1535-9778/05/$08.00⫹0 doi:10.1128/EC.4.12.1971–1981.2005
Copyright © 2005, American Society for Microbiology. All Rights Reserved.
Pde1 Phosphodiesterase Modulates Cyclic AMP Levels through a
Protein Kinase A-Mediated Negative Feedback Loop in
Cryptococcus neoformans
Julie K. Hicks,
1Yong-Sun Bahn,
1and Joseph Heitman
1,2,3*
Departments of Molecular Genetics and Microbiology,1Medicine,2and Pharmacology and Cancer Biology,3
Duke University Medical Center, Durham, North Carolina 27710
Received 25 July 2005/Accepted 20 September 2005
The virulence of the human pathogenic fungus Cryptococcus neoformans is regulated by a cyclic AMP
(cAMP)-dependent protein kinase A (PKA) signaling cascade that promotes mating and the production of
melanin and capsule. In this study, genes encoding homologs of theSaccharomyces cerevisiaelow- and
high-affinity phosphodiesterases,PDE1andPDE2, respectively, were deleted in serotype A strains ofC. neoformans.
The resulting mutants exhibited moderately elevated levels of melanin and capsule production relative to the
wild type. Epistasis experiments indicate that Pde1 functions downstream of the G␣ subunit Gpa1, which
initiates cAMP-dependent signaling in response to an extracellular signal. Previous work has shown that the
PKA catalytic subunit Pka1 governs cAMP levels via a negative feedback loop. Here we show that apde1⌬
pka1⌬mutant strain exhibits cAMP levels that are dramatically increased (⬃15-fold) relative to those in a
pka1⌬single mutant strain and that a site-directed mutation in a consensus PKA phosphorylation site reduces
Pde1 function. These data provide evidence that fluctuations in cAMP levels are modulated by both Pka1-dependent regulation of Pde1 and another target that comprise a robust negative feedback loop to tightly constrain intracellular cAMP levels.
Signal transduction cascades are the primary means by which external cues are communicated to the nuclei of eukary-otic organisms. The resulting changes in gene transcription are manifested as an appropriate response to the given cue. The importance of signal transduction cascades is perhaps best reflected in the high degree of cascade component conserva-tion that exists between animals and fungi, despite the fact that they diverged from a common ancestor over 800 million years ago (29).
In fungi, mitogen-activated protein kinase and cyclic AMP (cAMP)-dependent signal transduction cascades are two major signal transduction conduits. The highly conserved compo-nents of the cAMP signaling cascade have been co-opted by many different fungi for a variety of purposes, including regu-lation of asexual and sexual spore production, mycotoxin pro-duction, pseudohyphal growth, mating, pathogenesis, and vir-ulence (1, 16, 17, 20, 32, 36, 42, 48).
The cAMP-dependent signaling pathway has been well char-acterized for the human-pathogenic basidiomycete fungus
Cryptococcus neoformans. Both serotype A (C. neoformansvar.
grubii) and serotype D (C. neoformansvar.neoformans) strains infect immunocompromised individuals and cause meningoen-cephalitis (6). The virulence of this fungus depends on several factors, including the ability to grow at 37°C and the produc-tion of melanin and a polysaccharide capsule (6). Addiproduc-tionally, the ability to mate and grow filamentously also likely has a role in survival and propagation in the environment and may gen-erate infectious propagules (23).
C. neoformansvirulence factor production and mating ability are regulated by the cAMP-dependent signaling pathway (for reviews, see references 12 and 29). Components of this path-way that have been identified include a G␣subunit of a het-erotrimeric G protein (Gpa1), adenylyl cyclase (Cac1), the PKA catalytic (Pka1 and Pka2) and regulatory (Pkr1) subunits, and an adenylyl cyclase-associated protein (Aca1) that works independently of Gpa1 to activate adenylyl cyclase (1–3, 13, 19) (Fig. 1). Deletion of theGPA1,CAC, orACA1gene results in a loss of mating and capsule and melanin production and attenuates virulence. These defects can be rescued by the ad-dition of exogenous cAMP (1–3). Deletion of the Pkr1 regu-latory subunit results in increased capsule production and hy-pervirulence (13). The roles of Pka1 and Pka2 in serotype A and serotype D have diverged. In serotype A, Pka1 plays the primary role in the regulation of mating, capsule and melanin production, and virulence, while Pka2 plays only an ancillary role (3, 13). In serotype D, Pka2 contributes to the regulation of virulence trait production but is not required for virulence (13, 19).
In Saccharomyces cerevisiae, intracellular cAMP levels are influenced by the activity of the low- and high-affinity phos-phodiesterases (PDEases), Pde1 and Pde2, respectively. These phosphodiesterases are structurally distinct, sharing virtually no sequence similarity. Pde2 is a high-affinity PDEase that is represented in organisms ranging from yeast to mammals (7). In contrast, Pde1, the low-affinity PDEase, is less well charac-terized but is found in a range of organisms, including S. cerevisiae,Schizosaccharomyces pombe,Candida albicans, Dic-tyostelium discoideum, Vibrio fischeri, Leishmania mexicana,
Trypanosoma brucei, andTrypanosoma cruzi(9, 11, 14, 22, 27, 28, 34, 39).
Pde1 and Pde2 have distinct functions. Pde2 regulates basal
* Corresponding author. Mailing address: Department of Molecular Genetics and Microbiology, 322 CARL Bldg., Duke University Med-ical Center, Research Dr., Durham, NC 27710. Phone: (919) 684-2824. Fax: (919) 684-5458. E-mail: [email protected].
1971
on September 8, 2020 by guest
http://ec.asm.org/
cAMP levels in bothS. cerevisiaeandC. albicans (5, 30). Be-cause the regulation of basal cAMP levels is important in determining tolerance to stress, including temperature ex-tremes and high salt and heavy metal concentrations, Pde2 has a pivotal role in stress tolerance (30, 37). InC. albicans, Pde2 is also involved in filamentation, nutrient sensing, entry into stationary phase, and cell wall and membrane integrity (5, 25). In contrast,S. cerevisiaePde1 does not significantly affect the basal levels of cAMP and does not confer any obvious mutant phenotype. However, in bothS. cerevisiaeandS.pombe, Pde1 does regulate cAMP levels induced by the presence of glucose (21, 30). Furthermore, the cAMP degradation activity of Pde1 in S. cerevisiae is positively regulated by the PKA catalytic subunits (35). The regulation of Pde1 activity is also observed
in S. pombe, but it is not clear if this regulation is effected directly via PKA catalytic or allosteric activation by cAMP, as occurs inD. discoideum(21, 31). In humans, it has been sug-gested that some PDEases may be regulated independently of PKA (8).
Here we report the characterization of Pde1 and Pde2 from
C. neoformans. We show that, unlike in S. cerevisiae and C. albicans, Pde2 has no apparent role in regulating intracellular cAMP levels, and deletion of the gene results in subtle mutant phenotypes. However, Pde1 clearly is part of the cAMP-de-pendent signaling pathway that regulates virulence attributes. Not only does Pde1 disruption rescuegpa1⌬mutant pheno-types, but we also provide evidence that Pde1 is activated by Pka1 and that it is an important part of the PKA-activated negative feedback loop that governs intracellular cAMP levels in this pathogenic fungus.
