Copyright © 2000, American Society for Microbiology. All Rights Reserved.
Problems Related to Determination of MICs of Oximino-Type
Expanded-Spectrum Cephems for
Proteus vulgaris
AKIRA OHNO,* YOSHIKAZU ISHII, LING MA,ANDKEIZO YAMAGUCHI Department of Microbiology, Toho University School of Medicine, Ota-ku, Tokyo, Japan
Received 22 March 1999/Returned for modification 1 June 1999/Accepted 29 October 1999
During in vitro susceptibility testing of clinical isolates ofProteus vulgaris, we noted that the MICs of several expanded-spectrum cephems were much higher in the broth microdilution method than in the agar dilution method (termed the MIC gap phenomenon). Here we investigated the mechanism of the MIC gap phenomenon. Cephems with the MIC gap phenomenon were of the oximino type, such as cefotaxime, cefteram, and cefpo-doxime, which serve as good substrates for inducible class A-lactamase (CumA) enzymes produced byP.
vulgaris; this finding suggests a relationship between the MIC gap phenomenon and CumA. Since
peptidogly-can recycling shares a system common to that inducing CumA, we analyzed the mechanism of the MIC gap phenomenon usingP. vulgarisB317 and isogenic mutants with mutations in the peptidoglycan recycling and
-lactamase induction systems. The MIC gap phenomenon was observed in the parent strain B317 but not in B317G (cumG-defective mutant; defective peptidoglycan recycling) and B317R (cumR-defective mutant; defec-tive CumA transcriptional regulator). No-lactamase activity was detected in B317G and B317R.-Lactamase activity and the MIC gap phenomenon were restored in B317G/pMD301 (strain transcomplemented by a clonedcumGgene) and B317R/pMD501 (strain transcomplemented by a clonedcumRgene). MICs determined by the agar dilution method increased when lower agar concentrations were used. Our results indicated that the mechanism of the MIC gap phenomenon is related to peptidoglycan recycling and CumA induction systems. However, it remains unclear how-lactamase induction ofP. vulgarisis suppressed on agar plates.
Quantitative results obtained by in vitro antimicrobial sus-ceptibility testing are very important for proper management with antimicrobial chemotherapy for patients with serious in-fectious diseases (29). At present, a variety of laboratory meth-ods are used for susceptibility testing. It is well known that the results of susceptibility tests are influenced by inoculum size, composition of the medium, concentrations of divalent cations, calcium, and magnesium, pH, and incubation conditions. When the final results of susceptibility tests are influenced by meth-odology, interpretation errors with serious clinical implications may occur. To avoid such risks, a series of procedures must be standardized to ensure accurate and reproducible results. Such efforts to develop standardized procedures for susceptibility tests have been reported (25). In the United States, the Na-tional Committee for Clinical Laboratory Standards develops standardized procedures, which are published in approved or proposed form, for susceptibility testing (20, 21).
The broth and agar dilution methods are tests commonly used to quantitatively measure the in vitro activities of antimi-crobial agents against a given bacterial isolate. Recently, we determined the MICs of expanded-spectrum cephems against clinical isolates of Proteus vulgaris by using both the broth microdilution method and the agar dilution method (A. Ohno, M. Datz, Y. Ishii, L. Ma, and K. Yamaguchi, Abstr. 38th In-tersci. Conf. Antimicrob. Agents Chemother., abstr. 101, 1998). Surprisingly, for several oximino-type expanded-spectrum cephems, the majority of MICs determined by the broth mi-crodilution method were over 10 dilutions higher than those obtained by the agar dilution method. Inevitably, MIC results produced by the broth dilution method were considered resis-tant, but MIC results obtained by the agar dilution method
were interpreted as susceptible. This phenomenon was tenta-tively designated the MIC gap phenomenon (Ohno et al., 38th ICAAC).
