed by a Microflex LT mass spectrometer (Bruker Dal-tonik) using the MALDI Biotyper software package (version 3.0) with the reference database version 3.1.2.0 (3,995 database entries) (Bruker Dal-tonik) and default parameter settings as published previously (32). The Bruker bacterial test standard (Bruker Daltonik) was used for calibration according to the instructions of the manufacturer. For each strain, two preparations of colony/sample material were analyzed.
MALDI-TOF MS data interpretation.The Biotyper software
com-pares each sample mass spectrum to the reference mass spectra in the database, calculates an arbitrary unit score value between 0 and 3 reflect-ing the similarity between the sample and the reference spectrum, and displays the top 10 matching database records. Standard Bruker interpre-tative criteria were applied (32). Briefly, scores ofⱖ2.0 were accepted for species assignment and scores ofⱖ1.7 but⬍2.0 for identification to the genus level. Scores below 1.7 were considered unreliable. Variations of the cutoff score values were done by reducing the species cutoff values to 1.9, 1.8, and 1.7, and the genus cutoff values to 1.6 and 1.5, followed by rein-terpreting the top 10 matching database records.
Generation of in-house reference database.Reference spectra were created for all 190 clinical strains of the retrospective study and added to the Bruker database version 3.1.2.0. For each strain, a set of 24 spectra was measured and checked manually for flat-line, outlier, and single spectra with peaks differing form the other spectra. Questionable spectra were removed, and a total of 20 to 24 spectra were used to calculate a reference spectrum, using the automated function of the Biotyper software.
Discrepancy analysis.Discrepancies between MALDI-TOF MS and
phenotypic identification were resolved by 16S rRNA gene andrpoBgene sequence analyses, which were considered the gold standards for identifi-cation (1,30,31).
Statistical analysis.Statistical calculations were done using IBM SPSS Statistics software, version 20 (SPSS, Inc., Chicago, IL). Overall differ-ences among the three MALDI-TOF MS preparation methods (direct transfer, direct transfer-formic acid preparation, and ethanol-formic acid extraction) and the tested cutoff score values were evaluated using the Kruskal-Wallis test. Follow-up tests were conducted using the Mann-Whitney U test for pairwise comparison of the three preparation methods and the different cutoff score values. Chance agreement between direct transfer-formic acid preparation and ethanol-formic acid extraction was evaluated with an interrater reliability analysis using the Kappa statistics (33). Differences were considered statistically significant atPvalues of ⬍0.05.
on May 16, 2020 by guest
http://jcm.asm.org/
Nucleotide sequence accession numbers.16S rRNA gene sequences of the retrospective and prospective parts of the study are accessible at GenBank under accession numbersKF925884toKF926073andKJ081453 toKJ081536.rpoBgene sequences are accessible at GenBank under acces-sion numbersKJ081537toKJ081544andKJ150303toKJ150321.
RESULTS
Retrospective study. (i) MALDI Biotyper identification rates of Gram-positive rods (GPR) are increased by formic acid treat-ment.We evaluated the Bruker MALDI Biotyper system by ana-lyzing 190 well-characterized clinical GPR isolates, including 64 species from 21 genera (see Table S1 in the supplemental mate-rial). When applying the direct transfer sample preparation method and the standard interpretation criteria of the
manufac-turer, i.e., a species cutoff score value of 2.0 and a genus cutoff score value of 1.7, the MALDI Biotyper correctly identified 137 of 190 strains (72.1%) at the genus level (Table 1). Correct identifi-cation at the species level was achieved for 70 of 190 strains (36.8%). Genus and species identification rates significantly in-creased to 84.7% and 61.1%, respectively, when we used direct transfer-formic acid sample preparation with on-target formic acid treatment (Z⫽ ⫺4.63,P⬍0.001, andZ⫽ ⫺2.99,P⫽0.003, for genus and species identification of direct transfer and direct transfer-formic acid preparation, respectively). Comparable in-creases in identification rates to 87.4% and 62.1% for genus and species identifications, respectively, were observed for the etha-nol-formic acid extraction. The differences between direct
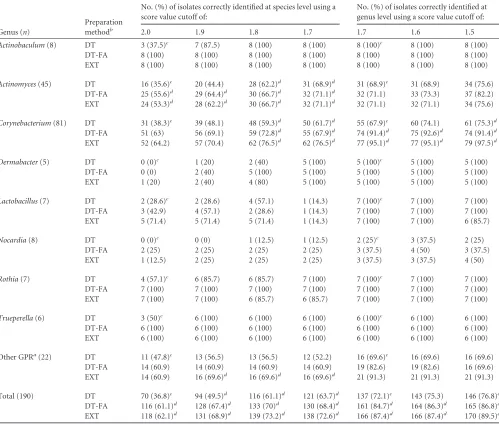
trans-TABLE 1Retrospective analysis of Gram-positive rods with MALDI-TOF MS: comparison of three sample preparation methods
Genus (n)
Preparation methodb
No. (%) of isolates correctly identified at species level using a score value cutoff of:
No. (%) of isolates correctly identified at genus level using a score value cutoff of:
2.0 1.9 1.8 1.7 1.7 1.6 1.5
Actinobaculum(8) DT 3 (37.5)c 7 (87.5) 8 (100) 8 (100) 8 (100)c 8 (100) 8 (100)
DT-FA 8 (100) 8 (100) 8 (100) 8 (100) 8 (100) 8 (100) 8 (100)
EXT 8 (100) 8 (100) 8 (100) 8 (100) 8 (100) 8 (100) 8 (100)
Actinomyces(45) DT 16 (35.6)c 20 (44.4) 28 (62.2)d 31 (68.9)d 31 (68.9)c 31 (68.9) 34 (75.6)
