The human T transcription factor:
a study of genetics and function
Charalambos Papapetrou
December 1998
Thesis submitted for the degree of Doctor of Philosophy
in the University of London
MRC Human Biochemical Genetics Unit
The Galton Laboratory
Department of Biology
University College London
ProQuest Number: 10010154
All rights reserved
INFORMATION TO ALL USERS
The quality of this reproduction is dependent upon the quality of the copy submitted. In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uest.
ProQuest 10010154
Published by ProQuest LLC(2016). Copyright of the Dissertation is held by the Author. All rights reserved.
This work is protected against unauthorized copying under Title 17, United States Code. Microform Edition © ProQuest LLC.
ProQuest LLC
789 East Eisenhower Parkway P.O. Box 1346
A b str a c t
The human T gene is a member of a family of transcription factors, the T-box genes, which play an important role during embryogenesis. T is expressed early in development in the primitive streak, axial mesoderm, notochord and tail bud and is
essential for normal mesoderm development and notochord differentiation. Mouse T
mutations are lethal and heterozygotes have features that resemble neural tube defects
and sacral agenesis in man.
This thesis describes the genetic analysis of the human T gene in healthy individuals and in neural tube defect and sacral agenesis patients. Eight new
polymorphisms were identified by a combination of SSCP and sequence analyses and
three of these involve an amino acid change in evolutionary conserved domains;
Glyl77Asp in the DNA binding domain and Gly356Ser and Asn369Ser in a
transcriptional activation domain. An association between an allele {TlVSy-l) and a particular haplotype (T363.G530.IVS7-2) and susceptibility to spina bifida was detected. Analysis of T in sacral agenesis patients identified a rare variant Ala338Val in the transcriptional activation domain, in a single patient and his mother. Overall it was
concluded that T may play a role in the aetiology of neural tube defects and sacral agenesis, but accounts for only a small proportion of the genetic component of
susceptibility to these disorders.
The proximal promoter of the human T gene was cloned, sequenced and compared with the mouse sequence in a search for conserved regulatory elements.
Several potential motifs were identified, the most significant of which was a 30 bp
region conserved not only between man and mouse, but also Xenopus.
In vitro DNA/protein binding studies were carried out to demonstrate that human T protein is able to bind to a DNA motif known to be the target for mouse T protein. In
addition, it was demonstrated that both human and mouse T bind their DNA target as a
dimer and could form a heterodimer. It was shown that the Asp 177 variant of the DNA
binding domain reduces the stability of T dimer formation. Mouse Tbx6 protein, another member of the T-box family, was shown to bind to the same DNA target, but it was
demonstrated that T and Tbx6 are not likely to bind this target as a heterodimer.
The human homologue of the TBX6 gene was cloned, sequenced and mapped to chromosome 16pl 1.2. Expression studies showed that TBX6 is expressed in the
notochord and tail bud in the early stages of development, but also in a wide range of
adult tissues. A novel member of the T-box gene family, designated T-like, was identified by sequence analysis of PGR products. Preliminary analysis suggests that
Statement from Charalambos Papapetrou regarding contribution to work in each Results Chapter
C h a p te r 3 T he g e n o ty p e analysis o f the TIVS^ p o ly m o rp h ism in N T D fa m ilies and controls was divided 50:50 with M s Katie M orrison. All o f the data analysis was carried out by me.
T h e pape r M o rriso n et al. 1996 was largely w ritten by P ro fe sso r Y v o n n e H. E dw a rds, how ever, I con trib u ted sections in the M aterials and M e th o d s section and the section describing the association analysis.
I was entirely responsible for the finding o f p o ly m o rp h ism s in T e x o n s 2 and 3 and for the g en o ty p e analysis in N T D fam ilies and controls and for an a ly sin g the data. I was also responsible for the S S C P analysis o f T exon 1.
T h e g en o ty p in g o f the M T H F R therm olabile variant in the British N T D fam ilies and controls, the statistical analysis and the association studies with the T variants was also m y work.
T h e Papapetrou et al., 1996 p ap e r was w ritten by m e in the first instance and revised by P rofessor Y v o n n e H. Edw ards.
C h a p te r 4 F or 15 o f the 31 sacral agenesis cases I carried out all o f the g e n o ty p e analysis, this included S S C P and direct sequencing. For the re m a in in g 16, w hich w ere only sequenced, the w ork w as divided 50:50 with D r Felicity D ru m m o n d .
1 was entirely responsible for the search for p o ly m o rp h ism s in T e x o n s 4, 5, 6, 7, and 8 and the 5 ’U T R and also for the characterisation o f these p o ly m o rp h is m s by s eq u e n cin g and their analysis in controls. I also investigated the o c c u rre n c e o f the rare variant in exon 7 in N T D patients and controls by genotyping.
T he lam b d a clone co n tain in g the hu m an T p ro m o te r w as isolated and D N A pre pare d by Ms W e n d y Putt. M s Putt also carried out the S o u th e rn analysis and su b clo n ed the pro m o ter contain in g fragm ents. I was responsible for the seq u e n c in g o f the p ro m o te r and the s u b seq u e n t seque nce c o m p ariso n s and search for transcription factor binding sites.
C h a p te r 5 All o f the w ork d escribed in this C h ap ter w as carried out in person, i.e. preparation o f the in vitro synthesised protein, all F M S A s , cro sslin k in g e x p e rim e n ts etc.
T h e P apapetrou et al., 1997 p ap e r was written by m e with critical reading and revision by P rofessor Y v o n n e FI. E dw ards.
C h a p te r 6 I was responsible for the library screening to identify T B X 6 and fo r the isolation o f the clone and p reparation o f D N A . M s W e n d y Putt su b c lo n e d exon c o n ta in in g fragm ents and these w ere seq u e n ced as a jo in t effort. F IS H analysis was carried out by Dr M arg aret Fox using the D N A I had prepared. All o f the su b se q u e n t T BX6 analysis was carried out in person. All o f the R T - P C R e xpression studies w e re my ow n work.
I was responsible for the identification o f the T-like g ene and its su b seq u e n t characterisation by partial seque nce analysis and R T -P C R .
Acknowledgements
M y first thanks go to Professor Yvonne Edwards, who supervised this
project and to whom I am m ost grateful for her invaluable help and endless
support, encouragem ent and enthusiasm throughout this work. Special thanks
also go to D r Jane Sowden for her support, useful discussions and help,
especially with the functional study. I would also like to thank everyone in the
laboratory for their support and help and for making a happy w orking
environm ent. I particularly thank W endy Putt for all her technical advice and
help with the cloning o f the hum an T prom oter and TBX6 gene and Katie M orrison for her help with the TIVSy polym orphism analysis. M y special thanks go to Dr Felicity Drum mond for her special friendship and support, as
well as technical advice, useful discussions and help with the sacral agenesis
work. Thanks also to James W ilson and Ira Islam for their valuable help. I
would also like to thank D r M argaret Fox for providing the FISH analysis. Dr
W illiam Reardon and Professor Robin W inter for providing the sacral agenesis
samples and our colleagues in Newcastle and The Netherlands for providing
the neural tube defects families. On a personal level, I would like to thank my
parents for all their love, encouragem ent and support throughout m y studies
and my girlfriend M arina for her love, support, patience and constructive
reading o f this manuscript.