P. vulgaris produces class A -lactamases (CumA). The
cumA gene exists on the chromosome (24). CumA is very different from TEM- or SHV-type class A-lactamases in that oximino-type expanded-spectrum cephems are preferred sub-strates for CumA (2, 12, 27). Therefore, the MIC gap phenom-enon may be due to the activity of CumA. On the other hand, CumA is inducible, as are class C-lactamases (AmpC) pro-duced by most gram-negative bacteria. The induction of AmpC is closely linked to the recycling system for muramyl peptides released from the bacterial peptidoglycan (15). Factors in-volved in the peptidoglycan recycling system are AmpG, a transmembrane protein which acts as a permease for GlcNAc anhMurNAc-tripeptide (18) or -pentapeptide (5) degraded en-zymatically from the peptidoglycan, and AmpD, a cytosolic
N-acetylmuramyl-L-alanine amidase (once inside, these
mu-ramyl peptides are cleaved by AmpD, and tripeptide or pen-tapeptide is released and recycled as the precursor of pepti-doglycan [14]). -Lactams result in increased degradation of peptidoglycan, and GlcNAc-anhMurNAc-tripeptide or -pen-tapeptide or anhMurNAc-tripeptide or -pen-pen-tapeptide accumu-lates in the cytoplasm. Excess muramyl peptide binds to the transcriptional regulator protein AmpR and converts AmpR into an activator of-lactamases (1).
Datz et al. (4) demonstrated that induction of CumA utilizes a pathway identical to that observed for the control of AmpC synthesis in gram-negative bacteria. CumG and CumD in
P. vulgaris correspond to AmpG and AmpD, and the func-tions are shared with those of AmpG and AmpD. Like AmpR, CumR is also a transcriptional regulator protein; however, the function is not shared with that of AmpR (4).
In this study, we defined the association between the MIC gap phenomenon and the peptidoglycan recycling and  -lac-tamase induction systems ofP. vulgaris.
* Corresponding author. Mailing address: Department of Microbi-ology, Toho University School of Medicine, 5-21-16, Omori-nishi, Ota-ku, Tokyo, Japan 143-8540. Phone: 81-3-3762-4151. Fax: 81-3-5493-5415. E-mail: [email protected].
677
on May 15, 2020 by guest
http://jcm.asm.org/
(Part of this study was presented at the 38th Interscience Conference on Antimicrobial Agents and Chemotherapy [Ohno et al., 38th ICAAC].)
MATERIALS AND METHODS
Strains.A total of 42 isolates ofP. vulgariswere collected from five medical centers across Japan during 1996. These isolates were unselected or sequential isolates. We also usedP. vulgarisB317 and isogenic mutants with mutations in the peptidoglycan recycling and CumA induction systems (B317D, B317G, and B317R) and strains transcomplemented by the clonedampDE,ampG,ampR, andcumRgenes (B317D/pMD201, B317G/pMD301, B317R/pMD101, B317R/ pMD401, and B317R/pMD501) (4), which were kindly provided by M. Datz, Laboratory d’Enzymologie, Centre d’Inge´nie`rie des Prote´ines, Institut de Chimie, Universite´ de Lie`ge, Sart Tilman, Belgium. These strains were stored at⫺80°C in 15% glycerol until used. The characteristics of each strain and plasmid were as follows. B317 is a clinical isolate. B317D carries a defect in CumD (a cytosolic
N-acetylmuramyl-L-alanine amidase) and overproduces CumA. B317G carries a
defect in CumG (a permease for GlcNAc-anhMurNAc-tripeptide or -pentapep-tide) and must be induced for CumA. B317R carries a defect in CumR (tran-scriptional regulator protein for CumA; corresponds to AmpR for AmpC) and must be induced for CumA. pMD101 is a plasmid harboringampRandampC
fromCitrobacter freundii. pMD201 is a plasmid harboring the clonedampDE
gene fromEscherichia coli. pMD301 is a plasmid harboring the clonedampG
gene fromE. coli. pMD401 is a plasmid harboring the clonedampRgene from
C. freundii. pMD501 is a plasmid harboring the clonedcumRgene fromP. vul-garis.
Antimicrobial agents.Cefotaxime (CTX; Hoechst Japan Co., Osaka, Japan), cefteram (CEM; Toyama Chemical Co., Tokyo, Japan), cefpodoxime (CPD; Sankyo Co., Tokyo, Japan), ceftizoxime (ZOX; Hujisawa Yakuhin Co., Osaka, Japan), moxalactam (MOX; Shionogi & Co., Osaka, Japan), ceftibuten (CTB; Shionogi), cephaloridine (CER; Shionogi), and ampicillin (AMP; Meiji Seika Kaisha, Tokyo, Japan) were used in this study. Each antibiotic was in a powder form of known potency.