MATERIALS AND METHODS
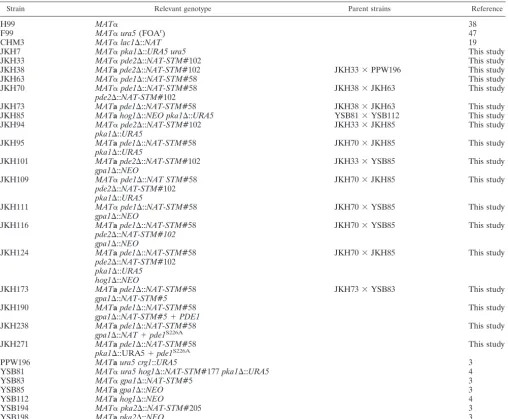
C. neoformansstrains and media.All strains used for this study are listed in Table 1.C. neoformansstrains were grown on standardS. cerevisiaemedium (41). Selective medium for biolistic transformation, Niger seed medium, V8 medium, and Dulbecco’s modified Eagle’s medium (DMEM) were prepared as described previously (1, 18, 44). For cAMP suppression experiments, cAMP was added at a concentration of 10 mM to the media.
pka1⌬::URA5transformants were selected on synthetic medium lacking uracil and containing 1 M sorbitol.pde1⌬::NATandpde2⌬::NATtransformants were selected on yeast extract-peptone-dextrose (YPD) medium containing 100g/ml nourseothricin.pde1⌬::NEOandpde2⌬::NEOwere selected on YPD medium containing 200g/ml G418. Genotypes were confirmed by both Southern hy-bridization and expression analysis.
Identification ofC. neoformans PDE1andPDE2genes.TheC. neoformans PDE1andPDE2genes were identified in serotype A and D strains by performing a tBLASTn search of the Duke BioinformaticsC. neoformansserotype A data-base (http://cneo.genetics.duke.edu/menu.html) and the Stanford Genome Tech-nology Center serotype D database (http://www-sequence.stanford.edu/group/C .neoformans/overview.html), using the S. cerevisiae Pde1 and Pde2 protein sequences.
Disruption ofPKA1,PDE1, andPDE2genes.Thepka1⌬,pde1⌬, andpde2⌬
null mutants were generated by overlap PCR as described previously (10). The primer sequences used for this study are available upon request. To construct the
pka1⌬::URA5allele, in the first round of PCR the 5⬘end of thePKA1gene was amplified with primers 9352 and 9354, the 3⬘end of the gene was amplified with primers 9355 and 9357, and theURA5gene was amplified with primers 9352 and 9356. Primers 9352 and 9357 were then used in an overlap amplification with the first three products as templates to yield the⬃4.0-kb PCR product bearing the
pka1⌬::URA5allele. The gel-extractedPKA1disruption cassette was precipitated onto 0.6-g gold microcarrier beads (0.8-m; Bioworld Inc.) and transformed into theMAT␣ura5serotype A strain F99 by biolistic transformation (44). The
pka1⌬::URA5allele deletes the entirePKA1open reading frame (ORF). ThePDE1andPDE2gene ORFs were disrupted by biolistic transformation of the congenicC. neoformansserotype A strains H99 and KN99awith constructs generated by overlap PCR (10). The 5⬘and 3⬘regions of thePDE1andPDE2
genes in serotype A strains were PCR amplified with the following primers: 10403/10404 and 10347/10348 for the 5⬘regions of thePDE1andPDE2genes, respectively, and 10405/10406 and 10349/10350 for the 3⬘regions ofPDE1and
PDE2, respectively. M13F and M13R primers were used to generate Natr dom-inant selectable markers from the template pNATSTM#58 (for PDE1) or pNATSTM#102 (forPDE2), with a unique signature tag (33), and Neor domi-nant selectable markers from the template pJAF1 (15). ThePDE1disruption alleles were generated by overlap PCR using primers 10403 and 10406. The
PDE2disruption alleles were generated by overlap PCR using primers 10347 and 10350. Prototrophic wild-type strains were biolistically transformed with the gel-extractedPDE1andPDE2disruption alleles as described previously (10).
pde1⌬andpde2⌬strains were screened by diagnostic PCR for the 5⬘and 3⬘
junctions and by Southern blot analysis using specific probes generated by PCR with the primers 10423/10424 and 11206/11207 for thePDE1andPDE2genes, respectively (data not shown).
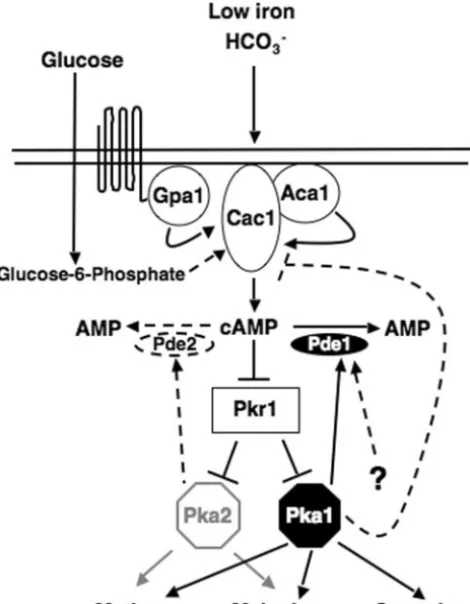
FIG. 1. Model for the role of Pde1 and Pde2 in the cAMP signaling cascade. Virulence factor production is regulated by the cAMP-depen-dent signaling pathway via the following components: an unicAMP-depen-dentified seven-transmembrane receptor(s); the G␣subunit (Gpa1); adenylyl cyclase (Cac1) and its associated protein, Aca1; PKA, composed of the Pkr1 regulatory subunit and the Pka1 and Pka2 catalytic subunits; and the PDEase Pde1. In serotype A, the Pka1 catalytic subunit plays a major role in regulating virulence factor production, while the Pka2 subunit plays only a minor role. Intracellular cAMP levels are modu-lated through a feedback loop controlled by Pka1. When active, Pka1 positively regulates Pde1 and may also negatively regulate Cac1, ex-erting control over both the production (Cac1) and degradation (Pde1) of cAMP. Although Pde2 has no apparent role in the regulation of intracellular cAMP levels, it is possible that Pde2 may be regulated by Pka2. Solid lines and arrows indicate experimentally proven portions of the pathway, and dashed lines and arrows indicate putative portions of the pathway for which there is, as yet, no experimental evidence. Gray arrows and lines indicate minor roles in the pathway.
on September 8, 2020 by guest
http://ec.asm.org/
Complementation of the pde1⌬ mutant.The serotype A PDE1 gene for complementation of thepde1⌬mutant was amplified via PCR using primers 13458 and 13459, with wild-type (H99) genomic DNA as the template. This
⬃3.8-kb product included the promoter, ORF, and terminator regions of the
PDE1gene and was initially cloned into the pCR2.1-TOPO vector (Invitrogen), creating plasmid pJH106. pJH106 was digested with ApaI and NotI, and the fragment was ligated into pJAF12 (3), creating plasmid pJH108. Strain JKH173 was transformed with plasmid pJH108, using the biolistic apparatus (44).
Site-directed mutagenesis.To mutate the putative PKA phosphorylation site in Pde1, we constructed plasmid pJH168, containing thepde1S226Amutation, via overlap PCR (10). An⬃1.3-kb SalI fragment comprising thePDE1sequence containing a mutation in the sole putative PKA phosphorylation site was created by first amplifying the regions upstream of the putative phosphorylation site (primers 14506 and 14509) and downstream of the site (primers 14507 and 14508). Thepde1S226Aallele was generated by overlap PCR with primers 14506 and 14507. This fragment was cloned into the pCR2.1-TOPO vector (Invitrogen) to create plasmid pJH155 and sequenced. pJH108 was digested with SalI, and the wild-type 1.3-kb fragment was replaced with thepde1S226A
-containing 1.3-kb SalI fragment from pJH155 to create plasmid pJH168. Strains JKH238 and JKH271, containing thepde1S226A
mutation, were generated by biolistically transforming plasmid pJH168 into strains JKH173 and JKH95, respectively. The presence of
the mutated allele in strains JKH238 and JKH271 was further confirmed by PCR amplification and sequencing of the relevant region.