Contents
Abstract 2
Acknowledgements 3
Contents 4
Figures 7
Tables 10
Abbreviations 12
CHAPTER ONE
Introduction 16
1.1 The r gene 17
1.1.1 Structure of the T gene 17
1.1.2 r expression 18
1.1.3 Regulation of T expression 22
1.1.4 T as a transcription factor 26
1.1.5 The function of T and its role in embryogenesis 28
1.1.6 The T-box transcription factors 3 3
1.2 Neural tube defects 36
1.2.1 Normal human neurulation 37
1.2.2 Clinical features of NTD 39
1.2.3 Genetics of NTD 42
1.2.4 Candidate genes in abnormal neural tube development 44
1.2.5 Environmental factors and NTD 46
1.2.6 The folic acid metabolism pathway 47
1.2.7 T and neural tube defects 50
1.3 Sacral agenesis 52
1.3.1 Classification and associated disorders 52
1.3.2 Genetic aetiology of caudal dysgenesis 56
1.3.3 T and sacral agenesis 58
1.4 Research aims 60
CHAPTER TWO
Materials and Methods 61
2.1 Materials 61
2.1.1 Standard reagents 61
2.1.2 Enzymes 61
2.1.3 Electrophoresis reagents 61
2.1.4 Miscellaneous 61
2.1.5 Commonly used solutions and buffers 62
2.1.7 DNA samples 63
2.2 Methods 64
2.2.1 PCR amplification of genomic DNA 65
2.2.2 Agarose gel electrophoresis and recovery of DNA fragments 67
2.2.3 DNA precipitation 67
2.2.4 Single strand conformation polymorphism (SSCP) analysis 67
2.2.5 DNA modification 70
2.2.6 DNA sequencing 70
2.2.7 Isolation of RNA 71
2.2.8 Preparation of cDNA by reverse transcription and RT-PCR 72
2.2.9 In vitro transcription/translation 73
2.2.10SDS-PAGE 75
2.2.11 Quantification of in vitro transcription/translation products by TCA
precipitation 75
2.2.12 Preparation of ^^P-labelled probes for EMSAs 75
2.2.13 Electrophoretic mobility shift assays (EMSAs) 76
2.2.14 Preparation of ^^P-labelled probes for library screening 77
2.2.15 h2a genomic library screening 78
2.2.16 À2001 genomic library screening 79
2.2.17 Fluorescent in situ hybridisation (FISH) 81
2.2.18 Association analysis 82
CHAPTER THREE
Investigation of the human T gene as a candidate gene for susceptibility to
neural tube defects 85
3.1 A common polymorphism in intron 7 of the human T gene 85
3.2 Human T and NTD: an association analysis 87
3.2.1 The transmission disequilibrium test (TDT) 88
3.2.2 Case/control studies 90
3.3 Detection of polymorphisms by SSCP 92
3.4 Analysis of exons 1-3 of the DNA binding domain of T 96
3.4.1 A Serl21Ser (C363T) polymorphism in Texon 2 98
3.4.2 A Glyl77Asp (G530A) polymorphism in Texon 3 102
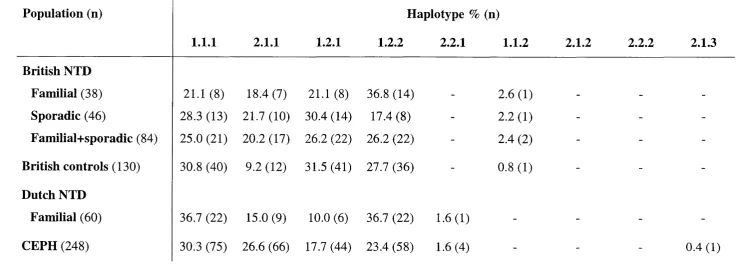
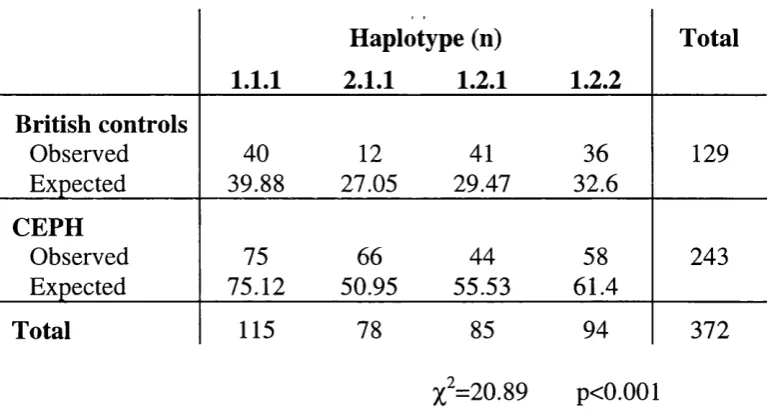
3.5 Haplotype analysis 106
3.6 Methylene tetrahydrofolate reductase and NTD 115
3.7 Discussion 121
3.7.1 T and susceptibility to NTD 121
CHAPTER FOUR
Genetic analysis of the human T gene in sacral agenesis 126
4.1 SSCP analysis 127
4.2 Automated fluorescent sequence analysis 134
4.3 A novel mutation in a sacral agenesis patient 137
4.4 Cloning of the human T promoter 144
4.5 Discussion 148
4.5.1 A comparison of SSCP and sequence analysis 148
4.5.2 Genetic variation in the human T gene 149
4.5.3 T and sacral agenesis 154
4.5.4 Human Tpromoter 154
CHAPTER FIVE
An investigation of the DNA binding properties of the human T protein 157
5.1 T protein synthesis 157
5.2 A comparison of the DNA binding activity of the truncated Glyl77 and
Asp 177 T proteins 162
5.3 T protein binds as a dimer 169
5.4 An investigation of the possible interaction between T and Tbx6 173
5.4.1 Tbx6 protein synthesis 174
5.4.2 Tbx6 binding properties 176
5.5 Discussion 178
5.5.1 DNA binding properties of T 178
5.5.2 DNA binding properties of Tbx6 183
CHAPTER 6
Cloning of other T-hox genes 187
6.1 Cloning of human TBX6 187
6.2 Identification of a human T-like gene 200
6.3 Discussion 203
6.3.1 Human TBX6 203
6.3.2 Human TBXT 205
CHAPTER SEVEN
Final discussion 208
Appendix I Primers used in the amplification of the human T gene 216 Appendix II Pileup of the deduced aa sequences of human T homologues 218
Appendix HI Sacral agenesis pedigrees 220
Appendix IV 5'-flanking sequence of the human T gene 223
F ig u res
Page
CHAPTER ONE
Introduction
Figure 1.1 Schematic representation of the T gene and its protein 19
Figure 1.2 The formation of the neural tube 38
Figure 1.3 Schematic representation of the neural tube closure 40
Figure 1.4 Intracellular pathways of the folic acid metabolism 48
CHAPTER THREE
Investigation of the human T gene as a candidate gene for susceptibility to
neural tube defects
Figure 3.1 Analysis of the human T intron 7 TIVSjC polymorphism 86
Figure 3.2 SSCP of the human T intron 7 TIVSjC polymorphism 95
Figure 3.3 SSCP analysis of the human T exon 1 coding sequence 97
Figure 3.4 Analysis of the human T exon 2 C363T polymorphism 99
Figure 3.5 Analysis of the human T exon 3 G530A (Glyl77Asp)
polymorphism 103
Figure 3.6 Diagnostic digest of mouse T exon 3 104
Figure 3.7 Cladogram illustrating possible ancestral Tchromosomes 113 Figure 3.8 Digestion analysis of the human MTHFR C677T
polymorphism 116
CHAPTER FOUR
Genetic analysis of the human T gene in sacral agenesis
Figure 4.1 SSCP analysis of T exon 6 129
Figure 4.2 SSCP analysis of T exon 4 (A) and exon 5 (B) 130
Figure 4.3 SSCP analysis of T exon 7 131
Figure 4.4 SSCP analysis of T exon 8 133
Figure 4.5 Analysis of the human T 5'UTR variant 135
Figure 4.6 Analysis of the human T exon8 G1176A variant 136
Figure 4.7 Analysis of the human T exon 8G1066A variant 138
Figure 4.9 Analysis of the human T intron 7 AIVS7G variant 140 Figure 4.10 Analysis of the human T exon 7 C1013T (Ala338Val)
mutation 143
Figure 4.11 Restriction enzyme map of the 5' end of the human T gene 145 Figure 4.12 Comparison of the human and mouse 5'-flanking
sequences 146
Figure 4.13 Analysis of T exon 3 of a sacral agenesis patient 150
Figure 4.14 The human T gene and protein showing various
polymorphisms 151
CHAPTER FIVE
An investigation of the DNA binding properties of the human T protein
Figure 5.1 Summary of in vitro synthesis of truncated human T
protein 158
Figure 5.2 SDS-PAGE of in vitro synthesised T proteins 161
Figure 5.3 EMSAs using the Glyl77 and Asp 177 truncated human
T protein 163
Figure 5.4 Thermostability assay of truncated human T/BS.p
complexes 164
Figure 5.5 EMSAs using variable amounts of truncated human T
protein 166
Figure 5.6 Comparison of the DNA binding properties of Glyl77 and