Susceptibility testing.All isolates were subjected to in vitro antimicrobial susceptibility tests by broth microdilution and agar dilution according to the guidelines of the NCCLS (20). Antimicrobial agents used for the determination of MICs against clinical isolates ofP. vulgariswere CTX, ZOX, MOX, CEM, CPD, and CTB. Furthermore, CTX, CER, and AMP were used forP. vulgaris
B317 and the isogenic mutants. Dehydrated Mueller-Hinton broth (MHB; Difco Laboratories, Detroit, Mich.) adjusted to the correct cation concentrations was used for broth microdilution testing. A 0.1-ml quantity of each antimicrobial agent dilution was dispensed into each well of a 96-well microdilution tray by using a dispensing device. For the agar dilution method, dehydrated Mueller-Hinton agar (Difco) was used. In this test, 18 ml of molten test agar was poured into petri plates containing 2 ml of the appropriate dilution of antimicrobial agent solution prepared at 10 times the desired final concentration and rapidly mixed. Prepared trays and plates were used on the same day. Cultures were adjusted to a 0.5 McFarland standard and diluted 1:10 in sterile saline. Inoculum suspensions were used simultaneously for both methods. The microdilution trays were inoculated using an automatic MIC-2000 inoculator (Dynatech Laborato-ries, Inc., Alexandria, Va.) so that the final inoculum was approximately 105 CFU/well. The agar plates were inoculated with an inoculum-replicating appa-ratus, Micro-Planter (Sakuma Seisakusho, Tokyo, Japan), at 104CFU/spot. The trays and plates were incubated for 48 h in ambient air at 35°C. The MIC represented the lowest concentration of antibiotic that completely inhibited visible bacterial growth and was read at 16 to 18 h and at 48 h after incubation. The effects of inoculum size and agar concentration on the MIC gap phenom-enon were also determined by using CTX for four clinical isolates andP. vulgaris
B317, B317D, B317D/pMD201, B317G/pMD301, and B317R/pMD501. The in-oculum sizes were 104, 105, 106, and 107CFU/spot for the agar dilution method. Bacto agar (Difco) at a concentration of 1.5, 0.75, 0.375, or 0.1875% was added to MHB, and MICs were compared with those obtained in MHB alone. The inoculum sizes were 104CFU/spot for 1.5 and 0.75% agars and 105CFU/ml for 0.375 and 0.1875% agars and MHB alone.
Induction of-lactamase and extraction of crude enzyme.Overnight cultures ofP. vulgarisB317 and isogenic strains in MHB were diluted 20-fold into 10 ml of fresh medium and incubated with continuous shaking at 35°C. After 2 h of incubation, CTX was added at a final concentration of 8g/ml. After a further 3 h of incubation, cells were harvested, washed three times with 0.1 M phosphate buffer (pH 7.0), and disrupted by ultrasonication. Broken cells were centrifuged at 100,000⫻gfor 30 min at 4°C, and the supernatant was used as the crude enzyme. The concentration of protein was determined by the method of Lowry et al. (19).-Lactamase activity was determined by a spectrophotometric method measuring the decrease in absorbance at 265 nm of CER (100M) at 30°C.
Determination of enzyme kinetic parameters.The values ofVmax,Km, andKcat of purified P. vulgaris-lactamase (Wako Pure Chemicals Industries, Ltd., Osaka, Japan) were determined by computerized spectrophotometry using var-ious absorbances at appropriate wavelengths for CER, CTX, CEM, CPD, ZOX, MOX, and CTB. The wavelength used for the photometric assay was that which yielded the maximum difference spectrum when an unhydrolyzed substrate was
scanned against a hydrolyzed substrate. The wavelengths determined for the agents were as follows: CER, see above; CTX, 264 nm; CEM, 262 nm; ZOX, 250 nm; CPD, 265 nm; MOX, 275 nm; and CTB, 260 nm.