Expression analysis.Fungal strains were inoculated into 5 ml YPD medium and grown overnight at 30°C. Fifty milliliters of YPD medium in a 125-ml flask was inoculated with 500l of overnight culture and grown at 250 rpm and 30°C for 5 h prior to harvesting. RNAs were isolated from the harvested cultures with Trizol (Gibco-BRL) according to the manufacturer’s instructions. Fifteen micro-grams (as indicated by spectrophotometric measurement) of RNA was separated in a denaturing gel and transferred to a nylon membrane. The resulting blots were probed withPDE1andPDE2gene-specific probes.
Capsule assays.To assess capsule and cell sizes, each strain was inoculated onto a plate of DMEM and grown at either 30°C (see Fig. 3) or 37°C (see Fig. 4 and 6) for 2 days. Aliquots of all cultures were stained with India ink and viewed with a 100⫻oil objective. Twenty cells from each culture were chosen at random, and the cell and capsule sizes were measured using Axiovision 3.0 software (Carl Zeiss Microscopy). The relative capsule diameter was calculated with the following equation: (Dw⫺Dc)⫻100/Dw, whereDwandDcindicate the
diameters of the whole-cell body (Dw; cell plus capsule) and the cell body only
(Dc) (49). The Bonferroni multiple comparison test was performed with Prism
4.0 software (GraphPad Software) to compare each strain to every other strain (3, 49). Each experiment was done in triplicate (totaln⫽60).
TABLE 1. Strains used for this study
Strain Relevant genotype Parent strains Reference
H99 MAT␣ 38
F99 MAT␣ura5(FOAr) 47
CHM3 MAT␣lac1⌬::NAT 19
JKH7 MAT␣pka1⌬::URA5 ura5 This study
JKH33 MAT␣pde2⌬::NAT-STM#102 This study
JKH38 MATapde2⌬::NAT-STM#102 JKH33⫻PPW196 This study
JKH63 MAT␣pde1⌬::NAT-STM#58 This study
JKH70 MAT␣pde1⌬::NAT-STM#58 JKH38⫻JKH63 This study
pde2⌬::NAT-STM#102
JKH73 MATapde1⌬::NAT-STM#58 JKH38⫻JKH63 This study
JKH85 MATahog1⌬::NEO pka1⌬::URA5 YSB81⫻YSB112 This study
JKH94 MAT␣pde2⌬::NAT-STM#102 JKH33⫻JKH85 This study
pka1⌬::URA5
JKH95 MATapde1⌬::NAT-STM#58 JKH70⫻JKH85 This study
pka1⌬::URA5
JKH101 MATapde2⌬::NAT-STM#102 JKH33⫻YSB85 This study
gpa1⌬::NEO
JKH109 MAT␣pde1⌬::NAT STM#58 JKH70⫻JKH85 This study
pde2⌬::NAT-STM#102 pka1⌬::URA5
JKH111 MAT␣pde1⌬::NAT-STM#58 JKH70⫻YSB85 This study
gpa1⌬::NEO
JKH116 MATapde1⌬::NAT-STM#58 JKH70⫻YSB85 This study
pde2⌬::NAT-STM#102 gpa1⌬::NEO
JKH124 MATapde1⌬::NAT-STM#58 JKH70⫻JKH85 This study
pde2⌬::NAT-STM#102 pka1⌬::URA5 hog1⌬::NEO
JKH173 MATapde1⌬::NAT-STM#58 JKH73⫻YSB83 This study
gpa1⌬::NAT-STM#5
JKH190 MATapde1⌬::NAT-STM#58 This study
gpa1⌬::NAT-STM#5⫹PDE1
JKH238 MATapde1⌬::NAT-STM#58 This study
gpa1⌬::NAT⫹pde1S226A
JKH271 MATapde1⌬::NAT-STM#58 This study
pka1⌬::URA5⫹pde1S226A
PPW196 MATaura5 crg1::URA5 3
YSB81 MAT␣ura5 hog1⌬::NAT-STM#177pka1⌬::URA5 4
YSB83 MAT␣gpa1⌬::NAT-STM#5 3
YSB85 MATagpa1⌬::NEO 3
YSB112 MATahog1⌬::NEO 4
YSB194 MAT␣pka2⌬::NAT-STM#205 3
YSB198 MATapka2⌬::NEO 3
on September 8, 2020 by guest
http://ec.asm.org/
Laccase assays.To quantitate melanin production, the wild-type (H99) and
pde1⌬(JKH63),pde2⌬(JKH33),pde1⌬pde2⌬(JKH70), andlac1⌬(CHM3) mutant strains were grown in 25 ml ofL-3,4-dihydroxyphenylalanine (L-DOPA) medium (0.1%L-asparagine, 0.1% glucose, 0.3% KH2PO4, 0.01% L-DOPA, 0.025% MgSO4· 7H2O; pH 5.6) at 30°C and 250 rpm for 16 h. The cultures were then incubated at 25°C and 250 rpm for 24 h, at which time 1 ml of culture was centrifuged, and the supernatant was spectrophotometrically read for the optical density at a wavelength of 475 nm (OD475).
cAMP assays.Examinations of intracellular cAMP levels were performed by growing the wild-type (H99) and pka1⌬ (JKH7), gpa1⌬ (YSB83), pde1⌬
(JKH63),pde2⌬(JKH33),pde1⌬pde2⌬(JKH70),pde1⌬pka1⌬(JKH95),pde2⌬
pka1⌬(JKH94),pde1⌬pde2⌬pka1⌬(JKH109),pde1⌬gpa1⌬(JKH111),pde2⌬ gpa1⌬(JKH101), andpde1⌬pde2⌬gpa1⌬(JKH116) mutant strains in 50 ml of YPD medium for 24 h. The cultures were centrifuged and washed twice with double-distilled water (ddH2O) and once with MES buffer (10 mM morpho-lineethanesulfonic acid [MES; pH 6.0], 0.5 mM EDTA) before being resus-pended in MES buffer. Fifteen milliliters of MES buffer was then inoculated in order to give the cultures a final OD600of 2.0. Following 2 h of incubation at 30°C and 250 rpm, 1-ml samples of each culture were collected via filtration on 0.45-m filters 0 min, 0.5 min, 1.0 min, and 3.0 min after the addition of 2% glucose. The cell-containing filters were incubated in a petri plate containing 1 ml of formic acid:butanol:ddH2O (3.7:76.3:20) for 1 h prior to transfer of the liquid to a microcentrifuge tube, centrifugation at 13,000 rpm for 10 min, and transfer of the supernatant to a new tube. The resulting liquid was lyophilized, at which time it was resuspended in the assay buffer supplied with the cAMP Biotrak enzyme immunoassay system (Amersham Biosciences). The assay was performed according to the manufacturer’s instructions.
Nucleotide sequence accession numbers.ThePDE1andPDE2serotype A gene sequences have been deposited in the NCBI GenBank database and have the following accession numbers: AY864841 (PDE1A) and AY874131 (PDE2A). ThePDE1andPDE2gene sequences from serotype D have also been deposited in the NCBI GenBank database and have the following accession numbers: AY864842 (PDE1⌬) and AY874132 (PDE2⌬).