Asp 177 T proteins 167
Figure 5.7 Comparison of the DNA-binding properties of Glyl77 and
Asp 177 T proteins 168
Figure 5.8 A. Formation of a heterodimeric T/BS.p complex between
full-length and truncated T proteins and B. thermostability
assay of homodimeric and heterodimeric T/BS.p complexes 170
Figure 5.9 A. EMSAs and B. SDS-PAGE of cross-linked truncated
human T protein 172
Figure 5.10 SDS-PAGE of in vitro synthesised truncated mouse Tbx6
protein (aa 1-288 and 46-288) 175
Figure 5.13 The crystallographic structure of the T-domain with its
DNA target 181
Figure 5.14 Sequence of the N-terminal aa of Xenopus, mouse and
human T homologues and mouse Tbx6 185
CHAPTER SIX
Cloning of other T-box genes
Figure 6.1 Comparison of the deduced aa sequences of human and
mouse TBX6 proteins 188
Figure 6.2 The human TBX6 cDNA and 5'-flanking sequences 190
Figure 6.3 Sequence analysis of human TBX6 to demonstrate the
presence of two extra nucleotides compared to mouse Tbx6 193 Figure 6.4 Comparison of the deduced aa sequences of the human and
adjusted mouse TBX6 proteins 194
Figure 6.5 FISH on normal metaphase chromosomes 195
Figure 6.6 RT-PCR using RNA from human and mouse tissues and
human TBX6 specific primers and diagnostic digests 197
Figure 6.7 RT-PCR using human tissue RNA and human T specific
primers and diagnostic digests 199
Figure 6.8 RT-PCR using human tissue RNA and human T specific
primers within the T-box region and diagnostic digests 201
Figure 6.9 A. Comparison of the fetal testis RT-PCR product and human
T and B. comparison of the deduced aa sequences of the
T a b le s
CHAPTER ONE
Introduction
Table 1.1 Main areas of expression of murine T-box genes
Page
35
CHAPTER TWO
Materials and methods
Table 2.1 Primers and annealing temperatures for the amplification of
the human T and MTHFR genes
Table 2.2 Primers and annealing temperatures for the amplification of
TandraXdcDNA
66
73
CHAPTER THREE
Investigation of the human Tgene as a candidate gene for
susceptibility to neural tube defects
Table 3.1 TDT using the TlVSy polymorphism 89
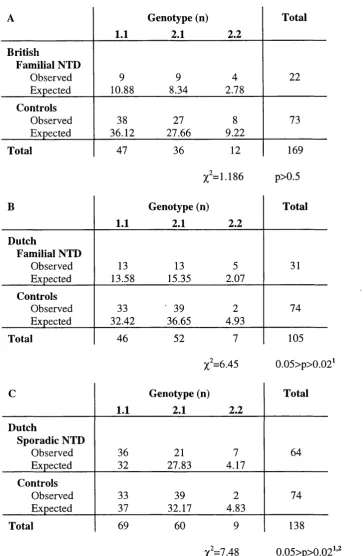
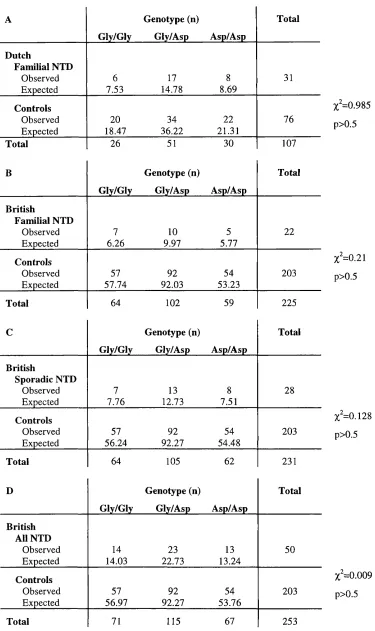
Table 3.2 Genotype and allele frequencies of the human TIVSj
polymorphism 91
Table 3.3 Contingency tables comparing TIVSj genotype distribution 93
Table 3.4 Conditions for SSCP analysis of the human T gene 96
Table 3.5 Genotype and allele frequencies of the human C363T
polymorphism 100
Table 3.6 TDT using the C363T (Serl21Ser) polymorphism 101
Table 3.7 Genotype and allele frequencies of the human Glyl77Asp
polymorphism 105
Table 3.8 TDT using the Glyl77Asp (G530A) polymorphism 107
Table 3.9 Contingency tables comparing Glyl77Asp genotype
distribution 108
Table 3.10 Haplotype frequencies 110
Table 3.11 Contingency tables comparing haplotype distribution 111
Table 3.12 Haplotype TDT 114
Table 3.13 Genotype and allele frequencies of the human MTHFR C677T polymorphism
Table 3.14 TDT using the MTHFR C677T polymorphism
Table 3.15 Association analysis of the T TIVSyC and MTHFR C677T
alleles
117
119
120
CHAPTER FOUR
Genetic analysis of the human Tgene in sacral agenesis
Table 4.1 Conditions for SSCP analysis of the human T gene Table 4.2 Sequence variation identified in the human T gene
128
A b b r e v ia tio n s
aa amino acid
Ala alanine
AMPS am m onium persulphate
Arg arginine
Asn asparagine
Asp aspartic acid
bp base pair
BS3 B is(sulfosuccinim idyl)suberate
BSA bovine serum albumin
BS.p palindrom ic DNA target of T protein
C- carboxy
CBS cystathionine-p-synthase
cDNA com plem entary deoxyribonucleic acid
CEPH Centre d ’Etude de Polym orphism e Hum aine
C l confidence interval
Ci Curie
DNA deoxyribonucleic acid
cpm counts per minute
ddNTP dideoxynucleoside triphosphate
DAP difference in allele frequencies
DEPC diethylpyrocarbonate
dNTP deoxynucleoside triphosphate
dpc days post coitum
DTT dithiothreitol
ED TA ethylenediam inetetra-acetic acid
EM SA electrophoretic m obility shift assay
epd eukaryotic prom oter database
FETi Fisher’s exact test, one-sided
FISH fluorescent in situ hybridisation
F R -a GCF Gin Gly GMS HAc HGM P HLA HNF-5 HPE IBD lie IVS kb L M APF2 M AZ M CK M HC M OPS m RNA MS M THFD
M TH FR
myc N-NTD P PAGE PBS PCR
folate receptor alpha
GC-binding factor
glutam ine
glycine
genome m ism atch scanning
acetic acid
hum an genom e mapping project
hum an leukocyte antigen
hepatocyte nuclear factor 5
holoprosencephaly
identical by descent
isoleucine
intervening sequence
kilo base
fast migrating, low band in EM SAs
m uscle actin prom oter factor 2
m yc-associated zinc finger protein
m otif from the m uscle creatine kinase prom oter
m ajor histocom patibility com plex
3-[A-m orpholino]propanesulfonic acid
messenger RNA
methionine synthase
methylene tetrahydrofolate dehydrogenase
methylene tetrahydrofolate reductase
m yelocytom atosis
amino
neural tube defect
probability
polyacrylam ide gel electrophoresis
phosphate-buffered saline
pfu PGM PM SF PN K PPi psi PSM R RN A RN Ase rpm RT-PCR E SA SAP SB SEA SBO SDS sdw SE
SEdaf
Ser SHH SSC SSCP SSPE TA TB E TC A TD Tplaque form ing units
phosphoglucom utase
phenylm ethylsulphonylfluoride
polynucleotide kinase
saturated tetrasodium pyrophosphate
pounds per square inch
phage storage m edium
repression
ribonucleic acid
ribonuclease
revolutions per m inute
reverse transcriptase polym erase chain reaction
sum
sacral agenesis
shrimp alkaline phosphatase
spina bifida
spina bifida aperta
spina bifida occulta
sodium dodecyl sulphate
sterile distilled w ater
standard error
standard error of difference in allele frequencies
serine
sonic hedgehog
salt sodium citrate
single strand conform ation polym orphism
salt sodium phosphate EDTA
transcriptional activation
Tris-borate/ED TA
trichloroacetic acid
transm ission disequilibrium test
TE Tris-ED TA
TEM ED N ,N ,N ',N '-tetram ethylethylenedi amine
TED transcription factors data base
Thr threonine
Tris Tris (hydroxym ethyl) am ino-m ethane
TRP-1 tyrosinase-related protein 1
UTR untranslated region
U slow migrating, upper band in EM SAs
uv ultraviolet
Val valine
VPA valproic acid
v/v volum e per volume
CHAPTER ONE
I n tr o d u c tio n
T is a transcription factor that plays a vital role in norm al axial
developm ent and form ation and differentiation of posterior m esoderm .