RESULTS
Susceptibility tests for clinical isolates. The susceptibility test results are summarized in Table 1 as the MICs at which 50 and 90% of the isolates were inhibited (MIC50 and MIC90, respectively). The MIC gap phenomenon, i.e., MICs obtained by the broth microdilution method that were markedly higher than those obtained by the agar dilution method, was observed for most oximino-type cephems but not for other types of cephems. However, the MIC gap phenomenon was not evident at the MIC50 of ZOX despite the fact that this drug is an oximino type. On the other hand, the susceptibility test results obtained for 42 isolates by the E test and disk diffusion meth-ods were similar to those obtained by the agar dilution method and also showed the MIC gap phenomenon with respect to the broth microdilution method (data not shown). The MICs were also higher following incubation for 48 h. Furthermore, the MIC gap phenomenon was not observed for CEM and was only weak for CPD at 48 h in the MIC90evaluation.
[image:2.612.312.552.102.397.2]Among the isolates that did not show the MIC gap phenom-enon, 5 such isolates were noted when tested against CTX, 10 were noted with CEM, and 7 were noted with CPD. The MICs of these oximino-type cephems forP. vulgarisT4 were low in both methods. Thus, four isolates were found resistant to all three antibiotics by both methods, two were found resistant to CPD and CEM, and three were found resistant to only CEM
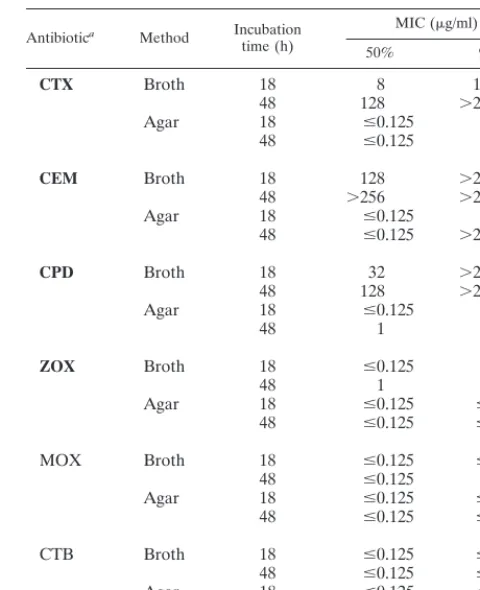
TABLE 1. Comparative susceptibilities of 42 clinical isolates of P. vulgaristo expanded-spectrum cephems, as determined
by broth microdilution and agar dilution methods
Antibiotica Method Incubation
time (h)
MIC (g/ml)
50% 90%
CTX Broth 18 8 128
48 128 ⬎256
Agar 18 ⱕ0.125 0.5
48 ⱕ0.125 8
CEM Broth 18 128 ⬎256
48 ⬎256 ⬎256
Agar 18 ⱕ0.125 0.5
48 ⱕ0.125 ⬎256
CPD Broth 18 32 ⬎256
48 128 ⬎256
Agar 18 ⱕ0.125 2
48 1 64
ZOX Broth 18 ⱕ0.125 2
48 1 32
Agar 18 ⱕ0.125 ⱕ0.125
48 ⱕ0.125 ⱕ0.125
MOX Broth 18 ⱕ0.125 ⱕ0.125
48 ⱕ0.125 0.5
Agar 18 ⱕ0.125 ⱕ0.125
48 ⱕ0.125 ⱕ0.125
CTB Broth 18 ⱕ0.125 ⱕ0.125
48 ⱕ0.125 ⱕ0.125
Agar 18 ⱕ0.125 ⱕ0.125
48 ⱕ0.125 ⱕ0.125
aExpanded-spectrum oximino-type cephems appear in bold.
on May 15, 2020 by guest
http://jcm.asm.org/
(data not shown). The “growth effect,” representing skip-ping of either a single or several consecutive concentrations, with insignificant growth at these concentrations but heavy growth at higher antimicrobial concentrations, was observed in the broth microdilution method for many isolates showing the MIC gap phenomenon (data not shown).