RESULTS
C. neoformanscontains homologs of low- and high-affinity
PDEases, Pde1 and Pde2.S. cerevisiaeexpresses two PDEases,
the low-affinity form, Pde1, and the high-affinity form, Pde2. Pde1 functions in response to agonist-induced increases in cAMP, while Pde2 regulates basal cAMP levels (30). Using the
S. cerevisiaePde1 and Pde2 protein sequences, we performed a tBLASTn search to identify C. neoformans serotype A and serotype D Pde1 and Pde2 homologs. One sequence for each protein was identified from each database and designated Pde1 and Pde2. Both Pde1 and Pde2 have the conserved signature
sequences for the low- and high-affinity PDEases, respectively (Fig. 2). The serotype A and D Pde1 proteins share 94% identity, and the serotype A and D Pde2 proteins share 93% identity. In contrast, the Pde1 and Pde2 proteins share only 12 to 14% identity, similar to the Pde1 and Pde2 proteins ofS. cerevisiae(13% identity).
Deletion ofPDE1andPDE2enhances melanin and capsule
production.The entire ORFs of thePDE1 and PDE2genes
were deleted from a serotype A wild-type strain to examine their role in the production of virulence factors. Expression analysis confirmed that the entire ORF had been deleted in both cases (data not shown). The wild-type strain and the
pde1⌬, pde2⌬, and pde1⌬ pde2⌬ mutant strains were inocu-lated on Niger seed agar (1) and incubated at 37°C for 2 days to examine melanin production. This qualitative assay did not reveal any obvious differences between the mutant strains and the wild type (Fig. 3A). Nevertheless, a quantitative analysis of laccase production inL-DOPA medium (19) showed that the
laccase activities of the mutants were increased relative to that of the wild-type strain (Fig. 3B). When 10 mM exogenous cAMP was added, all strains exhibited laccase activities that were indistinguishable from each other (Fig. 3B). Thelac1⌬
control strain, in which theLAC1laccase gene has been elim-inated, exhibited essentially no laccase activity in the presence or absence of exogenous cAMP.
To examine the capsule, the wild-type and mutant strains were inoculated onto DMEM (18) and incubated at 37°C for 2 days. By visual observation and quantitative analysis, capsule production by the mutant strains was modestly enhanced rel-ative to that of the wild type (Fig. 3C and D).
The mating ability of thepde1⌬andpde2⌬mutants was also examined, but no apparent mutant phenotype was observed (data not shown).
Thepde1⌬mutation restores virulence trait production in a
gpa1⌬mutant.We next examined if, like Pde1 and Pde2 inS.
cerevisiae, theC. neoformansPde1 and/or Pde2 protein func-tions in the cAMP-dependent PKA signaling pathway (re-viewed in reference 12). Gpa1 is the G␣subunit that functions early in this pathway, and deletion ofGPA1compromises
mel-FIG. 2.C. neoformansPde1 and Pde2 exhibit signature sequences of low- and high-affinity cAMP PDEases. MacVector software (Accelrys) was used to perform Clustal W comparisons of Pde1 (A) and Pde2 (B). Both proteins contain the conserved signature sequences found in low- and high-affinity homologs, respectively, fromS. cerevisiae,S. pombe, andC. albicans. The Pde1 signature sequence is H-x-H-L-D-H-(LIVM)-x-(GS)-(LIVMA)-(LIVM)2-x-S-(AP), and the Pde2 signature sequence is H-D-(LIVMFY)-x-H-x-(AG)-x2-(NQ)-x-(LIVMFY). Only the signature
se-quences are shown, and the numbers indicate the positions of the sese-quences in the respective proteins. Accession numbers for the proteins are as follows: CnPde1A, AY864841; CnPde1⌬, AY864842; ScPde1, CAA64139; SpPde1, CAA20848; CaPde1, XP720545; CnPde2A, AY874131; CnPde2⌬, AY874132; ScPde2, CAA99689; CaPde2, AAM89252.
on September 8, 2020 by guest
http://ec.asm.org/
anin and capsule production and reduces mating (1, 3). Addi-tionally,gpa1⌬mutants have low basal cAMP levels relative to wild-type strains. We hypothesized that if Pde1 and/or Pde2 functions downstream of Gpa1, then in epistasis analysisgpa1⌬
mutant strains also containing either thepde1⌬orpde2⌬ mu-tation should have higher levels of cAMP than thegpa1⌬single mutant, and these elevated cAMP levels might suffice to sup-pressgpa1⌬mutant phenotypes. This hypothesis was based on the assumption that deletion of PDE1 and/or PDE2 would result in a loss of PDEase activity and thereby reduce the degradation of even the basal levels of cAMP present in the
gpa1⌬strain.
To perform epistasis analysis, pde1⌬ gpa1⌬, pde2⌬gpa1⌬, and pde1⌬ pde2⌬ gpa1⌬ mutant strains were generated via meiotic crosses in a serotype A background. These strains were then inoculated onto Niger seed agar and DMEM to examine melanin and capsule production, respectively. Additionally, each mutant strain was mated with a wild-type strain, and relative mating abilities were determined by visualizing fila-ment production. Melanin production by the gpa1⌬ mutant was fully rescued by thepde1⌬mutation, but not by thepde2⌬
mutation (Fig. 4A), and mating ability was also partially re-stored by thepde1⌬but not thepde2⌬mutation (Fig. 4D). The capsule production ofgpa1⌬mutant cells was clearly rescued by thepde1⌬mutant and may have been modestly rescued by thepde2⌬mutation (Fig. 4B and C). The wild-typePDE1gene complemented thepde1⌬mutation when apde1⌬gpa1⌬strain was transformed with the wild-typePDE1gene. The resulting complemented strain (pde1⌬gpa1⌬PDE1) exhibited melanin, capsule, and mating defects similar to those of thegpa1⌬single mutant (Fig. 4A, C, and D).
To determine if the observed phenotypes of the pde1⌬ gpa1⌬andpde2⌬gpa1⌬double mutant strains and thepde1⌬ pde2⌬gpa1⌬triple mutant strain are correlated with intracel-lular cAMP levels, cAMP assays were performed on samples collected after glucose induction. As previously reported (2, 3), wild-type cAMP levels increased rapidly following the addition of glucose to glucose-starved cells, andgpa1⌬mutants failed to mount a cAMP increase in response to glucose (Fig. 4E). A similar increase was also observed for thepde1⌬ strain, and although the error in the measurements was large, there was a trend towards higher cAMP levels than those in the wild type. Consistent with the results of epistasis analysis, the pde1⌬
mutation enhanced cAMP levels in thepde1⌬gpa1⌬mutant, and glucose clearly enhanced cAMP production in thepde1⌬ pde2⌬gpa1⌬mutant strain (Fig. 4E).