The m ouse T gene was identified in 1927 as a loss o f function mutation,
Brachyury (D obrovolskaia-Zavadskaia and Kobozieff, 1927). Hom ozygous em bryos die at m id-gestation due to a defective allantois w ith severe posterior
abnorm alities and an undifferentiated notochord. H eterozygotes show
incom plete axial developm ent and have posterior abnorm alities and short tails.
The m ouse T gene is part of the r-complex, a region o f the proxim al half o f m ouse chrom osom e 17, which for m any years intrigued developm ental
biologists. A naturally occurring variant o f the r-complex, called the t-
haplotype, is defined by four non-overlapping inversions and characterised by
distortion of m ale transm ission ratio, em bryonic lethality and m ale sterility
(Silver, 1993). Originally, t-haplotypes w ere referred to as r-mutations or
t-alleles and w ere thought to be m utant t-alleles o f the T locus. H owever,
t-haplotypes are invisible them selves, because neither heterozygous +/t nor live- born hom ozygous t/t m ice show abnorm alities. Interestingly, T m utations expose the f-haplotype. M ice carrying a m utant T allele on one chrom osom e in com bination with a w ild type r-complex on the other chrom osom e have
short tails, whereas, animals carrying a m utant T allele in com bination with a r-haplotype are tail-less.
The nature o f the T-locus and the Brachyury allele rem ained obscure for over 60 years until in 1990, H errm ann et al. (1990) cloned the m ouse T
gene. The T gene was revealed to be a nuclear localised protein with a unique D N A binding dom ain, designated the T-box. The Brachyury allele (7^"“^^) was show n to be a 200 kb deletion encom passing the entire T gene. Since then, the hom ologues from various species as diverse| as sea urchin (Harada et a i , 1995) and m an (Edwards et al., 1996) have been cloned.
W ork carried out in the last 5 years has shown that T is the founder m em ber o f a large fam ily o f T-box transcription factors. To date as m any as
13 m em bers have been cloned in m ouse and for most of these, orthologues
have been identified in man (see section 1.1.6). These genes all appear to be
critical to norm al developm ent. M utations in two o f the hum an T-box genes
(TBX3 and TBX5) have been associated with developm ental disorders. Thus far, no m utations of the Tgene have been identified in man and no clinical condition has been associated with abnorm al Texpression. However, the expression pattern o f T and the phenotype of Brachyury and other mouse m utants m ake the hum an T gene a good candidate for disorders that affect posterior axial developm ent. Examples of disorders w hich can be considered
in this category are neural tube defects, sacral agenesis and chordomas.
In subsequent sections of this chapter, the structure, expression,
regulation, function and role in em bryogenesis o f the Tgene will be described. Tw o disorders o f posterior axial developm ent, neural tube defects and sacral
agenesis, will be looked at in detail. Their aetiology, phenotype, genetics and
the possible role of T in their pathogenesis will be described.
1.1 TH E r G ENE
1.1.1 Structure of the T gene
The T gene has been cloned in m ouse (Herrm ann et al., 1990), Xenopus laevis {Xbra\ Smith et al., 1991), zebrafish Brachydanio rerio {Zf-T; Schulte- M erker et al., 1992), the ascidians H alocynthia roretzi (As-T; Yasuo and Satoh, 1994) and Ciona intestinalis {Ci-Bra', Corbo et al., 1997), sea urchin
H em icentrotus pulcherrim us {HpTa\ H arada et al., 1995), chicken (Ch-T;
sequences vary in length, encoding proteins of 4 3 2 -4 3 6 amino acids (aa) with
the exception of amphioxus A m Bra (440 aa), ascidian A s-T (471 aa) and
zebrafish Zf-T (423 aa). The overall amino acid identity varies betw een
64% —91 % am ongst vertebrates and is lower betw een vertebrates and
invertebrates (40% -63% ). Across the T dom ain the identity is particularly
high even betw een species o f different subphyla of the chordates (for example,
78.5% identity betw een ascidian and mouse proteins) or species o f different
phyla (for exam ple, 70% identity betw een sea urchin and m ouse proteins).
Inform ation on the genomic structure of T is only available for the m ouse and hum an orthologues. The hum an T gene is 10 kb long and
com prises eight exons (Fig 1.1; Edw ards et a l , 1996). The overall structure appears to be sim ilar to that described for the m ouse T gene (Stott et al.,
1993). However, a direct com parison is not possible, since details on
exon/intron boundaries and sizes have only been reported for the hum an gene
and the m ouse genomic structure has only been referred to schem atically in
Stott et al. (1993) and H errm ann and Kispert (1994). Although genom ic clones o f other orthologues have been isolated (zebrafish, am phioxus, Ciona),
no inform ation on the genomic structure o f these genes has been published.
1.1.2 r expression
Expression o f the T gene is largely confined to the early stages of developm ent. M ost inform ation has been derived from the studies o f mouse,
Xenopus and zebrafish T genes with more lim ited inform ation from hum an, chicken and prim itive chordates.
In the mouse, T expression is first detected in the early prim itive streak at 6.5 dpc. T mRNA appears transiently in the nascent and migrating
m esoderm generated by the prim itive streak. Expression ceases as the cells
m ove aw ay from the streak to form paraxial or extraem bryonic tissues. The
allantois, w ith the exception of the very early basal com ponent, does not
express T. In the prim itive streak and its derivatives, the node, notochord and tail bud, expression continues at a high level until m orphogenesis is com plete
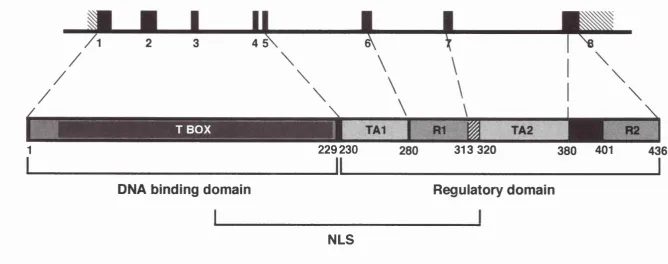
1Kb
TBOX
229230
313 320
380
401
DNA binding domain
I_ _
Reguiatory domain
NLS
Figure 1.1
Schematic representation of the
T
gene and its protein, showing the positions of the DNA binding and
(W ilkinson et al., 1990; Herrmann, 1991; Beddington et al., 1992; W ilson et al., 1993). A t later stages, T expression can be detected in the m ature notochord cells of the nucleus pulposus but not in adult tissues (intestine,
stomach, thymus, brain, testis, ovary, kidney, heart, adrenal; W ilkinson et a l,
1990).