DT-FA 25 (55.6)d 29 (64.4)d 30 (66.7)d 32 (71.1)d 32 (71.1) 33 (73.3) 37 (82.2)
EXT 24 (53.3)d 28 (62.2)d 30 (66.7)d 32 (71.1)d 32 (71.1) 32 (71.1) 34 (75.6)
Corynebacterium(81) DT 31 (38.3)c 39 (48.1) 48 (59.3)d 50 (61.7)d 55 (67.9)c 60 (74.1) 61 (75.3)d
DT-FA 51 (63) 56 (69.1) 59 (72.8)d 55 (67.9)d 74 (91.4)d 75 (92.6)d 74 (91.4)d
EXT 52 (64.2) 57 (70.4) 62 (76.5)d 62 (76.5)d 77 (95.1)d 77 (95.1)d 79 (97.5)d
Dermabacter(5) DT 0 (0)c 1 (20) 2 (40) 5 (100) 5 (100)c 5 (100) 5 (100)
DT-FA 0 (0) 2 (40) 5 (100) 5 (100) 5 (100) 5 (100) 5 (100)
EXT 1 (20) 2 (40) 4 (80) 5 (100) 5 (100) 5 (100) 5 (100)
Lactobacillus(7) DT 2 (28.6)c 2 (28.6) 4 (57.1) 1 (14.3) 7 (100)c 7 (100) 7 (100)
DT-FA 3 (42.9) 4 (57.1) 2 (28.6) 1 (14.3) 7 (100) 7 (100) 7 (100)
EXT 5 (71.4) 5 (71.4) 5 (71.4) 1 (14.3) 7 (100) 7 (100) 6 (85.7)
Nocardia(8) DT 0 (0)c 0 (0) 1 (12.5) 1 (12.5) 2 (25)c 3 (37.5) 2 (25)
DT-FA 2 (25) 2 (25) 2 (25) 2 (25) 3 (37.5) 4 (50) 3 (37.5)
EXT 1 (12.5) 2 (25) 2 (25) 2 (25) 3 (37.5) 3 (37.5) 4 (50)
Rothia(7) DT 4 (57.1)c 6 (85.7) 6 (85.7) 7 (100) 7 (100)c 7 (100) 7 (100)
DT-FA 7 (100) 7 (100) 7 (100) 7 (100) 7 (100) 7 (100) 7 (100)
EXT 7 (100) 7 (100) 6 (85.7) 6 (85.7) 7 (100) 7 (100) 7 (100)
Trueperella(6) DT 3 (50)c 6 (100) 6 (100) 6 (100) 6 (100)c 6 (100) 6 (100)
DT-FA 6 (100) 6 (100) 6 (100) 6 (100) 6 (100) 6 (100) 6 (100)
EXT 6 (100) 6 (100) 6 (100) 6 (100) 6 (100) 6 (100) 6 (100)
Other GPRa(22) DT 11 (47.8)c 13 (56.5) 13 (56.5) 12 (52.2) 16 (69.6)c 16 (69.6) 16 (69.6)
DT-FA 14 (60.9) 14 (60.9) 14 (60.9) 14 (60.9) 19 (82.6) 19 (82.6) 16 (69.6)
EXT 14 (60.9) 16 (69.6)d 16 (69.6)d 16 (69.6)d 21 (91.3) 21 (91.3) 21 (91.3)
Total (190) DT 70 (36.8)c 94 (49.5)d 116 (61.1)d 121 (63.7)d 137 (72.1)c 143 (75.3) 146 (76.8)d
DT-FA 116 (61.1)d 128 (67.4)d 133 (70)d 130 (68.4)d 161 (84.7)d 164 (86.3)d 165 (86.8)d EXT 118 (62.1)d 131 (68.9)d 139 (73.2)d 138 (72.6)d 166 (87.4)d 166 (87.4)d 170 (89.5)d a
Genera withⱕ5 isolates tested, includingAlloscardovia,Arcanobacterium,Arthrobacter,Bifidobacterium,Brevibacterium,Dietzia,Gardnerella,Gordonia,Listeria,Paenibacillus, Propionibacterium,Rhodococcus, andTerrabacter(for details, see Table S2 in the supplemental material).
b
MALDI-TOF MS preparation method; DT, direct transfer; DT-FA, direct transfer with formic acid treatment; EXT, ethanol-formic acid extraction.
cReference value. d
Value significantly differs from the reference value. Agreement between the reference method (direct transfer species cutoff value 2.0, genus cutoff value 1.7) and the method evaluated was compared using a two-tailed McNemar test for paired samples. Differences were considered statistically significant atPvalues of⬍0.05.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:3.585.43.542.78.502.2]fer-formic acid preparation and ethanol-formic extractions were not significant (Z⫽ ⫺0.37,P⫽0.710, andZ⫽ ⫺0.74,P⫽0.460, for the comparison of species and genus identification, respec-tively). In order to calculate the strength of agreement between these two methods, kappa statistical analysis was performed. The kappa coefficient for the direct transfer-formic acid method and the ethanol-formic acid extraction was 0.81 (P⬍0.001, 95% con-fidence interval [CI], 0.73, 0.88). This indicates almost perfect agreement between these two methods (33).
Genus and species identification rates varied between different genera (Table 1; also see Table S2 in the supplemental material). ForActinobaculum,Dermabacter,Lactobacillus,Rothia, and True-perella, genus identification was achieved for all strains tested, independently of the sample preparation method. In contrast, for theNocardia strains, genus identification rates varied between 25.0% and 37.5% for the different preparation methods. Direct transfer-formic acid preparation and ethanol-formic acid extrac-tion increased the average MALDI score values by 0.15 and 0.17 score units, respectively, as compared to direct transfer. No mass spectra (no peaks) were detected in 4.6% and 1.6% of the mea-surements when direct transfer or direct transfer-formic acid preparation was used, respectively. The majority of no-peaks re-sults were observed forNocardiaspp. and two distinct species of the genusCorynebacterium, i.e.,C.mucifaciensandC. pyruvicip-roducens. No-peaks results were not observed for the ethanol-for-mic acid extraction procedure.
(ii) Species coverage of the Bruker reference database. Spe-cies of 21 genera covered by 904 reference entries in the Bruker database version 3.1.2.0 were analyzed in the retrospective part of the study. Four species tested in the present work were not in-cluded in the database, i.e.,Actinomyces johnsonii, Corynebacte-rium pyruviciproducens, “CorynebacteCorynebacte-rium pseudogenitalium,”and Paenibacillus campinasensis. WhileA. johnsoniiandP. campina-sensis were not identified by MALDI-TOF MS, most of theC. pyruviciproducens and “C. pseudogenitalium” strains were as-signed to the genusCorynebacterium when the direct transfer-formic acid or the ethanol-transfer-formic acid procedure was used.
