Indian J. Plant PhYliol •• Vol. XXIX, No.4, pp. 351-356 (December. 1986)
EFFECT OF SOURCE MANIPULATION ON PHOTOSYNTHETIC. ACTIVITY IN GROUNDNUT
B. K. GHOSH· AND U. K. SENGUPTA Division ofPlant Physiology,
Indian Agricultural Research Institute, New Delhi-110 012
SUMMARY
Photosynthetic activity of groundnut leaves located at dift'erent branches during peak pod developmental phase was studied. The dift'e rences in photosynthetic rate of the leaves situated on the main shoot and three basal branches were almost negligible. The leaves of uppermost branch (5th) aJthoulh received greater solar radiation showed lowest photosynthetic activity. Partial defoliation or covering of leaves en hanced the photosynthetic rate which was more in the basal branches compared to main shoot. This enhancement in photosynthetic rates was more due to covering than defoliation.
INTRODUCTION
Investigations on the source-sink relationship through manipulation of source size have shown that partial defoliation enhanced carbon dioxide ex change rate of intact leaves (Carmi and Kollar, 1979; Hanson and West, 1982) and dependent on the relative position of the leaves to the developing fruits (Boote et 01., 1980). In ground nut, defoliation to different degrees and at different stages of growth, decreased the stem and pod growth rates (Williams et 01., 1976; Boote et 01., 1980). During early weeks of defoliation reduction in CO2 exchange rate occurred, but was followed by partial recovery at a later stage (Jones et 01., 1982). However, no efforts were made to identify tho importance of leaves in relation to position of branches and their relation to growth of plant.
Defoliation treatment imposes a shock-effect on the plant and recovery mechanism operates followed by the development of compensatory leaves. Another way, by which photosynthetic surface could be reduced is by covering the leaves without causing injury to the plants. RepOPts regarding comparative studies on defoliation and covering are rather meagre. The present study reports how reduction of leaf surface artificially at one position affect the - photosynthetic activity on the remaining part of the groundnut plant.
352 B. K. GHOSH AND U. K. SENGUPTA
MATERIALS AND METHODS
Groundnut cuitivar J-l1 (erect type) was raised in pot cultures under conditions of optimum nutrient supply and water. Photosynthetic surface was reduced through (i) defoliation and (ij) leaf covering. The later treatment was employed to analyse the impact of leaf removal on the photosynthetic rate.
The above treatments were started from initial flowering stage 30 DAS (days after sowing) and photosynthetic rates were measured at 85 DAS. For defoliation treatment, leaves of individual branches were removed, while for covering treatment, the whole branch was covered with thin black polythene sheets having small holes at the lower side for aeration. The treatments were given either on single branch or in combination with other which are as follows: Defoliation or covering of : 0) all branches except main shoot (ii) all branches except main shoot, branch I and 2 (iii) all branches except main shoot, branch 3 and 4 (iv) main shoot only (v) all branches except branch 1 and 2 (vi) none (control).
Photosynthetic activity of the top fully open leaves was measured using Infra red gas analyser under a light intensity of 800 p.E/m2/sec.
RESULTS AND DISCUSSION
Photosynthetic rate of leaves situated at different branch positions was measured at peak reproductive stage of groundnut (Table 1). At this stago
Table I: Photosynthetic rate of the leaves situated on different branches at peak pod developmental stage in groundnut.
Leaf sample Photosynthetic rate
taken from (mg COI/dmz/hr)
Main shoot 23.62 (100.00)
Branch-l 23.64 (100.00)
Branch-2 24.08 (101.00)
Branch-3
•
23.04 (97.00)Branch-4 22.48 ('5.00)
Branch-S 16.42 (69.50)
j /
PHOTOSYNTHETIC ACTIVITY IN GROUNDNUT 353
10)Vest ~te 9f photo~yntbesis was.recorded in the leaves of uppermost branch (5th) and highest rates were observed in the. maih shoot and lower branches. Leaves of branches I, 2, 3 and main shoot showed almost equal photosynthetic rates. Though. the two basal bra,nches 1 and 2 sho,wed, same photosynthetic rate as the main shoot, their potential is not fully utilized under field conditions as these lower branches remain under the canopy of upper branches and thus receive less sun light relative to upper branches. These two branches are the main ~ontributors of assimilates as maximum J1u~ber of pods are formed in these two branches (Sharma et al., 1981; Sengupta et al., 1985). The leaves of top most branch (5th), which receive maximum solar radiation. showed mini
mum rate of CO2 assimilation. This' branch has also been shown to be least efficient for supporting growth' requirement of groundnut crop (Sharmaital., 1981).
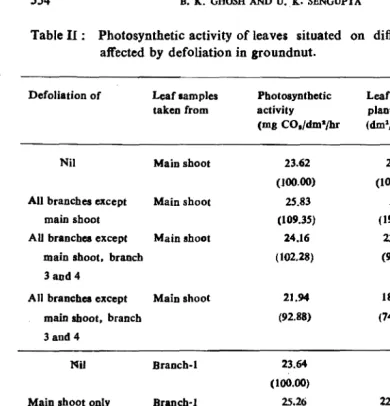
Partial reduction of photosynthetic surface from different branches .caused enhancement of C02 uptake rate in the intact leaves on other branches (Table 11). As most of the pods are produced from the main shoot and bi"anch~l, the changes in photosynthetic rates in the leaves of these two branches are reported (Sengupta et al., 1985). Complete defoliation except the main shoot caused greater increase in the rate of photosynthesis in the leaves of main shoot as compared to the plant with undefoliated main shoot plus two more branches. The enhancement in photosynthetic rate due to defoliation was greater on the intaet leaves of branch-l than on the main shoot. Thus defoliation of only main shoot caused higher effect on the rate of photosynthesis in the branch-I. Highest increase in COl uptake was recorded in the leaves of branch-l when only branches I & 2 were left undefoliated.