Susceptibilities of P. vulgarisB317, isogenic mutants with mutations in the peptidoglycan recycling and-lactamase in-duction systems, and transcomplemented strains to CTX, CER, and AMP.The MIC gap phenomenon was observed clearly for
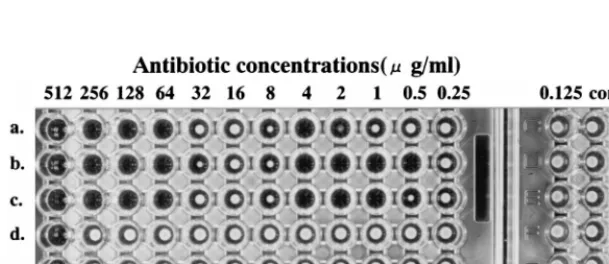
P. vulgaris B317, B317D, B317D/pMD201, B317G/pMD301, and B317R/pMD501 and slightly forP. vulgarisB317R/pMD101 but not for B317G, B317R, and B317R/pMD401 (Table 2). Furthermore, the phenomenon appeared with CTX but not with CER and AMP. A typical skip-growth effect was also ex-pressed in B317R/pMD501 (Fig. 1).
Effects of inoculum size and agar concentration on the MIC gap phenomenon.In the agar dilution method, MICs markedly increased when 107 CFU/spot was inoculated. Furthermore, the MICs significantly increased with decreases in agar con-centrations (Table 3).
Induction of-lactamase.-Lactamase activity was not de-tected in crude extracts fromP. vulgarisB317G, B317R, and B317R/pMD401. P. vulgaris B317, B317D/pMD201, B317G/ pMD301, B317R/pMD101, and B317R/pMD501 produced -lactamase only in the presence of inducing conditions.P.
vul-garisB317D produced-lactamase in the presence or absence of inducing conditions (Table 4).
Enzyme kinetic parameters. The Vmax values of purified P. vulgaris-lactamase for CER, CTX, CEM, and CPD were markedly higher than those for ZOX (Table 5). TheKcat/Km
values of CER, CTX, CEM, CPD were 90-, 20-, 7-, and 4-fold higher, respectively, than those of ZOX (Table 5). No -lac-tamase activity was detected for MOX and CTB.
DISCUSSION
It is well known that the results of susceptibility tests are influenced by various methodological factors, such as the type of medium, inoculum size, pH, temperature, and incubation time (29); however, major differences in MICs (the MIC gap phenomenon) between the broth microdilution method and the agar dilution method in susceptibility testing ofP. vulgaris
[image:3.612.52.554.92.251.2]against several-lactams, such as noted here, have not been reported. At present, the underlying mechanisms that cause such differences in MICs are not well understood. However, it is possible that the MIC gap phenomenon is related to the chromosomal class A-lactamase (CumA) ofP. vulgaris be-cause the phenomenon (i) is limited to oximino-type expand-ed-spectrum cephems, which are among the preferred sub-strates of CumA; (ii) is not observed with MOX and CTB, which are not degraded by CumA; and (iii) is observed only
[image:3.612.151.456.577.709.2]FIG. 1. Results of determination of MICs of CTX (a to c) and AMP (d to f) againstP. vulgarisB317R/pMD501 by the broth microdilution method. Reading of the MIC was done after 48 h of incubation.
TABLE 2. Comparative susceptibilities ofP. vulgarisB317 and other strains to various antibiotics, as determined by broth microdilution and agar dilution methods
Strain Genotype
MIC (g/ml)aof the following drug in the indicated test:
CTX CER AMP
Broth Agar Broth Agar Broth Agar
B317 Wild type 64 ⱕ0.125 256 128 512 256
B317D cumDmutation (ampDlike) 256 2 256 256 1,024 512
B317D/pMD201 ampDEfromE. coli 32 0.25 256 64 512 256
B317G cumGmutation (ampGlike) ⱕ0.125 ⱕ0.125 32 8 64 16
B317G/pMD301 ampGfromE. coli 32 ⱕ0.125 256 128 512 256
B317R cumRmutation (ampRlike) 0.5 ⱕ0.125 2 4 8 4
B317R/pMD101 ampR ampCfromC. freundii 4 ⱕ0.125 ⬎1,024 ⬎1,024 256 256
B317R/pMD401 ampRfromC. freundii 0.5 ⱕ0.125 2 4 4 2
B317R/pMD501 cumRfromP. vulgarisB317 64 (skip-growth) ⱕ0.125 256 256 512 512
aReading of the MIC was done after 48 h of incubation.