(iii) Low discrimination at the species level using the MALDI Biotyper.For some isolates, MALDI-TOF MS identification rank lists showed scores ofⱖ2.0 for⬎1 species, resulting in species inconsistency and identification to the genus level only. Such low discrimination at the species level was observed for the following strains:Corynebacterium aurimucosum(scores wereⱖ2.0 forC. aurimucosum/minutissimum), Corynebacterium simulans(scores wereⱖ2.0 forC.simulans/striatum),Lactobacillus gasseri(scores wereⱖ2.0 for L. gasseri/johnsonii), and Listeria monocytogenes (scores wereⱖ2.0 forL. monocytogenes/ivanovii/innocua). We ob-served low species discrimination more frequently when applying direct transfer-formic acid preparation (2.6%) and ethanol-formic acid extraction (2.6%) but less frequently when using the direct trans-fer method (1.6%). In all cases, the species with the highest score corresponded to the identification determined by phenotypic and molecular methods representing the gold standard.
(iv) Misidentifications using the MALDI Biotyper. MALDI-TOF MS analysis ofRhodococcus gordoniae,Rhodococcus coryne-bacterioides,Terrabacter tumescens, andGordonia terraeregularly produced poor-quality or no mass spectra, preventing identifica-tion. In addition, identification asArthrobacter castelliwas occa-sionally observed for these strains. The latter discrepancy is appar-ently due to the matching of a poor-quality spectrum with the
reference spectrum ofA.castelli. OneRhodococcus gordoniaestrain (species assignment by 16S rRNA gene analysis) was identified by
MALDI-TOF MS asRhodococcus rhodochrous.
(v) Individual cutoff score values for species and genus iden-tification.Standard cutoff score values for species and genus iden-tification are set to 2.0 and 1.7 by the manufacturer. We evaluated the effects of reducing the species and genus cutoff values from 2.0 to 1.9, 1.8, and 1.7, and from 1.7 to 1.6 and 1.5, respectively (Table 1; also see Table S2 in the supplemental material). Overall, reduc-tion of the genus cutoff value marginally increased identificareduc-tion rates. The cutoff reduction was accompanied by increasing num-bers of low-discrimination results at the genus level and by more misidentifications (low-discrimination rates of 0% versus 2% and misidentification rates of 0% versus 1% when applying genus-level cutoff scores of 1.7 and 1.5, respectively). In particular, Acti-nomycesisolates were identified asAcinetobactersp.,Arthrobacter sp., orPseudomonassp., and one Corynebacteriumisolate was as-signed to the genusClostridium.
Significantly higher species identification rates were observed for all three preparation methods when lower species cutoff values were applied (Table 1; also see Table S2 in the supplemental ma-terial). Reducing the species cutoff from 2.0 to 1.7 increased iden-tification rates from 36.8% to 63.7%, from 61.1% to 68.4%, and from 62.1% to 71.6% for direct transfer, direct transfer-formic acid preparation, and ethanol-formic acid extraction, respec-tively. Using a cutoff of 1.7, the species identification rates of direct transfer, direct transfer-formic acid preparation, and ethanol-for-mic acid extraction no longer differed significantly (2[2,n⫽ 570]⫽5.65,P⫽0.059, with the Kruskal-Wallis test). However, at the same time, the rate of low-discrimination results increased from 1.6% to 5.8% (species cutoff of 2.0 versus 1.7) for direct transfer, and from 2.6% to 10% for direct transfer-formic acid preparation and ethanol-formic acid extraction. The higher rates of low discrimination at the species level were mainly due to less differentiation betweenC.aurimucosum/minutissimum,C. stria-tum/simulans,L. gasseri/johnsonii,L. rhamnosus/casei,L. monocy-togenes/innocua, and Rhodococcus corynebacterioides/kroppenst-edtii. Reducing the species cutoff from 2.0 to 1.7 increased the rate of misidentifications from 0.5% to 5% for all three methods. The increase in misidentification rates was caused by (i) the misiden-tification ofRhodococcus corynebacterioidesasRhodococcus krop-penstedtiiand (ii) the misidentification of “C. pseudogenitalium” as “Corynebacterium lipophile.”
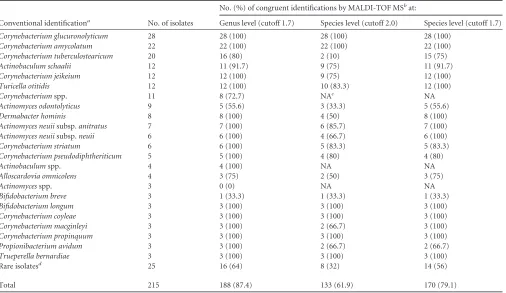
Prospective study. (i) Comparison of MALDI-TOF MS iden-tification with the conventional ideniden-tification algorithm (phe-notypic, genotypic) in the routine diagnostic laboratory.In a prospective study, 215 clinically relevant GPR isolates, including 13 genera and 36 species, were identified by MALDI-TOF MS and according to the conventional identification algorithm as previ-ously published (1). The MALDI Biotyper system was used with the direct transfer-formic acid sample preparation method apply-ing the Bruker database version 3.1.2.0. (3,995 entries), and a spe-cies-level identification cutoff score value of 2.0 (Table 2). Overall, for 188 of 215 isolates (87.4%), congruence on the genus level was observed between conventional and MALDI-TOF MS identifica-tion. Of 215 isolates, 133 (61.9%) yielded concordant species identification, and 26 of 215 isolates (12.1%) were not identified by MALDI-TOF MS.