FIG. 3. Deletion of thePDE1andPDE2genes causes modest in-creases in melanin and capsule production. (A) Serotype A wild-type (H99) and pde1⌬ (JKH63), pde2⌬ (JKH33), and pde1⌬ pde2⌬ (JKH70) mutant strains were grown on Niger seed agar, with or with-out 10 mM cAMP, and incubated for 2 days at 37°C prior to being photographed. Melanin production was not obviously different in any of the mutant strains relative to the wild-type strain. (B) Melanin production in serotype A strains was quantitated by growing wild-type (H99) andpde1⌬(JKH63),pde2⌬(JKH33),pde1⌬pde2⌬(JKH70), andlac1⌬(CHM3) mutant strains at 250 rpm and 30°C for 16 h in
L-DOPA medium, with or without 10 mM cAMP, prior to transferring cultures to 250 rpm and 25°C for 24 h. Spectrophotometric readings of the culture supernatants were taken to determine the OD475. One unit
of laccase is equal to an OD475of 0.001 (50). Each bar represents an
average of five independent replicates. Standard deviations are indi-cated by error bars. (C) Serotype A wild-type (H99) and pde1⌬ (JKH63),pde2⌬(JKH33), andpde1⌬pde2⌬(JKH70) mutant strains were inoculated onto agar-based DMEM (18), with or without 10 mM cAMP, incubated at 30°C for 2 days, and examined by the exclusion of India ink (magnification,⫻1,000). (D) Quantitative measurements of relative capsule diameters (see Materials and Methods). A total of 60 cells (20 cells from three independent experiments) were measured for each strain, and error bars indicate standard deviations.
on September 8, 2020 by guest
http://ec.asm.org/
on September 8, 2020 by guest
http://ec.asm.org/
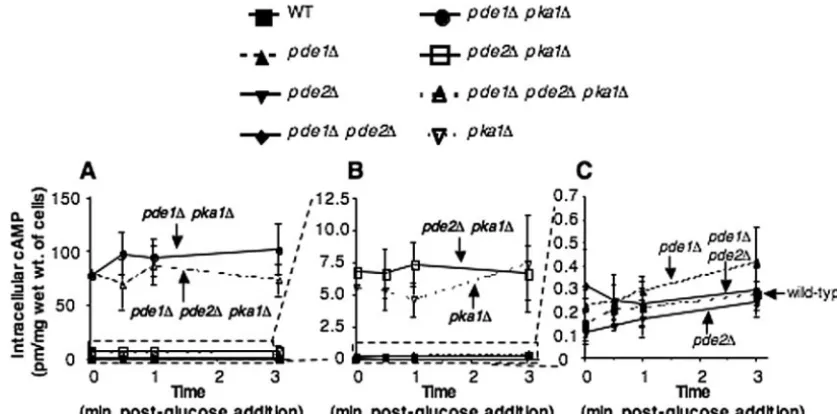
Intracellular cAMP levels are elevated in a pde1⌬
back-ground. InS. cerevisiae, the PKA catalytic subunits regulate
cAMP levels through a negative feedback loop in which Pde1 is activated by PKA phosphorylation (30). Pka1, theC. neofor-mansPKA catalytic subunit, also negatively regulates cAMP levels, but the regulatory mechanism was not known (13). We tested whether this negative feedback loop involves a positive regulation of Pde1 by Pka1.
Previous work has shown thatpka1⌬mutants have elevated intracellular cAMP levels relative to a wild-type strain (13). One plausible model is that activated Pka1 normally activates Pde1, resulting in cAMP degradation, whereas thepka1⌬ mu-tant is unable to activate Pde1. It is also possible that Pka1 has other targets in addition to Pde1 as part of the regulation it imposes on cAMP levels (35). Also, Pde1 may be subject to both Pka1-dependent and -independent regulation (8), allow-ing for a minimal degradation of cAMP even in the absence of a functional Pka1. In either scenario, pde1⌬ pka1⌬ mutants may have elevated cAMP levels relative to thepka1⌬mutant. To test this, intracellular cAMP levels were measured during glucose sensing (Fig. 5).
The wild-type strain and thepde1⌬,pde2⌬, andpde1⌬pde2⌬
mutant strains all showed similarly elevated cAMP levels in response to glucose (Fig. 5C). Thepka1⌬mutant had cAMP levels that were significantly higher (⬃50-fold) than those in the wild-type strain and thepde1⌬,pde2⌬, andpde1⌬pde2⌬
mutant strains (Fig. 5B), as expected based on its established role in a negative feedback loop (13, 46). The pde1⌬ pka1⌬
mutant exhibited cAMP levels that were approximately 10 times higher than those found in the pka1⌬ single mutant strain and⬃500-fold higher than those in the wild-type strain (Fig. 5A), indicating that Pde1 plays a significant role in con-straining cAMP levels, even in the absence of Pka1. These data also suggest that Pka1 has additional targets besides Pde1, because the phenotype conferred by thepka1⌬mutation on cAMP levels is exerted even in the absence of Pde1 (compare thepde1⌬mutant and thepde1⌬pka1⌬mutant in Fig. 5). Also, these data provide further support that Pde2 plays only a minor role, if any, in regulating cAMP levels.
Elimination of a PKA phosphorylation site causes a loss of
Pde1 function.TheS. cerevisiaePde1 enzyme has one
consen-sus PKA phosphorylation site [(K/R)-(K/R)-(X)-(S/T)] (26). Mutagenesis of the serine residue in this site (pde1S252A)
re-sults in reduced Pde1 activity (30). To determine if Pka1
phos-FIG. 5.pde1⌬mutation results in highly elevated intracellular cAMP levels in apka1⌬background. Intracellular cAMP levels were measured in a serotype A wild-type (H99) strain and thepka1⌬(JKH7),pde1⌬pka1⌬(JKH95),pde2⌬pka1⌬(JKH94),pde1⌬pde2⌬pka1⌬(JKH109),pde1⌬ (JKH63),pde2⌬(JKH33), andpde1⌬pde2⌬(JKH70) mutant strains. Thepde1⌬pka1⌬andpde1⌬pde2⌬pka1⌬strains produced significantly higher levels of cAMP than any of the other strains, and the scale for the data shown in panel A was expanded to illustrate this (B). Thepka1⌬ andpde2⌬pka1⌬strains had levels higher than those of the other strains, and the scale for the data in panel B was expanded to show this (C).
FIG. 4.pde1⌬mutation restores mating and melanin and capsule production of gpa1⌬ mutants. (A) Serotype A wild-type (H99),gpa1⌬ (YSB83),pde1⌬gpa1⌬(JKH111),pde2⌬gpa1⌬(JKH101),pde1⌬pde2⌬gpa1⌬(JKH116),pde1⌬(JKH63),pde2⌬(JKH33), andpde1⌬pde2⌬ (JKH70) mutant, andpde1⌬gpa1⌬PDE1complemented strains were grown on Niger seed agar and incubated for 2 days at 37°C prior to being photographed. (B) The strains used for panel A were grown in YPD overnight prior to being plated onto agar-based DMEM, incubated for 2 days at 37°C, resuspended in ddH2O, stained with India ink, and photographed at a magnification of⫻1,000. (C) For each strain used for panels A and
B, a total of 60 cells (20 cells from three independent experiments) were treated as described in panel B and then measured (see Materials and Methods). Error bars indicate standard deviations. (D)MAT␣andMATastrains were cocultured on V8 (pH 5.0) medium and incubated at room temperature for 14 days in the dark prior to being photographed (magnification,⫻100). These strains included␣ ⫻a(H99⫻KN99a),gpa1⌬ ⫻
a(YSB83⫻KN99a),pde1⌬gpa1⌬ ⫻a(JKH111⫻KN99a),pde2⌬gpa1⌬ ⫻a(JKH101⫻H99),pde1⌬pde2⌬gpa1⌬ ⫻a(JKH116⫻H99), and pde1⌬gpa1⌬PDE1⫻a(JKH190⫻H99). (E) Intracellular cAMP levels were measured in each of the strains used for panels A to C.
on September 8, 2020 by guest
http://ec.asm.org/
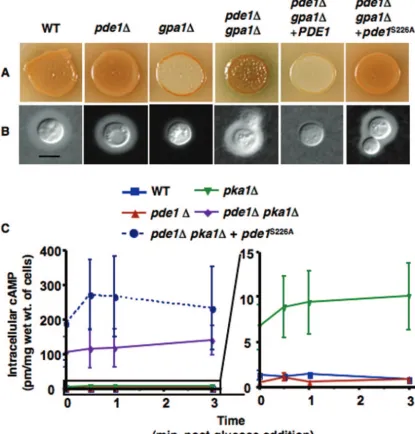
phorylates and subsequently activates Pde1 inC. neoformans, we examined the Pde1 sequence for putative PKA phosphor-ylation sites, using the PROSCAN site/signature sequence rec-ognition program, and found that the serotype A Pde1 protein contains one putative PKA phosphorylation site (RKKS). We predicted that if Pka1 activates Pde1 by phosphorylation of the serine residue in this site, then mutagenesis of the serine to an alanine would result in a Pde1 protein that was unable to be activated and would hence act as apde1⌬allele.