on May 15, 2020 by guest
http://jcm.asm.org/
slightly with ZOX, which is relatively stable in the presence of CumA despite being an oximino-type agent. Furthermore, the observation that one clinical isolate that does not produce CumA did not show the MIC gap phenomenon also seems to support this possibility. When examined by PCR, this iso-late possessed a 206-bp DNA segment containing the 165-bp
cumR-cumA intercistronic region (4); however, the 1.26-kb DNA fragment containingcumA(27) was not amplified (data not shown). The deletion or insertion might occur in thecumA
region of isolate T4. The MIC gap phenomenon was not ob-served in AmpC -lactamase-producing bacteria, such as C. freundii,Enterobacter cloacae,Serratia marcescens, and Pseudo-monas aeruginosa, or in the chromosomal CdiA-lactamase of
Citrobacter diversus, which belongs to Ambler’s class A and is very similar to CumA (data not shown). On the other hand, the MIC gap phenomenon disappeared inP. vulgarisB317G and B317R but reappeared in B317G/pMD301 and B317R/pMD501. The findings strongly suggested that the MIC gap phenomenon was specific forP. vulgarisand was due to the production of CumA-lactamase in sufficient quantities through the pepti-doglycan recycling and -lactamase induction systems ex-pressed within the broth microdilution environment.P. vulgaris
B317G is a mutant with defective CumG, a transporter of anh-MurNAc-tripeptide or pentapeptide which functions as an activator of thecumAtranscriptional regulator, CumR;P.
vul-garisB317R is a mutant with a defectivecumR gene. Further-more, B317G/pMD301 and B317R/pMD501 are the strains transcomplemented by clonedcumG gene andcumR genes, respectively.
The skip-growth effect may also support the relationship be-tween the MIC gap phenomenon and CumA. Laboratorians often encounter a similar skip-growth effect in routine suscep-tibility testing, and several reports have described similar find-ings (8, 17, 22). The underlying mechanisms of the skip-growth effect are diverse although not yet sufficiently defined. How-ever, the finding that the typical skip was noted inP. vulgaris
B317R/501 (carrying the clonedcumRgene) but not in B317R/ pMD401 (carrying the clonedC. freundii ampRgene) indicates that the skip-growth effect is likely to be seen in organisms in whichcumAhas been induced.
On the other hand, the reason for the significantly low MICs noted with the agar dilution method remains undetermined.
[image:4.612.53.293.91.209.2]P. vulgarisB317D is a mutant with defective function of CumD, a negative modulator of CumA, and constitutively produces a large quantity of CumA. The MIC of CTX for B317D in the agar dilution method was higher than that for the parent strain B317, suggesting that CumA is amply produced on agar plates because of the constitutive productivity of CumA in B317D. However, the MIC for B317D in the broth microdilution method was also higher than that in the agar dilution method, and the degree of the MIC gap phenomenon with both meth-ods was nearly identical to that seen with B317 or B317D/201 (carrying theampDEgenes). Because the production of CumA is suppressed on 1.5% agar plates by an unknown mechanism, even in B317D, which shows a high level of production of CumA, the MIC obtained by agar dilution was not as high as we would have predicted. Although there is no definite evi-dence, the possible suppression of CumA induction in agar may also be supported by the result that MICs increased in proportion to the decrease in agar concentrations. Moreover, the observation that the MIC gap phenomenon was not ob-served for CER and AMP may be due to the easy hydrolysis of these agents even by a small amount of CumA produced on agar plates. This conclusion is based on the finding that the narrow-spectrum cephems and penicillin serve as better sub-strates of CumA than do oximino-type expanded-spectrum cephems(24).However,oximino-typeexpanded-spectrumceph-ems showed markedly high MICs against a few clinical isolates in the agar dilution method (data not shown); therefore, an-other mechanism may also explain the MIC gap phenomenon. A few studies have examined the influence of agar on anti-microbial activity (9, 13). For example, antibiotics such as ami-noglycosides and polymyxins are bound to negatively charged groups on the agar molecule (6). Therefore, the MICs of such antibiotics shift to high values. Furthermore, the divalent cat-ion present in agar also reduces the antibacterial activity of tetracyclines (26). On the other hand, Ward et al. (28) reported that the addition of agar to susceptibility testing media lowered TABLE 3. Effect of agar concentrations on susceptibility
testing ofP. vulgarisagainst CTX
Strain MIC (g/ml)
aat the following % agar concnb:
1.5 0.75 0.375 0.1875 0
Isolate 1 2 64 512 512 512
Isolate 2 0.25 64 64 64 128
Isolate 3 ⱕ0.125 32 64 32 128
Isolate 4 0.5 32 256 256 256
B317 ⱕ0.125 ⱕ0.125 32 64 64
B317D 4 4 128 256 256
B317D/pMD201 0.25 0.25 2 32 32
B317G/pMD301 ⱕ0.125 ⱕ0.125 0.5 8 32
B317R/pMD501 ⱕ0.125 ⱕ0.125 32 32 64
aReading of the MIC was done after 48 h of incubation.