Spectra that did not yield a scoreⱖ2.0 were reanalyzed using a species cutoff of 1.7. This significantly increased the total number
on May 16, 2020 by guest
http://jcm.asm.org/
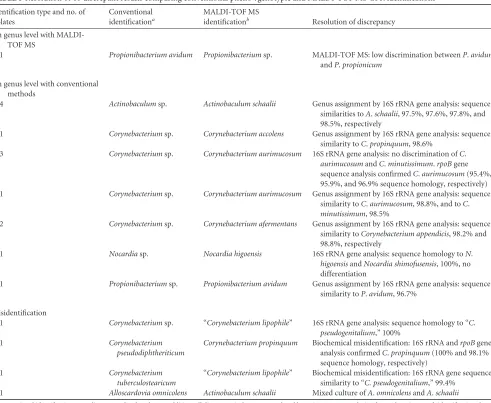
of congruent species results of conventional and MALDI-TOF MS identification from 133 (61.9%) to 170 (79.1%) (Table 2). In par-ticular, a major increase of the identification rate from 10% to 75% was observed forCorynebacterium tuberculostearicum. We observed discrepancies on the genus and/or species level for 18 isolates (8.4%) when using a species cutoff of 1.7 (Table 3). For 3 of these 18 isolates, identification by molecular reference methods (16S rRNA gene and/orrpoBgene sequencing) confirmed species identification by MALDI-TOF MS. For 11 out of 18 isolates, MALDI-TOF MS allowed assignment to the species level, while 16S rRNA gene sequence analysis allowed assignment to the genus level only. Two biochemical misidentifications were observed. OneC.propinquumwas misidentified asC. pseudodiphtheriticum, and one “C. pseudogenitalium” was misidentified asC. tuberculo-stearicum. For onePropionibacterium avidum, low discrimination ofP. avidumandPropionibacterium propionicumby MALDI-TOF MS was observed.
One discrepancy on the genus level was observed (MALDI-TOF MS identification,Actinobaculum schaalii; 16S rRNA gene analysis, Alloscardovia omnicolens). Closer analysis revealed the presence of a mixed culture ofA.schaaliiandA.omnicolens.
Application of our amended Bruker-IMM database, i.e., an extended database combining the commercially available Bruker database with the in-house-generated reference spectra from the
retrospective study, increased the number of congruent genus and species identifications from 188 to 202 (94.0%) and from 170 to 180 (83.7%), respectively. The rate of nonidentifications was re-duced to 5.6%. In particular, 14 isolates that were previously not
identified by MALDI-TOF MS were identified as Actinomyces
odontolyticus(3 isolates),Actinomyces urogenitalis(1 isolate), Acti-nomycessp. (3 isolates),Bifidobacterium breve(1 isolate), Coryne-bacterium imitans(1 isolate),Corynebacterium pyruviciproducens (1 isolate), andCorynebacterium tuberculostearicum(4 isolates).
DISCUSSION
MALDI-TOF MS is increasingly used in diagnostic laboratories for the identification of bacteria and fungi. We have analyzed the performance of the Bruker MALDI Biotyper system for the iden-tification of GPR and evaluated possible ways to improve its per-formance, i.e., different sample preparation methods, data inter-pretation algorithms (particularly the variation of score cutoff values), and extension of the database by proprietary reference entries.
[image:5.585.42.548.77.370.2]Sample preparation.Using the manufacturer’s standard inter-pretation criteria, i.e., a genus cutoff of 1.7 and a species cutoff of 2.0 in combination with the direct transfer method, identification rates of 72.1%, and 36.8% were achieved at the genus and species levels, respectively. Treatment of the sample with formic acid by
TABLE 2MALDI-TOF MS identification versus conventional identification for 215 GPR clinically relevant isolates
Conventional identificationa No. of isolates
No. (%) of congruent identifications by MALDI-TOF MSbat:
Genus level (cutoff 1.7) Species level (cutoff 2.0) Species level (cutoff 1.7)
Corynebacterium glucuronolyticum 28 28 (100) 28 (100) 28 (100)
Corynebacterium amycolatum 22 22 (100) 22 (100) 22 (100)
Corynebacterium tuberculostearicum 20 16 (80) 2 (10) 15 (75)
Actinobaculum schaalii 12 11 (91.7) 9 (75) 11 (91.7)
Corynebacterium jeikeium 12 12 (100) 9 (75) 12 (100)
Turicella otitidis 12 12 (100) 10 (83.3) 12 (100)
Corynebacteriumspp. 11 8 (72.7) NAc NA
Actinomyces odontolyticus 9 5 (55.6) 3 (33.3) 5 (55.6)
Dermabacter hominis 8 8 (100) 4 (50) 8 (100)
Actinomyces neuiisubsp.anitratus 7 7 (100) 6 (85.7) 7 (100)
Actinomyces neuiisubsp.neuii 6 6 (100) 4 (66.7) 6 (100)
Corynebacterium striatum 6 6 (100) 5 (83.3) 5 (83.3)
Corynebacterium pseudodiphtheriticum 5 5 (100) 4 (80) 4 (80)
Actinobaculumspp. 4 4 (100) NA NA
Alloscardovia omnicolens 4 3 (75) 2 (50) 3 (75)
Actinomycesspp. 3 0 (0) NA NA
Bifidobacterium breve 3 1 (33.3) 1 (33.3) 1 (33.3)
Bifidobacterium longum 3 3 (100) 3 (100) 3 (100)
Corynebacterium coyleae 3 3 (100) 3 (100) 3 (100)
Corynebacterium macginleyi 3 3 (100) 2 (66.7) 3 (100)
Corynebacterium propinquum 3 3 (100) 3 (100) 3 (100)
Propionibacterium avidum 3 3 (100) 2 (66.7) 2 (66.7)
Trueperella bernardiae 3 3 (100) 3 (100) 3 (100)
Rare isolatesd 25 16 (64) 8 (32) 14 (56)
Total 215 188 (87.4) 133 (61.9) 170 (79.1)
a
Identification combining biochemical methods and 16S rRNA gene sequence analysis according to Bosshard et al. (1).
bMALDI-TOF MS identification applying direct transfer-formic acid preparation, a genus cutoff value of 1.7, and species cutoff values of 2.0 and 1.7, respectively with the Bruker
database version 3.1.2.0 (containing 3995 entries).