Areas of T protein accum ulation overlap with those o f mRNA
expression but appear to be wider and not strictly correlated with the
com m itm ent of cells to mesoderm. In addition to the nascent and m igrating
m esoderm , T protein is also detected transiently in early endoderm and in
prospective neuroectoderm after 8.5 dpc, during the late stages o f the prim itive
streak. T protein is also present in m igrating extraem bryonic mesoderm,
possibly the precursor cells o f the allantois. W hile T protein accum ulates in
these non-m esoderm al sites, mRNA has not been detected. A nother difference
betw een T protein and mRNA distribution is that although T transcripts can be detected in the intervertébral notochord cells at 14.5 dpc, T protein disappears
in a rostrocaudal direction and is undetectable at this stage (K ispert and
Herrm ann, 1994).
The pattern of expression of the chicken T hom ologue (Ch-T) is very sim ilar to that in mouse. However, both C h-T transcripts and protein are detectable transiently in differentiated non-m esoderm al cells o f the
neuroectoderm and paraxial endoderm (Kispert et al., 1995b; Knezevic et al.,
1997).
Hum an em bryos are inaccessible during the very early stages of
developm ent, thus, although the expression pattern o f hum an T is likely to be sim ilar to that of the m ouse gene during gastrulation, data is not available. T
m RN A is readily detected in 13- and 14-w eek gestation hum an intervertébral
notochord, a stage of developm ent equivalent to 17.5 dpc in the mouse.
H um an T, like m ouse T, is confined to the notochord and was not detected in hum an adult gut, adult and fetal muscle, erythroid and intestinal cell lines and
14-w eek gestation spinal cord (Edwards et al., 1996).
The patterns of expression o f the T hom ologues in Xenopus (Xbra) and zebrafish {Zf-T) are very sim ilar to that of the m ouse T. Xbra is expressed throughout the m arginal zone and in the notochord and posterior m esoderm .
The m arginal zone, the equivalent o f the prim itive streak, is m ade up of
undifferentiated cells that give rise to m esoderm and endoderm cells during
gastrulation (Smith et aL, 1991). Similarly, Z f-Tis expressed in the germ ring, w hich gives rise to m esoderm and endoderm , and the notochord and posterior
m esoderm . m RN A and protein accum ulate in the m arginal zone and germ
ring transiently during gastrulation, but in the notochord and posterior
m esoderm persist until later stages of developm ent (Schulte-M erker et a l ,
1992).
Therefore, it seems that in all vertebrates the expression of T
hom ologues is conserved and this extends to the cephalocordate am phioxus
w hose em bryos and larvae express two T hom ologues A m B ra-1 a.ndAmBra-2.
U sing a cRN A probe that does not distinguish betw een A m B ra-1 and Am Bra-2, expression was identified in presum ptive m esoderm in early stages of developm ent and in posterior m esoderm and the notochord in late gastrulation
and neurulation (Holland et al., 1995). However, in the protochordates
H alocynthia roretzi {As-T) and Ciona intestinalis {Ci-Bra) the expression pattern appears to be different. A s-T and Ci-Bra are expressed only in notochord cells and not m esenchym e cells, although notochord and
m esenchym e cells, as well as muscle and nerve tissue, are derived from the
same cell lineage (Yasuo and Satoh, 1994; Corbo et a l , 1997).
Little is know n about the expression of Tin echinoderm s. In the sea urchin, H pTa expression is detected transiently in the archenteron and then at a high level at the tip of the archenteron and secondary m esenchym e cells
m igrating from there (H arada et a l , 1995). However, it is not yet clear w hether the archenteron and secondary m esenchym e cells are prim itive
equivalents o f the prim itive streak and notochord.
chordates. In Drosophila, for example, the Trelated gene Trg is expressed in the hindgut and anal pads (Kispert et a i , 1994).
The difference in T expression between the vertebrates and am phioxus on one hand, and the urochordates on the other, led to some interesting
speculations. It is possible that the prim itive streak and posterior m esoderm al
expression o f T in vertebrates and amphioxus, is related to evolutionary
progress and represents an acquired property associated w ith the possession of
a tail (H olland et a l, 1995). M oreover, it also seems possible that T
expressions in the prim itive streak and in the notochord w ere acquired
separately during evolution and m ay consequently be regulated by different
factors.
1.1.3 R egulation o f T expression
Tis required for normal m esoderm form ation and w hen over-expressed causes ectopic m esoderm formation. However, little is know n about how the
T gene is regulated and what factors determ ine prim itive streak and notochord expression. M ost work in this area has been carried out in X e n o p u s^ q c2l\xsq
the em bryos are easily manipulated and cultured.
In Xenopus, Xbra expression can be induced in anim al pole cells by vegetal pole cells, which are known to induce m esoderm form ation via grow th
factor-like signals. Xbra is activated by exogenous m esoderm inducing
proteins o f the fibroblast growth factor (FGF) and transform ing grow th factor-
13 (TGF-(3) families, bFG F and activin A respectively. Xbra expression is reduced w hen abnormal truncated versions of FGF and activin receptors are
over-expressed, presum ably because the active ligands are rem oved by the
truncated non-functional receptors (Smith et a l , 1991; Schulte-M erker and Smith, 1995). Xbra induction can occur in dispersed cells, is rapid and takes place even in the presence o f a protein synthesis inhibitor (cyclohexam ide).
This suggests that Xbra is an im m ediate early response gene of m esoderm induction (Smith et a l , 1991). Although FGF and activin can activate X bra
expression, continuous expression requires an intact FG F signalling pathw ay.
Expression o f Xbra declines when cells are dispersed and the intercellular link is diluted, but is m aintained when bFG F is added to the m edium o f the
dispersed cells (Isaacs et a l, 1994; Schulte-M erker and Smith, 1995). However, initial activation of Xbra by activin does not require a functional FG F signalling pathway (Schulte-M erker and Smith, 1995). Sim ilar pathw ays
are seen in other organisms. For example in chicken, C h-T is also induced by FG F and activin, but the induction is not an im m ediate early response
(Knezevic et a l , 1997). In zebrafish, Z f-T expression in anim al poles is induced by m arginal zone cells and exogenous activin A (Schulte-M erker et al., 1992). In Halocynthia, A s-T expression in the notochord can be induced by FG F (Nakatani et a l , 1996).
Em bryonic (e)FGF and Xbra can activate the expression o f each other
and eFG F maintains Xbra expression in dispersed animal pole cells (Isaacs et a l , 1994; Schulte-M erker and Smith, 1995). Xbra can activate its own expression but autoinduction requires protein synthesis and intercellular
signals (Rao, 1994; Tada et a l , 1997). This suggests that eF G F and Xbra are part of an autoregulatory loop in which X bra activates the expression o f
eFGF, which, in turn, maintains Xbra expression (Isaacs et a l , 1994; Schulte- M erker and Smith, 1995).