Leaves of different branches were covered in order to avoid injury and to compare with defoliation treatment. Covering of the leaves caused much higher rate of photosynthesis on the leaves of remaining part of the plant as compared to defoliation treatment (Table III). When all the branches were eovered except the main shoot, the enhancement effect on the leaves of main shoot was 25% compared to 9% due to defoliation. Similarly, when only branches 1 and 2 were covered, the enhancement was 19% on the main shoot and nil in defoliation treatment.
354
~h
1
B. K. GHOSH AND U. K. SENGUPTA
Table II: Photosynthetic activity of leaves situated on different branches as affected by defoliation in groundnut.
Defoliation of Leaf samples Photosynthetic Leaf area/ Pod yield/
taken from activity plant plant
(rna CO./dmsjhr (dm1/plant) (a/plant)
Nil Main shoot 23.62 25.10 16.30
(100.00) (100.00) (100.00)
All branches except Main shoot 25.83 5.01 8.69
main shoot (109.35) ( 19.90) (53.30)
AU branches except Main shoot 24.16 23.19 1S.33
main shoot, branch (102.28) (92.33) (94.40)
3 and 4
All branches except Main shoot 21.94 18.70 12.42
main shoot, branch (92.88) (74.50) (76.20)
3and4
Nil Branch-I 23.64
(100.00) / '
Main shoot only Branch-J 25.26 22.16 15.32
(106.85) (88.12) (93.90) All branches except Branch-I 24.24
main shoot and branch (192.53) J and 2
All branches exccPt Branch-I 26.12 18.80 12.77
branch 1 and 2 (110.49, (74.90) (78.30)
Values on paranthesis indicate per cent over respective control.
of branch-l when the leaves of all the branches were covered except branches 1 and 2. Covering the branches 3 and 4 had less effect on the photosynthetic rate of branch-I.
The results tbus sbowed. tbat the reduction in leaf area either by defolia tion or covering affected the photosynthetic rates of the remaining leaves (Carmi and Kollar, 1979; Hanson and West, 1982) through the enhancement in photo synthetic rate was not same in tbe leaves of all the branches. The enhancement
,.
/
PHOTOSYNTHETIC ACTIVITY IN GROUNDNUT 355
Table III: Photosynthetic activity of leaves situated on different branches as affected by leaf covering.
Leaf covering of Leaf IIAmpJes Photosynthetic
taken from activity
(mg CO./dma/hr)
Nil
All branches except main shoot
All branches except main shoot, branch 1 " 2
An branches eXcep-t main shoot, branch 3 &; 4
Main shoot Main shoot Main shoot Main shoot
23.62 (100.00)
29.40 (125.64)
24.44
(103.47)
28.32
(119.80)
Nil Branch-! 23.64
Main shoot only Branch-1
(100.00)
30.)0
All bl'anches t;xcept main shoot, branch 1 " 2 All branches except branch 1 "2
Branch-l Branch-!
(127.23)
2S.66 (108.28)
30.80
(130.28)
Values on parenthesis indicate per cent over respective control.
was more in the basal branches where, most of the pods were formed. This indicates that photosynthetic compensation occurs more in the branch-] than other branches due to reduction in leaf surface on other parts of the plant. This effect was observed more due to covering than defoliation treatment which may be traced due to injury and shock effect of defoliation and discontinuity of defoliated leaves from other part of the plant. Under covering treatments, the leaves undergo senescence and some of the metabolites probably move from these leaves to other parts of the plant. As a consequence, there was a higher rate of photosynthesis in the remaining part of the plant.
REFERENCES
356 B. K. GHOSH AND U. K. SENGUPTA
<;anQi. A. and Kollar. D. (]979). Regulation of pbotosyntbetic activity in the primary leaves of bean (Phaleolus vulagril L.) by materials moving in tbe water conducting system. Plant PhYliol •• 64: 28'-288.
Hanlon, W. D. and West. D. R. (1912). Source-sink relationsbips in soybeans. I. Effect of source manipulatian during vegetative growth on drY matter distribution. Crop Sci., 22: 327-376.
Jones, J. W•• Barfield, C. S., Boote. K. J., Scnerage, G. H. and Mangold, J. (1~82). Photo synthetic recoverY of peanut to defoliation at various growth stages. Crop Sci•• 22: 741-746.
Sengupta, U. K. and Sharma, Aruna (1984). Studies on assimilate translocation in relation to yield in groundnut.lndian J. Plant PhYliol., 27: 232-238.
Sbarm~. A•• Reddy, K. J•• Sirohi, G. S. and Sengupta. U. K. (1981). Pattern of partitioning of pbotosyntbates during pod development in groundnut. Indian J. Expt. BifJl•• 19: 2So..2S2.