To test this hypothesis, we generated aPDE1allele identical to the construct that was utilized to complement thepde1⌬
mutation in agpa1⌬background, except for an S226A muta-tion at the serine residue in the putative PKA phosphorylamuta-tion site. Both thepde1⌬gpa1⌬strain and thepde1⌬pka1⌬strain were transformed with thepde1S226Aallele. Northern analysis
indicated that thepde1S226Atranscript level was comparable to
that of the wild-type PDE1gene, although this does not ex-clude the possibility that the resulting phenotypes could be attributable to a loss of function or instability of thepde1S226A
protein. Thepde1⌬gpa1⌬pde1S226Atransformant was
exam-ined for the ability to produce melanin and capsule. As pre-dicted if thepde1S226Aallele acts as apde1null mutant,
mel-anin and capsule production in the pde1⌬ gpa1⌬ pde1S226A
strain was indistinguishable from that in thepde1⌬gpa1⌬ mu-tant (Fig. 6A and B). This is in contrast to thepde1⌬ gpa1⌬ PDE1-complemented strain, in which the melanin and capsule defects conferred by thegpa1⌬mutation were readily appar-ent. When thepde1S226Aallele was placed in apde1⌬pka1⌬
background, cAMP levels were elevated similarly to the levels found in apde1⌬pka1⌬mutant (Fig. 6C), again suggesting that
thepde1S226Aallele acts like apde1⌬null mutant.
DISCUSSION
Cyclic AMP signaling plays multiple roles in regulating de-velopment in eukaryotic organisms as diverse as slime molds, fungi, and humans. Here we define thePDE1andPDE2genes in the basidiomycete fungusC. neoformansand their biological function in cAMP signaling. Although similar to proteins in the divergent fungal modelS. cerevisiaein some aspects, both the Pde1 and Pde2 proteins also show functional distinctions from their S. cerevisiae counterparts. The PDE1 gene encodes a phosphodiesterase protein that has a significant role in regu-lating cAMP levels in the cell. ForS. cerevisiae, it has been determined that Pde1 is responsible for regulating agonist-induced cAMP levels, based on cAMP assays showing that the initial, transient spike in cAMP levels that occurs after glucose addition was enhanced in a pde1⌬ mutant (30). In the C. neoformans pde1⌬ mutant, only a modest enhancement of cAMP levels was observed upon glucose induction. Pde1 may be regulated through other pathways in addition to the cAMP-dependent signaling pathway (Fig. 1) (8), a hypothesis sup-ported by our observation that in a pde1⌬ pka1⌬ mutant, cAMP levels are much higher than in apka1⌬single mutant, suggesting that Pka1-independent regulation of Pde1 may oc-cur such that in apka1⌬mutant there is still some activity of Pde1 allowing for cAMP degradation. Also, it is possible that the feedback inhibition of cAMP levels initiated by Pka1 (see below) is robust enough that cAMP levels do not escalate substantially when Pde1 is absent.
Deletion of thePDE1gene by itself conferred only modest increases in melanin and capsule production in otherwise wild-type cells. However, deletion of PDE1 in combination with mutation of the G␣ subunit Gpa1 remediated the melanin, capsule, and mating defects caused by thegpa1⌬single muta-tion. cAMP assays showed significantly higher cAMP levels in apde1⌬gpa1⌬double mutant than in agpa1⌬single mutant, supporting a model in which the deletion ofPDE1in agpa1⌬
background results in cAMP levels that are sufficient to sub-stantially activate the protein kinase catalytic subunit Pka1. The observation that thepde1⌬ pde2⌬ gpa1⌬ mutant strain responded to glucose in the absence of a functional G␣subunit suggests that this response is independent of the G-protein-coupled receptor. As in S. cerevisiae, it is possible that C. neoformansis able to sense intracellular glucose levels, possibly as the metabolite glucose-6-phosphate (40), but that intracel-lular glucose levels stimulate cAMP to such a low extent that only in the absence of one or both phosphodiesterases is the increase in cAMP levels detectable.
Previous studies have shown that inC. neoformans, activated Pka1 mediates a negative feedback loop that controls intracel-lular cAMP levels (13, 46). However, the mechanism for this regulation had not been elucidated. S. pombe also exhibits negative feedback regulation of cAMP levels, and it has been established that Pde1 is part of this regulatory network, al-though the relationship between Pde1 and the PKA catalytic subunit Pka1 is not clear (21). In contrast, forS. cerevisiaeit has been definitively shown that the PKA catalytic subunits phosphorylate serine residue 252 in Pde1, causing the activa-tion and subsequent degradaactiva-tion of cAMP. Site-directed mu-tagenesis of this residue to create apde1S252Aallele resulted in
decreased Pde1 activity in vitro (30). Our data suggest that a similar situation to that inS. cerevisiaeexists inC. neoformans. A single putative PKA phosphorylation site was found in Pde1, at residues 223 to 226. A site-directed mutation (pde1S226A) in
the serine residue of this putative phosphorylation site reduced the protein activity of Pde1. Additionally, both the pde1⌬ pka1⌬ mutant and the pde1⌬ pka1⌬ pde1S226A mutant had
cAMP levels that were elevated relative to that in thepka1⌬
single mutant. This suggests that Pde1 is activated indepen-dently of Pka1, in addition to Pka1-dependent regulation, and that Pka1 targets other proteins in addition to Pde1 as part of the negative feedback loop that modulates intracellular cAMP levels (Fig. 1).
Additional candidate targets of Pka1 include proteins up-stream of Pka1 in the cAMP-dependent signaling pathway, such as the Pkr1 protein kinase A regulatory subunit and the Cac1 adenylyl cyclase protein homologs from divergentC. neo-formansserotypes, which both have putative conserved PKA phosphorylation sites. InS. cerevisiae, the Bcy1 PKA regulatory subunit is not thought to be a candidate for additional feed-back regulation, while the Cyr1 adenylyl cyclase is (43). InS. pombe, cAMP levels are controlled by the regulation of both adenylyl cyclase and the Pde1 homolog, Cgs2 (21). Studies with
S. pombesuggest that shortly after the glucose-stimulated ac-tivation of adenylyl cyclase, the Cgs2 phosphodiesterase is also activated, resulting in a modulation of cAMP levels (45). Site-directed mutagenesis of the sites in theC. neoformans Cac1 and Pkr1 proteins should elucidate whether these candidate proteins are targets of Pka1 and if they have roles in the
on September 8, 2020 by guest
http://ec.asm.org/
negative feedback loop that regulates cAMP levels inC. neo-formans.