However, there is some evidence against the idea o f an autoregulatory
loop. Studies of chimaeric mice (see section 1.1.5) show ed
that T prom oter activity is m aintained in the prim itive streak and tail bud in
jBnuiiijBnuii carrying a T prom oter/lacZ construct up to the 14th day of gestation and thus, no T-dependent m echanism is required to m aintain
expression from this prom oter in T null regions. A ctivity from the T prom oterA acZ construct is apparent in null cells some distance (8 cell
diam eters) from neighbouring wild-type cells in the tail bud, and the level o f
expression o f the lacZ reporter gene is the same regardless o f the distance from T-expressing cells. This suggests that a T-dependant short-range
Furtherm ore, in embryos, which express a non-functional truncated
form o f the protein, Texpression ceases in early somite stage, indicating that up to this point w ild-type T is not required to m aintain its ow n expression
(Herrm ann, 1991; Kispert and Herrmann, 1994). The same is true for
zebrafish no tail mutants {ntl, functional T deficient), w hich continue to express m utant protein in the absence of functional protein (Schulte-M erker et a l , 1994). In Xenopus, expression o f a synthetic Xbra construct containing a repressor elem ent down-regulates Xbra expression in the notochord, but does not influence Texpression in ventral m esoderm (Conlon et a l , 1996). Thus, it seems that in the whole embryo the regulation of T is com plex and it is not clear w hether an autoregulatory loop involving T and FG F is in operation (Smith, 1997).
Recently, more evidence supporting the idea o f an autoregulatory loop
involving Xbra and eFG F in the notochord and dorsal m esoderm has emerged. It was shown that inhibition of Xbra function, by injection o f a synthetic Xbra
construct containing a repressor element, inhibits eF G F expression in the notochord and dorsal m esoderm of early embryos and FG F induced anim al
caps (Casey et a l , 1998). Thus, X bra function is required for m aintenance of eFG F expression. Furthermore, it was shown that induction of eFG F by X bra does not require protein synthesis or an intact FG F signalling pathw ay but is
cell autonom ous and occurs through binding of Xbra to eF G F 5'-flanking sequences (see section 1.1.4).
In Xenopus, 2.1 kb of Xbra 5'-flanking sequence and 50 bp of 5'- untranslated region is sufficient to direct expression in the m arginal zone but
not the notochord. Sequences within 381 bp from the transcription start site
are sufficient for induction of Xbra by activin and FGF. Response to activin is dose dependent and, at low levels activin induces Xbra expression, but at high levels suppresses Xbra. As yet, nothing is known about the factors w hich m ediate activin activation of T, however, suppression is m ediated by the hom eodom ain proteins goosecoid, M ix .l and Xotx2. These hom eodom ain proteins w ere shown to dow n regulate expression o f X bra by direct interaction
with B icoid and A ntennapedia binding sites in its 5'-flanking sequence
(Latinkic et a l, 1997). A second study also identified goosecoid as a repressor o f Xbra but associated with a different binding site in the 5'-untranslated region o f Xbra (A rtinger et a l , 1997).
In Ciona, Ci-Bra is only expressed in the notochord and transcriptional repression is essential for this specific expression pattern. A 434 bp m inim al
enhancer has been identified 5' of the putative TA TA elem ent which contains
three regulatory domains; a distal repressor elem ent that excludes expression
in m esenchym e and muscle, a central elem ent required for notochord
expression and proxim al elements which m ediate expression in ectopic
m esoderm al lineages. Activation of the central elem ent possibly involves a
regulatory elem ent related to the recognition sequence o f the Suppressor o f H airless [Su(H)] transcription activator. Su(H), is localised in the nucleus in response to activation of the Notch receptor (Corbo et a i , 1997). In addition, two binding sites for the Ciona snail gene (Ci-sna) were identified in the 434 bp enhancer. Ci-sna is expressed in the tail m uscles and it was suggested that it directly represses Ci-Bra expression in those m uscles (Fujiwara et a l ,
1998
^
In the mouse, 430 bp of proximal 5' flanking sequence is sufficient to
direct T expression in the prim itive streak. Two distinct regions have been identified; elem ents necessary for initiation o f expression in the first
gastrulating cells are found betw een nucleotides -4 3 0 and -2 8 0 and elem ents
required for expression in the later stages are found betw een nucleotides -2 8 0
and -1 9 0 . Thus far, sequences which confer expression in the head process
and notochord have not been identified in 8.3 kb o f 5' sequence and 5 kb o f 3'
sequence, suggesting that additional cw-acting sequences are necessary for T
expression in those regions. Thus, axial and non-axial expression o f T, and early and late prim itive streak expression, are regulated by different
1.1.4 T as a transcription factor
T protein is localised in the nucleus (Schulte-M erker et al., 1992; Cunliffe and Smith, 1994; Kispert and Herrmann, 1994; Schulte-M erker et al.,
1994). This localisation is mediated by nuclear localisation sequences w hich,
in the mouse, are betw een amino acids 137 and 320 (Fig 1.1; K ispert et al.,
1995a).
The consensus DNA binding site for mouse T protein was identified by
in vitro selection o f DNA binding sites from a pool o f random 26-m er oligonucleotides. This is a nearly palindrom ic consensus sequence o f 20 bp
with a core sequence A G GTG (TC/GA CACCTA G G T G T G A A A T T ). It was
shown that a half-site o f 12 bp (underlined) was not sufficient for binding in vitro and a m inim um o f six bases on one half-site in com bination w ith a com plete second half-site was required (Kispert and Herrm ann, 1993). C o
transfection experim ents showed that T can transactivate reporter genes
through binding to variously oriented and spaced half-sites. In general, T was
shown to transactivate better through binding to directly repeated dodecam ers
than inverted repeats, although, the highest activity was obtained through
binding to an inverted repeat of the dodecam er with a 24 bp spacer (K ispert et al., 1995a). A perfect palindrom e o f the half-site, flanked by Sm al half-sites (BS.p: 5 -G G G A A TTTCA CA CCTA G G T G T G A A A T T CCC-3 ) has been
w idely used in DN A /protein binding studies. Xenopus, zebrafish and
am phioxus T proteins also bind to the same BS.p DNA target in vitro (K ispert and Herrm ann, 1993; Herrm ann and Kispert, 1994; Holland et al., 1995).
T binds to its target DNA via the N-term inal 229 am ino acid residues,
the T dom ain or T-box (F ig l.l). Deletion of amino acids from the N- or C-
term inal o f this dom ain weakens or abolishes binding. In vitro D N A /protein binding studies have shown that T protein can bind to the BS.p palindrom e as
a m onom er, although, in this early work, the possibility that T can bind as a
dim er in vivo was not excluded (Kispert and Herrmann, 1993). D uring the course o f the project described here, the crystallographic structure o f the T
dom ain o f X bra in com plex with a 24 bp palindrom ic D N A target was
described and dem onstrated that T binds to DNA as a dim er (M üller and
Herrm ann, 1997). However, other studies have shown that X bra can regulate
transcription through binding to sites that com prise half of the BS.p
palindrom ic DNA target (Casey et al., 1998; Tada et al., 1998). These findings are discussed further in section 5.5.1.
Co-transfection experiments and the use o f heterologous systems
showed that T has a m odular structure. In addition to the N -term inal DNA
binding domain, the C-terminal half of the mouse protein contains two
transactivation (TA) domains, which act autonom ously and have an additive
effect; T A l, amino acids 2 3 0 -2 8 0 and TA2, amino acids 313-380. There are
also two repression (R) domains; R l, amino acids 2 8 1 -3 2 0 and R2, am ino
acids 4 0 1 -4 3 6 (Fig 1.1). The exact boundary betw een R l and TA 2 is not
know n (am ino acids 313-320; Kispert et al., 1995a).
Using a sim ilar approach it was shown that Xenopus Xbra and zebrafish Zf-T proteins also activate expression of reporter genes. However, a single
transactivation dom ain was identified in the carboxyl-term inal of these
proteins, m apping in X bra between amino acids 303 and 387, and in Zf-T
betw een am ino acids 312 and 407 (Conlon et al., 1996). Interestingly, the single transactivation domains of X bra and Zf-T map to the region of the
m ouse TA 2 domain, suggesting that this domain has been better conserved in
evolution than T A l.
