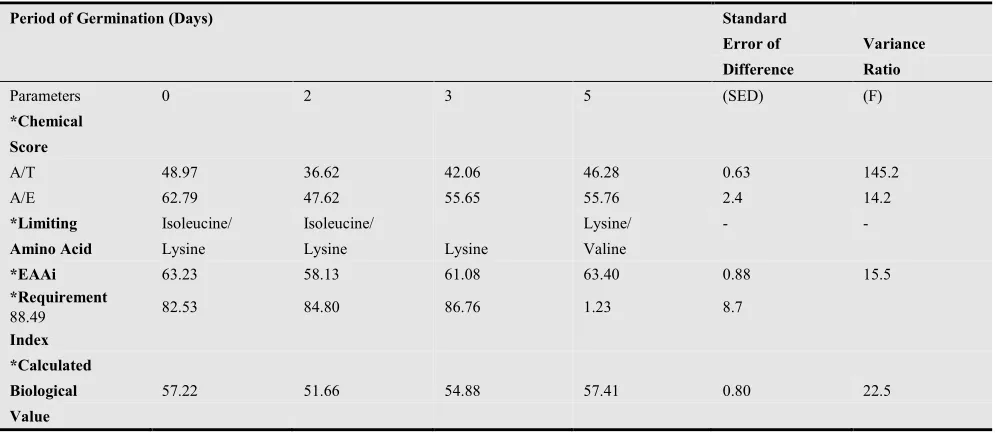
Chemical Evaluation of Protein Quality of Sprouted Maize and Cowpea
Full text
Figure
Related documents
Methods: The development on intensive animal production, non- pollution foods, and marketing mode of animal drugs of animal pharmaceutical industry, and the standardized
In studies performed on animals, increased expression of HSP70 proteins and HSP40 proteins was detected in the colorectal mucosa of mice with pharmacologically induced
In conclusion, findings from this study suggest that COPD/CB patients with a history of severe exacerba- tions as well as a high use of COPD medication experi- enced a
and no signifi- cant difference of overall survival probability was observed in the patients with organ-confined bladder cancer ( p = 0.675) and in patients with no
Twenty-five percent of our respondents listed unilateral hearing loss as an indication for BAHA im- plantation, and only 17% routinely offered this treatment to children with
Despite critics, I argue that advance directives are especially advisable in dementia and provide reasons in favor of their promotion at an early stage of the disease as a valuable