The role of Pde2 in regulating cAMP (30) production is less obvious than that of Pde1. Pde2 contains the class I signature sequence found in numerous high-affinity PDEase proteins, and RNA analysis confirmed that a transcript was produced. However, deletion of thePDE2gene has only modest effects on mating or melanin or capsule production, does not result in a significant change from the wild-type strain with regard to
cAMP levels, and results in no change in heat shock or oxida-tive stress resistance. Pde2 regulates heat shock sensitivity inS. cerevisiae(30); that it does not inC. neoformanssupports other evidence that the cAMP signaling pathway is not as involved in the response to environmental stresses as it is in other fungi (24). We noted that the pde2⌬ mutation appears to weakly rescue capsule production in agpa1⌬strain (Fig. 4A) and thus might function under some more limited physiological con-texts, in contrast to Pde1. These data suggest that Pde2 either
FIG. 6. Elimination of a PKA phosphorylation site results in a loss of Pde1 activity and increased cAMP levels in apka1⌬background. Site-directed mutagenesis was utilized to eliminate the single PKA phosphorylation site in Pde1 (pde1S226A). The mutantPDE1alleles were
introduced into thepde1⌬gpa1⌬background, and the resultantpde1⌬gpa1⌬pde1S226A(JKH238) strain was compared with the wild-type strain
(H99) and thegpa1⌬(YSB83),pde1⌬(JKH63),pde1⌬gpa1⌬(JKH173), andpde1⌬gpa1⌬PDE1(JKH190) mutant strains. Melanin (A) and capsule (B) production were examined as described in Materials and Methods. Cultures for examinations of capsule production were incubated at 37°C for 2 days prior to being observed by India ink staining. Bar, 10m. (C) cAMP levels were also examined as described in Materials and Methods. Thepde1⌬pka1⌬andpde1⌬pka1⌬pde1S226Astrains produced significantly higher levels of cAMP than any of the other strains, and the
pka1⌬strain also had levels higher than those of the wild type or thepde1⌬mutant; the scale for the data shown in the graph on the left was expanded to illustrate this.
on September 8, 2020 by guest
http://ec.asm.org/
is not in the same signaling pathway as Gpa1, does not function similarly to Pde1, or is not regulated in the same manner as Pde1.
It is probable that C. neoformanscontains two related but distinct feedback regulation systems controlling the cAMP pathway, one involving cAMP degradation (via induction of the Pde1 phosphodiesterase) and another involving cAMP production (via the repression of adenylyl cyclase). This dual regulation suggests that precise regulation of the cAMP path-way may be critical to the maintenance of virulence attributes of the pathogen. The fact that several other fungi, including the nonpathogenic organismsS. cerevisiaeandS. pombe, share this regulatory network supports the contention that the constraint of intracellular cAMP levels is critical for cellular function in many organisms. Therefore, future studies ofC. neoformans
will investigate the impact of this strict constraint on the cAMP pathway in fungal survival and virulence during the course of disease.
ACKNOWLEDGMENTS
We thank Xiaorong Lin, Jeffrey Batten, Chaoyang Xue, and Alex Idnurm for helpful suggestions in the preparation of the manuscript. This research was funded in part by an NIH interdisciplinary AIDS training grant (AI07392-14) (J.K.H.) and by RO1 grant AI39115 and PO1 grant AI44975 (J.H.).
REFERENCES
1.Alspaugh, J. A., J. R. Perfect, and J. Heitman.1997.Cryptococcus neofor-mansmating and virulence are regulated by the G-protein␣subunit GPA1 and cAMP. Genes Dev.11:3206–3217.
2.Alspaugh, J. A., R. Pukkila-Worley, T. Harashima, L. M. Cavallo, D. Fun-nell, G. M. Cox, J. R. Perfect, J. W. Kronstad, and J. Heitman.2002. Adenylyl cyclase functions downstream of the G␣protein Gpa1 and controls mating and pathogenicity ofCryptococcus neoformans. Eukaryot. Cell1:75– 84.
3.Bahn, Y. S., J. K. Hicks, S. S. Giles, G. M. Cox, and J. Heitman.2004. Adenylyl cyclase-associated protein Aca1 regulates virulence and differenti-ation ofCryptococcus neoformansvia the cyclic AMP-protein kinase A cas-cade. Eukaryot. Cell3:1476–1491.
4.Bahn, Y. S., K. Kojima, G. M. Cox, and J. Heitman.2005. Specialization of the HOG pathway and its impact on differentiation and virulence of Cryp-tococcus neoformans. Mol. Biol. Cell16:2285–2300.
5.Bahn, Y. S., J. Staab, and P. Sundstrom.2003. Increased high-affinity phos-phodiesterasePDE2gene expression in germ tubes counteractsCAP1- de-pendent synthesis of cyclic AMP, limits hypha production and promotes virulence ofCandida albicans. Mol. Microbiol.50:391–409.
6.Casadevall, A., and J. R. Perfect.1998.Cryptococcus neoformans. ASM Press, Washington, D.C.
7.Charbonneau, H., N. Beier, K. A. Walsh, and J. A. Beavo.1986. Identifica-tion of a conserved domain among cyclic nucleotide phosphodiesterases from diverse species. Proc. Natl. Acad. Sci. USA83:9308–9312.
8.Conti, M.2000. Phosphodiesterases and cyclic nucleotide signaling in endo-crine cells. Mol. Endocrinol.14:1317–1327.
9.D’Angelo, M. A., S. Sanguineti, J. M. Reece, L. Birnbaumer, H. N. Torres, and M. M. Flawia.2004. Identification, characterization and subcellular localization of TcPDE1, a novel cAMP-specific phosphodiesterase from
Trypanosoma cruzi. Biochem. J.378:63–72.
10.Davidson, R. C., J. R. Blankenship, P. R. Kraus, M. D. Berrios, C. M. Hull, C. D’Souza, P. Wang, and J. Heitman.2002. A PCR-based strategy to generate integrative targeting alleles with large regions of homology. Micro-biology148:2607–2615.
11.De Voti, J., G. Seydoux, D. Beach, and M. McLeod.1991. Interaction be-tweenran1⫹protein kinase and cAMP dependent protein kinase as negative regulators of fission yeast meiosis. EMBO J.10:3759.
12.D’Souza, C., and J. Heitman.2001. Conserved cAMP signaling cascades regulate fungal development and virulence. FEMS Microbiol. Rev.25:349– 364.
13.D’Souza, C. A., J. A. Alspaugh, C. Yue, T. Harashima, G. M. Cox, J. R. Perfect, and J. Heitman.2001. Cyclic AMP-dependent protein kinase con-trols virulence of the fungal pathogenCryptococcus neoformans. Mol. Cell. Biol.21:3179–3191.
14.Dunlap, P. V., and S. M. Callahan.1993. Characterization of a periplasmic 3⬘:5⬘-cyclic nucleotide phosphodiesterase gene,cdpP, from the marine sym-biotic bacteriumVibrio fischeri. J. Bacteriol.175:4615–4624.
15.Fraser, J. A., R. L. Subaran, C. B. Nichols, and J. Heitman.2003. Recapit-ulation of the sexual cycle of the primary fungal pathogenCryptococcus neoformansvar.gattii: implications for an outbreak on Vancouver Island. Eukaryot. Cell2:1036–1045.
16.Gao, S., and D. L. Nuss.1996. Distinct roles for two G protein␣subunits in fungal virulence, morphology, and reproduction revealed by targeted gene disruption. Proc. Natl. Acad. Sci. USA93:14122–14127.
17.Gold, S., G. Duncan, K. Barrett, and J. Kronstad.1994. cAMP regulates morphogenesis in the fungal pathogenUstilago maydis. Genes Dev.8:2805– 2816.
18.Granger, D. L., J. R. Perfect, and D. T. Durack.1985. Virulence of Crypto-coccus neoformans:regulation of capsule synthesis by carbon dioxide. J. Clin. Investig.76:508–516.
19.Hicks, J. K., C. A. D’Souza, G. M. Cox, and J. Heitman.2004. Cyclic-AMP dependent protein kinase catalytic subunits have divergent roles in virulence factor production in two varieties of the fungal pathogenCryptococcus neo-formans. Eukaryot. Cell3:14–26.