The m odular structure o f T has been confirm ed using various m utant
forms o f the protein. For example, when proteins corresponding to the
zebrafish no tail m utations ntl^^^^ and ntf^^^ were used in cotransfection experim ents, no transactivation was observed (Conlon et al., 1996). ntl^^^° protein is 245 amino acids long and, although it retains its DNA binding
activity, it lacks a transactivation domain, ntl'’^^^ contains only 103 am ino
acids of the Zf-T protein, and thus lacks both a transactivation dom ain and an
the D rosophila engrailed protein is unable to activate transcription in transfection assays but competes with w ild-type X bra protein for D N A
binding (Conlon et a l , 1996).
This structure/function data for T protein bas been determ ined in vitro
and in cultured cell systems and therefore, should be treated with som e caution
since they m ight not necessarily mimic the situation in vivo. For exam ple, Rao (1994) showed in Xenopus animal cap assays that a truncated X bra protein o f 344 am ino acids, which lacks the C-terminal half o f the
transactivation dom ain, behaves like full-length X bra and induces m esoderm
form ation. Further truncation of X bra to 304 amino acids (B304) resulted in a
change from m esoderm to neural induction activity, w hich appears to
antagonise the m esoderm inducing activity o f full-length Xbra. Furtherm ore,
in the presence o f FG F or a low concentration of activin, the neuralising
activity o f B304 is increased.
The identification of target genes regulated by T has been relatively
slow. Very recently, using oocyte and cell culture transfection systems, X bra
has been show n to directly activate the expression o f eFG F (Casey et a l,
1998) and B ix l {Brachyury induced hom eobox gene 1; T ada et al., 1998). M is-expression o f B ix l can induce m esoderm or endoderm form ation in anim al pole tissue in a dose-dependent fashion and ventralisation o f dorsal
m esoderm in the m arginal zone.
1.1.5 The function of T and its role in embryogenesis
A m ajor source of inform ation about the in vivo function o f T protein comes from studies o f the phenotypes o f Tm utant and chim aeric mice.
The m ost com m only studied Tm utant is T Brachyury
w hich is due to a large deletion (1 8 0 -2 0 0 kb) encom passing the entire gene
and extensive flanking sequences. Hom ozygous m ice die at m id
gestation due to a defective allantois. The prim itive streak is thickened and
the node is not properly formed. As a result, somites posterior to the seventh
pair are absent or abnorm al and the notochord does not form. T acts in a dose
dependent fashion such that heterozygous m ice have short tails and the
notochord in the caudal region is branched or fused with the neural tube or gut
(Grüneberg, 1958). Thus, with the exception o f the allantois, the expression
pattern of T is consistent with the m utant phenotype (see section 1.1.2).
Sim ilarly, zebrafish hom ozygous no tail mutants (functional T null alleles) are lethal and lack a tail, a differentiated notochord and more than one third o f the
m ost posterior somites (H alpem et al., 1993; Schulte-M erker et al., 1994). These observations suggest that T plays a role in norm al posterior m esoderm
form ation and notochord differentiation.
The short-tail phenotype of the heterozygous m ice can be
rescued by the addition of a single copy o f a T transgene, verifying the dose
dependent action of T. Furtherm ore, transgenic studies suggest that a dosage
o f T higher than normal can increase embryonic lethality (Stott et al., 1993). It was proposed that over-expression of T could lead to aberrant cell type specification in the mesoderm, eventually leading to death. Interestingly, the
prom oter used to drive the expression of the transgene in these experim ents
has subsequently been shown to drive expression only in the prim itive streak
(Clem ents et al., 1996).
Studies o f other T mutants, which are not due to large deletions, also highlight the im portance of T dosage and led to hypotheses that T interacts with other proteins. is a m utant due to an insertion o f a retroviral-like
elem ent in the seventh exon and 7^ involves a 19 bp deletion in exon 8
(review ed in Herrm ann and Kispert, 1994). Both these m utations alter the
open reading fram e resulting in proteins that retain their D N A binding
property but are truncated at the carboxy terminus of the regulatory domain.
T^*® is lacking the TA 2 and R2 domains and T^ is lacking the R2 dom ain (see
previous section). Both and T m utant m ice exhibit a m ore severe phenotype than mice; homozygotes have no somites, the rostral
boundary o f em bryonic defects extends into the cervical region and
(M acM urray and Shin, 1988; Herrmann, 1991; K ispert and Herrm ann, 1994).
In addition, the T / + phenotype, but not the 7^/7^ phenotype, is dependent on the num ber of wild type T copies and two are required to rescue the tailless phenotype (M acM urray and Shin, 1988; Stott et a l , 1993). It is possible that
and proteins antagonise the wild type protein, effectively reducing the
am ount o f functional protein. There is an alternative explanation for the m ore
severe phenotype o f and ' f . If T interacts with other proteins to form a transcriptional complex, then a truncated T protein, which nevertheless binds
DNA, could interfere with the assembly or function o f this transcriptional
com plex. This interference in not a feature of mutants w hose T protein is
com pletely absent.
Further clues about the way that T protein exerts its influence on
m esoderm cell differentiation and m igration come from studies o f chim aeric
m ice generated by injecting embryonic stem (ES) cells into
blastocysts. chimaeras with high levels (>70% ) o f null alleles
are alm ost indistinguishable from intact hom ozygous mutants, w hereas low
level chim aeras show a range of defects that can vary from a severe
phenotype, sim ilar to that of mice, to localised defects in the tail
(truncation, branching) or allantois. In all chimaeras there is a rostrocaudal
increase in T null cell contribution and the level of chim aerism in the
neuroectoderm is higher than that in paraxial mesoderm. W here heterozygous
jSnuii/_^ cells w ere used to generate chimaeras, 7^”“^V+x+/+ mice are norm al with m ild tail defects (kinking, abnormal tail tip) and cells can colonise
rostral regions efficiently. The phenotypes of these chim aeric m ice support
the idea that T acts in a dose dependent, cell autonom ous w ay (Rashbass et al.,
1991; Beddington et a l , 1992; W ilson et al., 1993; W ilson et al., 1995). Careful analysis o f chimaeras and com parison with wild type and
jBnuiiijBnuii j^ytants showed that the cells fail to m igrate aw ay from the prim itive streak and subsequently accum ulate in the node, prim itive
streak and later in the tail bud, leading to abnorm alities in the notochord and
posterior m esoderm (Rashbass et a l , 1991; Beddington et al., 1992; W ilson et a l , 1993; W ilson et a l , 1995). It was suggested that T affects cell m igration by regulating genes involved in cell adhesion, an idea supported by other
observations. Brachyury m ouse mutants have a thickened prim itive streak and a reduced m esoderm /ectoderm ratio, which appears to be due to abnorm al cell
m ovem ent during gastrulation (Y anagisawa et a l , 1981). In a separate series o f cell culture experim ents (reviewed in W ilson et a l , 1993), it was shown that w hen cells were com pared with + /+ cells, they show reduced
m otility on extracellular m atrix and form smaller aggregates. However,
jSnuiiijBnuii tiave the same rate o f DNA synthesis and m itotic index as wild type cells, and in culture, they have the same differentiative capacity and
their life-span extends beyond the tim e of death in the embryo.
Thus, it seems unlikely that the abnormal behaviour o f cells in vivo can be explained by differential cell growth, aberrant cell differentiation, or low
survival time.
Interestingly, a 5 integrin deficient mice resem ble hom ozygous
Brachyury m ice (Yang et a l , 1993). a5(3l integrin is a cell surface receptor that binds fibronectin and is thought to be involved in m any cellular processes
including cell migration. In hom ozygous a 5 integrin null m ice the anterior
part o f the embryo from somites 1 -1 0 develops normally, but the posterior
regions are highly defective. Posterior somites are not form ed, paraxial
m esoderm is reduced and the neural tube is kinked. The notochord is formed,
but it is not know n if it is functional.
Analysis of chimaeras of wild-type cells and cells carrying a
w ild type T transgene, showed that the expression o f T in cells restores their normal behaviour in the prim itive streak and tail bud.