cNA, not applicable. d
Rare isolates (withnⱕ2), includingActinomycesspp. (A.funkei,A.israelii,A.radingae,A.turicensis,A.urogenitalis, andA.viscosus),Brevibacteriumspp. (B.ravenspurgense), Corynebacteriumspp. (C.accolens,C.afermentans,C.aurimucosum/minutissimum,C.bovis,C.imitans,C.mastitidis,C.mucifaciens, andC.ureicelerivorans),Nocardiaspp., Paenibacillusspp., andPropionibacteriumspp. (P.acnes).
on May 16, 2020 by guest
http://jcm.asm.org/
either on-target overlay or tube-based extraction significantly in-creased genus and species identification rates to approximately 85% and 60%, respectively (Table 1). Identification rates for the direct transfer-formic acid method were comparable to those of the ethanol-formic acid extraction procedure that is considered the gold standard and which is used to generate the reference database. These findings are in agreement with previous reports which indicated that on-target treatment with formic acid or eth-anol-formic acid extraction improved identification of GPR by MALDI-TOF MS (13,14,24,26). However, the ethanol-formic acid procedure includes various manual preparation steps and is time-consuming (22). In contrast, total time to result is only mod-erately increased by the direct transfer-formic acid method com-pared to direct transfer (32), and thus seems more suited for rou-tine use in diagnostic laboratories. While this study was in progress, Farfour et al. analyzed a large collection of GPR using the Andromas system (25) and reported accurate species identifica-tion of GPR using direct transfer with addiidentifica-tional ethanol treat-ment.
[image:6.585.49.540.73.476.2]Individual data interpretation algorithms.Reducing the ge-nus cutoff value from 1.7 to 1.5 only slightly increased overall genus identification rates for direct transfer and ethanol-formic acid extraction (from 72.1% to 76.3% and from 87.4% to 89.5%, respectively), while the identification rate for direct transfer-for-mic acid preparation remained constant at 84.7% (Table 1). At the same time, a genus cutoff score value of 1.5 resulted in an in-creased misidentification of 1% of the isolates. In contrast to ge-nus identification, species identification was significantly en-hanced by reducing the species cutoff value from 2.0 to 1.7 independent of the preparation method applied (36.8% versus 63.7%, 61.1% versus 68.4%, and 62.1% versus 71.6% for direct transfer, direct transfer-formic acid preparation, and ethanol-for-mic acid extraction, respectively). These results are in agreement with previous studies on MALDI-TOF MS identification of GPR, such asCorynebacterium spp., anaerobic GPR, and difficult-to-identify GPR, which all support the application of a lower species cutoff (mostly 1.7) to increase the MALDI-TOF MS identification rate (13,19–21,26).
TABLE 3Resolution of 18 discrepant results comparing conventional pheno-/genotypic and MALDI-TOF MS GPR identifications
Identification type and no. of isolates
Conventional identificationa
MALDI-TOF MS
identificationb Resolution of discrepancy
On genus level with MALDI-TOF MS
1 Propionibacterium avidum Propionibacteriumsp. MALDI-TOF MS: low discrimination betweenP.avidum
andP.propionicum
On genus level with conventional methods
4 Actinobaculumsp. Actinobaculum schaalii Genus assignment by 16S rRNA gene analysis: sequence similarities toA.schaalii, 97.5%, 97.6%, 97.8%, and 98.5%, respectively
1 Corynebacteriumsp. Corynebacterium accolens Genus assignment by 16S rRNA gene analysis: sequence similarity toC.propinquum, 98.6%
3 Corynebacteriumsp. Corynebacterium aurimucosum 16S rRNA gene analysis: no discrimination ofC.
aurimucosumandC.minutissimum.rpoBgene sequence analysis confirmedC.aurimucosum(95.4%, 95.9%, and 96.9% sequence homology, respectively) 1 Corynebacteriumsp. Corynebacterium aurimucosum Genus assignment by 16S rRNA gene analysis: sequence
similarity toC.aurimucosum, 98.8%, and toC.
minutissimum, 98.5%
2 Corynebacteriumsp. Corynebacterium afermentans Genus assignment by 16S rRNA gene analysis: sequence similarity toCorynebacterium appendicis, 98.2% and 98.8%, respectively
1 Nocardiasp. Nocardia higoensis 16S rRNA gene analysis: sequence homology toN.
higoensisandNocardia shimofusensis, 100%, no differentiation
1 Propionibacteriumsp. Propionibacterium avidum Genus assignment by 16S rRNA gene analysis: sequence similarity toP.avidum, 96.7%
Misidentification
1 Corynebacteriumsp. “Corynebacterium lipophile” 16S rRNA gene analysis: sequence homology to “C. pseudogenitalium,” 100%
1 Corynebacterium
pseudodiphtheriticum
Corynebacterium propinquum Biochemical misidentification: 16S rRNA andrpoBgene analysis confirmedC.propinquum(100% and 98.1% sequence homology, respectively)
1 Corynebacterium
tuberculostearicum
“Corynebacterium lipophile” Biochemical misidentification: 16S rRNA gene sequence similarity to “C. pseudogenitalium,” 99.4%
1 Alloscardovia omnicolens Actinobaculum schaalii Mixed culture ofA.omnicolensandA.schaalii aConventional identification according to Bosshard et al.1. In addition, all discrepant isolates were analyzed by 16S RNA gene analysis. If not otherwise stated, identifications by
16S rRNA gene analysis and conventional identification were identical.
bMALDI-TOF MS identification using the direct transfer-formic acid protocol and a species cutoff of 1.7 with the Bruker database version 3.1.2.0.