bA bacterial suspension was inoculated at a final inoculum size of 105CFU/ml with agar concentrations of 0, 0.1875, and 0.375%, while 104CFU/spot was used with agar concentrations of 0.75 and 1.5%.
TABLE 4. Expression of-lactamase activity inP. vulgarisB317 and other strains under noninducing and inducing conditions
Strain Plasmid-bornegene(s)
-Lactamase activity (mol/min/ mg of protein) under the
following conditions:
Noninducing Inducing (CTX at8g/ml)
B317 —a 0.069
B317D (cumDmutant) 0.209 0.158
B317D/pMD201 ampDE — 0.022
B317G (cumGmutant) — —
B317D/pMD301 ampG — 0.061
B317 (cumRmutant) — —
B317R/pMD101 ampRC — 0.061
B317R/pMD401 ampR — —
B317R/pMD501 cumR — 0.198
[image:4.612.311.553.93.158.2]a—, hydrolytic activity was not detected.
TABLE 5. Kinetic parameters for-lactams of theP. vulgarisCumA-lactamasea
-Lactam Vmax(M/min) Kcat(s⫺1) Km(M) Kcat/Km(M⫺1s⫺1)
CER 108 8.3 920 0.009
CTX 114 4.4 2502 0.002
CEM 44 0.85 1222 0.0007
CPD 114 1.5 4073 0.0004
ZOX 0.78 0.005 35 0.0001
aTheV
max,Kcat, andKmvalues were determined by a UV spectrophotometric method (see the text).
on May 15, 2020 by guest
http://jcm.asm.org/
[image:4.612.53.293.559.718.2]the MICs of amoxicillin-clavulanate against gram-negative ba-cilli. Regrettably, they did not discuss the reason for this finding. We examined and identified the effect of inoculum size on results obtained with the agar dilution method. The MICs of CTX againstP. vulgarisat an inoculum size of 107 CFU/spot were markedly higher than those at 104, 105, and 106 CFU/ spot. It is not clear at this stage whether the increase in MIC due to the effect of inoculum size is related to the production of large amounts of CumA, although a few reports have shown a relationship between the inoculum size effect and -lactama-ses (7, 30).
The duration of incubation was also noted to influence the results (29); MICs of oximino-type cephems were higher fol-lowing 48 h of incubation for more than half of the clinical isolates, B317, B317D/pMD201, B317G/pMD301, and B317R/ pMD501. Such an effect of incubation time is often recognized in relation to slow growth in fastidious bacteria (10, 16). How-ever,P. vulgarisis typically a nonfastidious bacterium; there-fore, the effect of incubation time does not seem to be due to the influence of growth rate.