Furtherm ore, these experiments suggested that the level o f T expression determ ines the speed with which cells move away from the prim itive streak.
streak, w hereas cells with low levels of T tend to stay in the prim itive streak
for a longer time and populate the tail bud (W ilson and Beddington, 1997).
M uch o f w hat has been proposed regarding T and m esoderm
differentiation and migration refers to the situation in the prim itive streak and
node. However, it is not clear how this applies to the notochord or tail bud.
A lthough, the supply of cells to the axial mesoderm, and thus the notochord, is
directly dependent on the prim itive streak and node, it is likely that T protein
m ay have a distinct function in the differentiation and survival of notochordal
cells. D isruption o f these functions would lead to the notochord abnorm alities
seen in zebrafish and mouse T mutants and chim aeric m ice (Beddington et al.,
1992; W ilson et al., 1995). Similarly, although the prim itive streak is directly related to the tail bud, it has been proposed that T has a distinct role in tail bud function and affects tail differentiation and elongation. It follows from this
proposal that the abnormal tail m esoderm form ation seen in m ice is
a prim ary outcom e o f T deficiency and not a secondary response to a
degenerate notochord (W ilson et al., 1993; W ilson et al., 1995).
The im portance o f T in m esoderm form ation is also highlighted by
expression studies in Xenopus. O ver-expression of Xbra in animal caps cause the ectopic form ation of posterior mesoderm. M esoderm al markers that are
indirectly activated by X bra are Xsna, a zinc-finger transcription factor,
m uscle-specific actin and Xhox3, a hom eobox transcription factor expressed in posterior m esoderm (Cunliffe and Smith, 1992). The type o f m esoderm
form ed depends on the dose o f Xbra RNA, with low am ounts inducing m ésothélial smooth m uscle and m esenchym e and high am ounts inducing
somitic m uscle (O'Reilly et a l , 1995).
Interestingly, the outcom e of Xbra over-expression is influenced by co expression with a variety o f other transcription factors and inducers, im plying
that in vivo several genes co-operate with Xbra to specify dorsal m esoderm . These collaborating factors probably include Pintallavis, a m em ber o f the forkhead/H N F3p transcription factor fam ily (O'Reilly et al., 1995), noggin, a bone m orphogenetic protein (BMP) antagonist and Xwnt-8, a m em ber o f the
w nt fam ily o f secreted glycoproteins (Cunliffe and Smith, 1994). Changing
the level of co-expression and the order o f expression of these factors
influences whether muscle, notochord and/or neural tissue are form ed and
w hether dorsalisation of m esoderm occurs.
Recently, expression studies in the ascidian Halocynthia roretzi have shown that the role o f T in notochord form ation is probably conserved in all
chordates (Yasuo and Satoh, 1998). O ver-expression of A s -T is sufficient for notochord differentiation of notochord precursors in a cell-autonom ous way.
However, in a sim ilar way to Xbra, the response to over-expression in non notochord lineages depends on the presence of other factors.
1.1.6 The T-box transcription factors
r is a m em ber of a growing fam ily of transcription factors, the T-box
genes. Thus far, 13 T-box genes have been identified in m am m als; T, Tbxl-8, TbxlO, T bxlS, T brl and Eomes (Herrm ann et al., 1990; Bollag et al., 1994; Bulfone et a l , 1995; Agulnik et a l , 1996; A gulnik et al., 1998; Law et al.,
1998; W attler et al., 1998). The hom ologues of 8 of these genes have been cloned in hum ans; T, TBXl-3, TBX5, TBXIO, TBX15 and T B R l (Bulfone et a l ,
1995; Cam pbell et a l , 1995; Edwards et a l , 1996; Bam shad et a l , 1997; Basson et a l , 1997; Chieffo et a l , 1997; Li et a l , 1997; A gulnik et a l , 1998; Law et a l , 1998). The identification of T-box genes in other species as
divergent as C.elegans, Drosophila, Xenopus, chicken etc. suggests that the T- box fam ily of transcription factors is evolutionarily conserved. Phylogenetic
analysis and com parison o f the genomic structure o f the T dom ain have shown
that the T-box genes can be divided into subfamilies. It has been proposed
that m ost anim al species are likely to have at least five T-box genes and these
will be related to the T, T b xl, Tbx2, Tbx6 and T brl of the m ouse (Papaioannou and Silver, 1998).
The characteristic feature of T-box proteins is the DNA binding
domain, a conserved region o f 180-200 amino acid residues, w hich although
relative to the 5' end. Both DNA-binding and gene activation properties have
been directly dem onstrated in T hom ologues from mouse, Xenopus and
zebrafish (Kispert and Herrmann, 1993; Herrm ann and Kispert, 1994; K ispert
et a l , 1995a; Conlon et a l , 1996), in mouse Tbx2 (Carreira et a l , 1998) and in Xenopus V egT (Zhang and King, 1996; Tada et a l , 1998). DNA binding has been dem onstrated for Drosophila omb (Pflugfelder et a l , 1992) and am phioxus A m Bra (Holland et a l , 1995) and transcriptional regulation has been shown for the zebrafish spadetail, spt (Griffin et a l , 1998). A lthough there is no other direct evidence for the transcriptional properties o f other T-
box proteins, it is likely that they will all have DNA -binding and
transactivation properties.
M ost inform ation on the expression patterns o f different T-box genes
comes from mouse and chicken, but it is reasonable to assum e that these
patterns would be sim ilar in other species, taking into account differences in
m orphology and tim e of development. Indeed, w here orthologues in different
species have been identified, the expression patterns are similar.
The T-box genes have distinctive but overlapping patterns of
expression. They are generally most abundant during em bryonic
developm ent, although some persist in adult tissues (for exam ple Tbx2 and
Tbx3, Table 1.1). A t the time of murine organogenesis there are at least two T-box genes expressed in all the m ajor organs, but not necessarily in the same
cell type (Bollag et a l , 1994; Bulfone et a l , 1995; A gulnik et a l , 1996; Chapm an et a l , 1996b). For example, Tbx2-5 are expressed in lung
m esenchym e, whereas, T bxl is expressed in lung epithelium . An overlap in the expression pattern of different genes is evident in m em bers o f the same
subfam ily; for exam ple Tbx2 and Tbx3 are coexpressed in kidney and
peripheral nervous system (PNS), whereas, Tbx4 and Tbx5 are coexpressed in extraem bryonic m esoderm and heart atrium (Table 1.1).
The im portance of the T-box genes in hum an developm ent has been
highlighted by the identification o f disease-associated m utations in two genes.
M utations in TBX3 cause Ulnar-M am m ary syndrome, an autosom al dom inant
Stage & Tissue T Tbxl Tbx2 Tbx3 Tbx4 Tbx5 Tbx6 T brl
Blastocyst
Inner cell mass - - - + - -
-Gastrulation
Extraembryonic mesoderm - - - + + +
-Embryonic mesoderm + + - - - - +
Notochord + - - - - -
-Organogenesis
CNS - - + + - - - 4
-PNS - - + + - - -
-Body W all, thorax - - - - - + -
-Cartilage/skeleton - + + + - - -
-Eye - - + + - + -
-Genital papilla - - + + + + -
-Genital ridge - - + + - - -
-Heart
Atrium - - - - + + -
-Ventricle - - - - - + -
-Kidney - - + + - - -
-Limbs
Forelimb - - + + + + -
-Hindlimb - - + + + - -
-Lungs
M esenchym e - - + + + + -
-Epithelium - + - - - - -
-Mammary gland
M esenchym e - - + - - - -
-Epithelial buds - - - + - - -
-Mandible m esenchym e - - + + + + -
-M axilla m esenchym e - - + + - - -
-M uscle - - + + - - -
-Optic vesicle - - + + - + -
-Otic vesicle - + + + - - -
-Tail + - - - + - +
-Adult
Brain - - - + +
Heart - - + +
Kidney - - + +
Liver - +
Lungs - - +
Ovaries - - + +
Testes - + -