on May 16, 2020 by guest
http://jcm.asm.org/
Reducing the cutoff value increased the number of low dis-criminations and misidentifications. However, this effect was lim-ited to certain species and reflects taxonomic inconsistencies rather than technical problems. The most frequent discrepancy was the misidentification of “C. pseudogenitalium” (per 16S rRNA gene analysis) as “C. lipophile.” “C. lipophile” is not an officially validated species, and the assignment of the single “C. lipophile” isolate in the Bruker database is unclear. Unfortunately, “C. pseu-dogenitalium” is not included in the Bruker database. It was de-scribed by Furness et al. (34) but has not yet been officially added to the list of recognized species, and would thus be better referred to as “CandidatusCorynebacterium pseudogenitalium.” As “C. lipophile” is not an officially accepted species name, we suggest that MALDI-TOF MS identifications of “C. lipophile” be ignored and that alternative identification methods be used for such iso-lates until additional entries have been added to the MALDI data-base and/or the taxonomy has been clarified. In conclusion, re-ducing the genus cutoff score value offers minimal gain of information but an increased number of misidentifications. In contrast, lower species cutoff values lead to significantly increased identification rates, thus, reducing the necessity for additional tests and increasing the efficiency of the laboratory workflow in terms of labor and costs. To avoid the problem of low discrimina-tion at the species level, we suggest using a two-step interpretadiscrimina-tion: In a first step, data interpretation should be done according to the standard criteria of the manufacturer (species cutoff value 2.0). In a second step, the Biotyper results list should be reinterpreted,
with the application of a lower species cutoff (preferably 1.7) for those isolates that did not yield species identification when using the standard species cutoff value of 2.0.
Coverage of the reference database.Analyses of some more rarely encountered genera and species were limited to the retro-spective part of the study. Several species are not included in the commercial Biotyper database, e.g.,A.johnsonii,C. pyruvicipro-ducens, “C. pseudogenitalium,” andP.campinasensis. Addition of those and other reference spectra from the extended in-house da-tabase that was generated in the retrospective study part increased the overall identification rate from 71.9% (commercial database) to 83.7% (commercial database amended with in-house data-base). As a consequence, the rate of nonidentifications was re-duced to 5.6%. Thus, the addition of single spectra of rarely iso-lated species significantly improved the performance of the Biotyper system. Proprietary in-house reference spectra were pro-vided to Bruker Daltonik for integrating selected data sets into the commercial database.
Identification of the clinically most common GPR isolates by MALDI-TOF MS in the prospective study yielded genus and spe-cies identifications of 87.4% and 79.1%, respectively, and was highly reliable. The identification rates were, however, lower than those reported for Gram-positive cocci (9,26,32). Apart from “C. lipophile,” no misidentifications occurred.
MALDI-TOF MS identification versus established identifica-tion algorithm.The current aerobic GPR identification algorithm in our clinical laboratory relies on a two-step procedure that
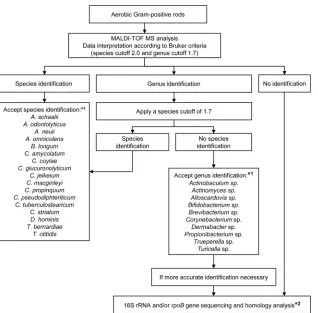
com-FIG 1Algorithm for the identification of Gram-positive rods in routine diagnostics using MALDI-TOF MS. Recommendations are based on the results of the prospective study. *1, Genera and species with⬍3 correctly identified isolates were not integrated in this algorithm. It is suggested that these isolates be identified by molecular analysis until sufficient data are available to update the approved lists; *2, 16S rRNA gene andrpoBgene sequencing as described previously (1,30,31).
on May 16, 2020 by guest
http://jcm.asm.org/
[image:7.585.137.455.63.376.2]bines phenotypic and molecular methods (1). In a first step, GPR isolates are identified by phenotypic and biochemical traits. For clinically relevant isolates, which are not identified to species level in this first step, 16S rRNA gene sequence analysis is done in a second step. Thirty-three percent of the isolates in the prospective study could not be identified to the species level by phenotypic methods and were subjected to 16S rRNA gene analysis. By MALDI-TOF MS, using the Bruker database and a species cutoff of 2.0, 33% of the isolates were also not identified to the species level (data not shown). Reduction of the species cutoff to 1.7 in-creased the species identification rate to 79.1%. The additional use of our extended in-house database further reduced the percentage of isolates not identified to the species level by MALDI-TOF MS to 16.3%. Thus, replacing phenotypic methods by MALDI-TOF MS has the potential to significantly reduce the amount of isolates which need to be sequenced for proper identification. We propose an adapted diagnostic algorithm for the clinical laboratory by combining MALDI-TOF MS and nucleic acid sequencing (Fig. 1). Isolates are first identified by MALDI-TOF MS, and 16S rRNA gene analysis is done only if no species identification is achieved. AdditionalrpoBgene sequencing is required for the identification ofCorynebacteriumspp. when the 16S rRNA target fails to dis-criminate closely related species. To optimize the species identifi-cation rate without increasing numbers of low discrimination, a species cutoff of 2.0 is first applied. For isolates that are not iden-tified to the species level, a species cutoff of 1.7 is subsequently used.
In conclusion, this study showed that the identification of GPR by the Bruker MALDI Biotyper is highly reliable, although iden-tification rates are generally lower than those for Gram-positive cocci or Gram-negative bacteria. Identification rates can be im-proved by (i) using direct transfer-formic acid sample prepara-tion, (ii) reducing the species cutoff score value, and (iii) by ex-panding the commercial database with in-house-generated reference spectra. Based on our data, we suggest a practical algo-rithm combining MALDI-TOF MS with nucleic acid sequence analysis for the identification of GPR in clinical laboratories. This algorithm is based on the current manufacturers system setup (i.e., score value cutoffs and consistency rules), but includes cutoff value variations if no species identification can be achieved by the standard rules. This algorithm covers the most frequently found genera and species and can easily be complemented by validated new database entries.
ACKNOWLEDGMENTS
We thank the laboratory technicians of the Institute of Medical Microbi-ology for their assistance and F. Mouttet for aid with statistical analyses.
The Institute of Medical Microbiology, University of Zurich, Switzer-land, has a collaboration agreement with Bruker Daltonik GmbH (Bre-men, Germany) for the purpose of improving the commercially available Bruker database. Bruker Daltonik had no influence on data collection or the interpretation of this study.
This study was supported by the University of Zurich.