20.Hicks, J. K., J. Yu, N. P. Keller, and T. H. Adams.1997.Aspergillus sporu-lation and mycotoxin production both require inactivation of the FadA G-protein-dependent signaling pathway. EMBO J.16:4916–4923. 21.Hoffman, C. S.2005. Glucose sensing via the protein kinase A pathway in
Schizosaccharomyces pombe. Biochem. Soc. Trans.33:261–264.
22.Hoyer, L. L., L. B. Cieslinski, M. M. McLaughlin, T. J. Torphy, A. R. Shatzman, and G. P. Livi.1994. ACandida albicanscyclic nucleotide phos-phodiesterase: cloning and expression inSaccharomyces cerevisiaeand bio-chemical characterization of the recombinant enzyme. Microbiology140: 1533–1542.
23.Hull, C. M., and J. Heitman.2002. Genetics ofCryptococcus neoformans. Annu. Rev. Genet.36:557–615.
24.Idnurm, A., Y.-S. Bahn, K. Nielsen, X. Lin, J. Fraser, and J. Heitman.2005. Deciphering the model pathogenic fungusCryptococcus neoformans. Nat. Rev. Microbiol.3:753–764.
25.Jung, W. H., P. Warn, E. Ragni, L. Popolo, C. D. Nunn, M. P. Turner, and L. Stateva. 2005. Deletion ofPDE2, the gene encoding the high-affinity cAMP phosphodiesterase, results in changes of the cell wall and membrane inCandida albicans. Yeast22:285–294.
26.Kemp, B. E., and R. B. Pearson.1990. Protein kinase recognition sequence motifs. Trends Biochem. Sci.15:342–346.
27.Kunz, S., T. Kloeckner, L.-O. Essen, T. Seebeck, and M. Boshart.2004. TbPDE1, a novel class I phosphodiesterase ofTrypanosoma brucei. Eur. J. Biochem.271:637–647.
28.Lacombe, M.-L., G. J. Podgorski, J. Franke, and R. H. Kessin.1986. Molecular cloning and developmental expression of the cyclic nucleotide phosphodiester-ase gene ofDictyostelium discoideum. J. Biol. Chem.261:16811–16817. 29.Lengeler, K. B., R. C. Davidson, C. D’Souza, T. Harashima, W.-C. Sheng, P.
Wang, X. Pan, M. Waugh, and J. Heitman.2000. Signal transduction cas-cades regulating fungal development and virulence. Microbiol. Mol. Biol. Rev.64:746–785.
30.Ma, P., S. Wera, P. V. Dijck, and J. M. Thevelein.1999. ThePDE1-encoded low-affinity phosphodiesterase in the yeastSaccharomyces cerevisiaehas a specific function in controlling agonist-induced cAMP signaling. Mol. Biol. Cell10:91–104.
31.Meima, M. E., K. E. Weening, and P. Schaap.2003. Characterization of a cAMP-stimulated cAMP phosphodiesterase in Dictyostelium discoideum. J. Biol. Chem.278:14356–14362.
32.Mitchell, T. K., and R. A. Dean.1995. The cAMP-dependent protein kinase catalytic subunit is required for appressorium formation and pathogenesis by the rice blast pathogenMagnaporthe grisea. Plant Cell7:1869–1878. 33.Nelson, R. T., J. Hua, B. Pryor, and J. K. Lodge.2001. Identification of
virulence mutants of the fungal pathogenCryptococcus neoformansusing signature-tagged mutagenesis. Genetics157:935–947.
34.Nikawa, J., P. Sass, and M. Wigler.1987. Cloning and characterization of the low affinity cAMP phosphodiesterase gene ofS. cerevisiae. Mol. Cell. Biol. 7:3629–3636.
35.Nikawa, J.-I., S. Cameron, T. Toda, K. M. Ferguson, and M. Wigler.1987. Rigorous feedback control of cAMP levels in Saccharomyces cerevisiae. Genes Dev.1:931–937.
36.Pan, X., and J. Heitman.1999. Cyclic AMP-dependent protein kinase reg-ulates pseudohyphal differentiation inSaccharomyces cerevisiae. Mol. Cell. Biol.19:4874–4887.
37.Park, J. I., C. M. Grant, and I. W. Dawes.2005. The high-affinity cAMP phosphodiesterase ofSaccharomyces cerevisiaeis the major determinant of cAMP levels in stationary phase: involvement of different branches of the Ras-cyclic AMP pathway in stress responses. Biochem. Biophys. Res. Com-mun.327:311–319.
38.Perfect, J. R., N. Ketabchi, G. M. Cox, C. W. Ingram, and C. L. Beiser.1993. Karyotyping ofCryptococcus neoformansas an epidemiological tool. J. Clin. Microbiol.31:3305–3309.
39.Rascon, A., M. E. Viloria, L. De-Chiara, and M. E. Dubra.2000. Character-ization of cyclic AMP phosphodiesterases inLeishmania mexicanaand pu-rification of soluble form. Mol. Biochem. Parasitol.106:283–292.
on September 8, 2020 by guest
http://ec.asm.org/
40.Rolland, F., J. Winderickx, and J. M. Thevelein.2002. Glucose-sensing and signalling mechanisms in yeast. FEMS Yeast Res.2:183–201.
41.Sherman, F.1991. Getting started with yeast. Methods Enzymol.194:3–21. 42.Sonneborn, A., D. P. Bockmuhl, M. Gerads, K. Kurpanek, D. Sanglard, and J. F. Ernst.2000. Protein kinase A encoded byTPK2regulates dimorphism ofCandida albicans. Mol. Microbiol.35:386–396.
43.Thevelein, J. M., and J. H. D. Winde.1999. Novel sensing mechanisms and targets for the cAMP-protein kinase A pathway in the yeastSaccharomyces cerevisiae. Mol. Microbiol.33:904–918.
44.Toffaletti, D. L., T. H. Rude, S. A. Johnston, D. T. Durack, and J. R. Perfect. 1993. Gene transfer inCryptococcus neoformansby use of biolistic delivery of DNA. J. Bacteriol.175:1405–1411.
45.Wang, L., K. Griffiths, Jr., Y. H. Zhang, F. D. Ivey, and C. S. Hoffman.2005.
Schizosaccharomyces pombeadenylate cyclase suppressor mutations suggest a role for cAMP phosphodiesterase in feedback control of glucose/cAMP signaling. Genetics 10.1534/genetics.105.047233.
46.Wang, P., G. M. Cox, and J. Heitman.2004. A Sch9 protein kinase homo-logue controlling virulence independently of the cAMP pathway in Crypto-coccus neoformans. Curr. Genet.46:247–255.
47.Wang, P., C. B. Nichols, K. B. Lengeler, M. E. Cardenas, G. M. Cox, J. R. Perfect, and J. Heitman.2002. Mating-type-specific and nonspecific PAK kinases play shared and divergent roles inCryptococcus neoformans. Eu-karyot. Cell1:257–272.
48.Yu, J.-H., J. Wieser, and T. H. Adams.1996. TheAspergillusFlbA RGS domain protein antagonizes G protein signaling to block proliferation and allow development. EMBO J.15:5184–5190.
49.Zaragoza, O., B. C. Fries, and A. Casadevall.2003. Induction of capsule growth inCryptococcus neoformansby mammalian serum and CO2. Infect. Immun.71:6155–6164.
50.Zhu, X., J. Gibbons, J. Garcia-Rivera, J. Casadevall, and P. R. Williamson. 2001. Laccase ofCryptococcus neoformansis a cell wall-associated virulence factor. Infect. Immun.69:5589–5596.