REFERENCES
1.Bosshard PP, Abels S, Zbinden R, Böttger EC, Altwegg M.2003. Ribosomal DNA sequencing for identification of aerobic gram-positive rods in the clin-ical laboratory (an 18-month evaluation). J. Clin. Microbiol.41:4134 – 4140. http://dx.doi.org/10.1128/JCM.41.9.4134-4140.2003.
2.Neville SA, Lecordier A, Ziochos H, Chater MJ, Gosbell IB, Maley MW, van Hal SJ.2011. Utility of matrix-assisted laser desorption ionization– time of flight mass spectrometry following introduction for routine
labo-ratory bacterial identification. J. Clin. Microbiol.49:2980 –2984.http://dx .doi.org/10.1128/JCM.00431-11.
3.Tan EK, Ellis BC, Lee R, Stamper PD, Zhang SX, Carroll KC.2012. Prospective evaluation of a MALDI-TOF MS system in a hospital clinical microbiology laboratory for the identification of bacteria and yeasts: a bench-by-bench study to assess the impact on time-to-identification and cost-effectiveness. J. Clin. Microbiol.50:3301–3308.http://dx.doi.org/10 .1128/JCM.01405-12.
4.Demirev PA, Fenselau C.2008. Mass spectrometry for rapid character-ization of microorganisms. Annu. Rev. Anal. Chem.1:71–93.http://dx .doi.org/10.1146/annurev.anchem.1.031207.112838.
5.Croxatto A, Prod’hom G, Greub G.2012. Applications of MALDI-TOF mass spectrometry in clinical diagnostic microbiology. FEMS Microbiol. Rev.36:380 – 407.http://dx.doi.org/10.1111/j.1574-6976.2011.00298.x. 6.Wieser A, Schneider L, Jung J, Schubert S.2012. MALDI-TOF MS in
microbiological diagnostics-identification of microorganisms and be-yond (mini review). Appl. Microbiol. Biotechnol.93:965–974.http://dx .doi.org/10.1007/s00253-011-3783-4.
7.Carbonnelle E, Grohs P, Jacquier H, Day N, Tenza S, Dewailly A, Vissouarn O, Rottman M, Herrmann JL, Podglajen I, Raskine L.2012. Robustness of two MALDI-TOF mass spectrometry systems for bacterial identification. J. Microbiol. Methods 89:133–136. http://dx.doi.org/10 .1016/j.mimet.2012.03.003.
8.Bille E, Dauphin B, Leto J, Bougnoux ME, Beretti JL, Lotz A, Suarez S, Meyer J, Join-Lambert O, Descamps P, Grall N, Mory F, Dubreuil L, Berche P, Nassif X, Feroni A.2012. MALDI-TOF MS Andromas strategy for the routine identification of bacteria, mycobacteria, yeasts,Aspergillus
spp. and positive blood cultures. Clin. Microbiol. Infect.18:1117–1125. http://dx.doi.org/10.1111/j.1469-0691.2011.03688.x.
9.Dubois D, Grare M, Prere MF, Segonds C, Marty N, Oswald E.2012. Performances of the Vitek MS matrix-assisted laser desorption ioniza-tion–time of flight mass spectrometry system for rapid identification of bacteria in routine clinical microbiology. J. Clin. Microbiol.50:2568 – 2576.http://dx.doi.org/10.1128/JCM.00343-12.22593596.
10. Martiny D, Busson L, Wybo I, El Haj RA, Dediste A, Vandenberg O.
2012. Comparison of the Microflex LT and Vitek MS systems for routine identification of bacteria by matrix-assisted laser desorption ionization– time of flight mass spectrometry. J. Clin. Microbiol.50:1313–1325.http: //dx.doi.org/10.1128/JCM.05971-11.
11. van Veen SQ, Claas EC, Kuijper EJ.2010. High-throughput identifica-tion of bacteria and yeast by matrix-assisted laser desorpidentifica-tion ionizaidentifica-tion– time of flight mass spectrometry in conventional medical microbiology laboratories. J. Clin. Microbiol. 48:900 –907.http://dx.doi.org/10.1128 /JCM.02071-09.
12. Bizzini A, Durussel C, Bille J, Greub G, Prod’hom G.2010. Performance of matrix-assisted laser desorption ionization–time of flight mass spec-trometry for identification of bacterial strains routinely isolated in a clin-ical microbiology laboratory. J. Clin. Microbiol.48:1549 –1554.http://dx .doi.org/10.1128/JCM.01794-09.
13. Alatoom AA, Cazanave CJ, Cunningham SA, Ihde SM, Patel R.2012. Identification of non-diphtheriae Corynebacterium by use of matrix-assisted laser desorption ionization–time of flight mass spectrometry. J. Clin. Microbiol.50:160 –163.http://dx.doi.org/10.1128/JCM.05889-11. 14. Vila J, Juiz P, Salas C, Almela M, de la Fuente CG, Zboromyrska Y,
Navas J, Bosch J, Aguero J, de la Bellacasa JP, Martínez-Martínez L.
2012. Identification of clinically relevantCorynebacterium spp., Arcano-bacterium haemolyticum, andRhodococcus equiby matrix-assisted laser desorption ionization–time of flight mass spectrometry. J. Clin. Micro-biol.50:1745–1747.http://dx.doi.org/10.1128/JCM.05821-11.
15. Konrad R, Berger A, Huber I, Boschert V, Hormansdorfer S, Busch U, Hogardt M, Schubert S, Sing A.2010. Matrix-assisted laser desorption/ ionisation time-of-flight (MALDI-TOF) mass spectrometry as a tool for rapid diagnosis of potentially toxigenicCorynebacteriumspecies in the laboratory management of diphtheria-associated bacteria. Euro Surveill.
15:pii 19699.
16. Ng LS, Sim JH, Eng LC, Menon S, Tan TY. 2012. Comparison of phenotypic methods and matrix-assisted laser desorption ionisation time-of-flight mass spectrometry for the identification of aero-tolerant Actino-mycesspp. isolated from soft-tissue infections. Eur. J. Clin. Microbiol. Infect. Dis.31:1749 –1752.http://dx.doi.org/10.1007/s10096-011-1496-3. 17. Verroken A, Janssens M, Berhin C, Bogaerts P, Huang TD, Wauters G, Glupczynski Y.2010. Evaluation of matrix-assisted laser desorption ion-ization–time of flight mass spectrometry for identification ofNocardia
on May 16, 2020 by guest
http://jcm.asm.org/
species. J. Clin. Microbiol.48:4015– 4021.http://dx.doi.org/10.1128/JCM .01234-10.
18. Barbuddhe SB, Maier T, Schwarz G, Kostrzewa M, Hof H, Domann E, Chakraborty T, Hain T.2008. Rapid identification and typing ofListeria
species by matrix-assisted laser desorption ionization–time of flight mass spectrometry. Appl. Environ. Microbiol.74:5402–5407.http://dx.doi.org /10.1128/AEM.02689-07.
19. Schmitt BH, Cunningham SA, Dailey AL, Gustafson DR, Patel R.2013. Identification of anaerobic bacteria by Bruker Biotyper matrix-assisted laser desorption ionization–time of flight mass spectrometry with on-plate formic acid preparation. J. Clin. Microbiol.51:782–786.http://dx .doi.org/10.1128/JCM.02420-12.
20. Lau SK, Tang BS, Teng JL, Chan TM, Curreem SO, Fan RY, Ng RH, Chan JF, Yuen KY, Woo PC.18 October 2013. Matrix-assisted laser desorption ionisation time-of-flight mass spectrometry for identification of clinically significant bacteria that are difficult to identify in clinical laboratories. J. Clin. Pathol. http://dx.doi.org/10.1136/jclinpath-2013 -201818.
21. Bizzini A, Jaton K, Romo D, Bille J, Prod’hom G, Greub G. 2011. Matrix-assisted laser desorption ionization–time of flight mass spectrom-etry as an alternative to 16S rRNA gene sequencing for identification of difficult-to-identify bacterial strains. J. Clin. Microbiol.49:693– 696.http: //dx.doi.org/10.1128/JCM.01463-10.
22. Freiwald A, Sauer S.2009. Phylogenetic classification and identification of bacteria by mass spectrometry. Nat. Protoc.4:732–742.http://dx.doi .org/10.1038/nprot.2009.37.
23. Haigh J, Degun A, Eydmann M, Millar M, Wilks M.2011. Improved performance of bacterium and yeast identification by a commercial ma-trix-assisted laser desorption ionization–time of flight mass spectrometry system in the clinical microbiology laboratory. J. Clin. Microbiol.49:3441. http://dx.doi.org/10.1128/JCM.00576-11.
24. Theel ES, Schmitt BH, Hall L, Cunningham SA, Walchak RC, Patel R, Wengenack NL.2012. Formic acid-based direct, on-plate testing of yeast andCorynebacteriumspecies by Bruker Biotyper matrix-assisted laser de-sorption ionization–time of flight mass spectrometry. J. Clin. Microbiol.
50:3093–3095.http://dx.doi.org/10.1128/JCM.01045-12.
25. Farfour E, Leto J, Barritault M, Barberis C, Meyer J, Dauphin B, Le Guern AS, Leflèche A, Badell E, Guiso N, Leclercq A, Le Monnier A, Lecuit M, Rodriguez-Nava V, Bergeron E, Raymond J, Vimont S, Bille
E, Carbonnelle E, Guet-Revillet H, Lécuyer H, Beretti JL, Vay C, Berche P, Ferroni A, Nassif X, Join-Lambert O.2012. Evaluation of the Andro-mas matrix-assisted laser desorption ionization–time of flight Andro-mass spec-trometry system for identification of aerobically growing Gram-positive bacilli. J. Clin. Microbiol.50:2702–2707.http://dx.doi.org/10.1128/JCM .00368-12.
26. McElvania Tekippe E, Shuey S, Winkler DW, Butler MA, Burnham CA.
2013. Optimizing identification of clinically relevant Gram-positive or-ganisms by use of the Bruker Biotyper matrix-assisted laser desorption ionization–time of flight mass spectrometry system. J. Clin. Microbiol.
51:1421–1427.http://dx.doi.org/10.1128/JCM.02680-12.
27. Goodfellow M, Parte A, Kämpfer P, Busse H-J, Trujillo ME, Suzuki K-i, Ludwig W, Whitman WB (ed).2012. Bergey’s manual of systematic bacteriology, vol 5A. TheActinobacteria. Springer, New York, NY. 28. Von Graevenitz A, Funke G.1996. An identification scheme for rapidly
and aerobically growing gram-positive rods. Zentralbl. Bakteriol.284:
246 –254.
29. Clinical and Laboratory Standards Institute.2008. Interpretative criteria for identification of bacteria and fungi by DNA target sequencing; Ap-proved Guideline. CLSI document MM18-A. Clinical and Laboratory Standards Institute, Wayne, PA.
30. Khamis A, Raoult D, La Scola B.2004.rpoBgene sequencing for iden-tification ofCorynebacteriumspecies. J. Clin. Microbiol.42:3925–3931. http://dx.doi.org/10.1128/JCM.42.9.3925-3931.2004.
31. Khamis A, Raoult D, La Scola B.2005. Comparison betweenrpoBand 16S rRNA gene sequencing for molecular identification of 168 clinical isolates ofCorynebacterium. J. Clin. Microbiol.43:1934 –1936.http://dx .doi.org/10.1128/JCM.43.4.1934-1936.2005.
32. Schulthess B, Brodner K, Bloemberg GV, Zbinden R, Böttger EC, Hombach M.2013. Identification of Gram-positive cocci by use of ma-trix-assisted laser desorption-time of flight mass spectrometry: compari-son of different preparation methods and implementation of a practical algorithm for routine diagnostics. J. Clin. Microbiol.51:1834 –1840.http: //dx.doi.org/10.1128/JCM.02654-12.
33. Viera AJ, Garrett JM.2005. Understanding interobserver agreement: the kappa statistic. Fam. Med.37:360 –363.
34. Furness G, Sambury S, Evangelist AT.1979.Corynebacterium pseudo-genitaliumsp. nov. Commensals of the human male and female urogenital tracts. Invest. Urol.16:192–295.