On the other hand, in vitro susceptibility tests do not always reflect the in vivo situation (23), and in vitro versus in vivo discrepancies may also occur because of problems with varying pHs and antibiotic tissue concentrations at the site of infection (3). The present study does not identify the method (agar dilution or broth microdilution) that may reflect the clinical response when oximino-type expanded-spectrum cephems are used to treat infectious diseases caused byP. vulgaris. Ikeda et al. (11), using a murine experimental model of infection with
P. vulgaris, reported that the therapeutic effect of cefmenoxime (oximino-type expanded-spectrum cephem) in mice treated with a high dose was lower than that in those treated with a low dose and that-lactamase activity in the peritoneal cavity increased at higher cefmenoxime doses. Their study perhaps suggests that oximino-type cephems induce-lactamases ofP. vulgaris
in tissues at high concentrations of antibiotics during therapy. The most important aspect of any susceptibility test is the accurate detection of resistance, because resistance carries a strong probability of therapeutic failure. The MIC gap phe-nomenon has been shown to also affect the more commonly used E test and disk diffusion susceptibility tests. We think that the susceptible results obtained with these methods may be false. Therefore, if the clinical laboratory reports to the clini-cian the antimicrobial susceptibility results for oximino-type expanded-spectrum cephems againstP. vulgarisas susceptible by agar dilution, the E test, or disk diffusion methods, the patient may be exposed to ineffective antibiotics with side ef-fects, including the modification of isolates of normal flora.
In summary, the present study indicates that appropriate consideration of proper standardization should be enforced in susceptibility testing withP. vulgaris.
ACKNOWLEDGMENT
We thank Martina Datz for kindly providingP. vulgarisB317 and isogenic mutants.
REFERENCES
1.Bartowsky, E., and S. Normark.1991. Purification and mutant analysis of
Citrobacter freundiiAmpR, the regulator for chromosomal AmpC -lacta-mase. Mol. Microbiol.5:1715–1725.
2.Bush, K., G. A. Jacoby, and A. A. Medeiros.1995. A functional classification scheme forlactamases and its correlation with molecular structure. Anti-microb. Agents Chemother.39:1211–1233.
3.Cunha, B. A.1997. Problems arising in antimicrobial therapy due to false susceptibility testing. J. Chemother.9(Suppl. 1):25–35.
4.Datz, M., B. Joris, E. A. M. Azab, M. Galleni, J. van Beeumen, J. M. Fre`re, and H. H. Martin.1994. A common system controls the induction of very
different genes. The class-A-lactamase ofProteus vulgarisand the enter-obacterial class-C-lactamase. Eur. J. Biochem.226:149–157.
5.Dietz, H., D. Pfeifle, and B. Wiedemann.1997. The signal molecule for
-lactamase induction inEnterobacter cloacaeis the anhydromuramyl-pen-tapeptide. Antimicrob. Agents Chemother.41:2113–2120.
6.Ford, J. H., M. E. Bergy, A. A. Brooks, E. R. Garrett, J. Alberti, J. R. Dyer, and H. E. Carter.1955. Further characterizations of neomycin B and neo-mycin C. J. Am. Chem. Soc.77:5311–5312.
7.Gould, J. M., G. J. Heidecker, and J. J. LiPuma.1996. Nontypeable Hae-mophilus influenzaesusceptibility: effect of inoculum size and-lactamase production. Diagn. Microbiol. Infect. Dis.26:95–98.
8.Gresser-Burns, M. E., C. J. Shanholtzer, L. R. Peterson, and D. N. Gerding.
1987. Occurrence and reproducibility of the “skip” phenomenon in bacteri-cidal testing ofStaphylococcus aureus. Diagn. Microbiol. Infect. Dis.6:335–342. 9.Hanus, F. J., J. G. Sands, and E. O. Bennett.1967. Antibiotic activity in the
presence of agar. Appl. Microbiol.15:31–34.
10. Hartzen, S. H., L. P. Andersen, A. Bremmelgaard, H. Colding, M. Arpi, J. Kristiansen, T. Justesen, F. Espersen, N. Frimodt-Møller, and O. Bonnevie.
1997. Antimicrobial susceptibility testing of 230Helicobacter pyloristrains: importance of medium, inoculum, and incubation time. Antimicrob. Agents Chemother.41:2634–2639.
11. Ikeda, Y., Y. Fukuoka, K. Motomura, T. Yasuda, and T. Nishino.1990. Paradoxical activity of-lactam antibiotics againstProteus vulgarisin exper-imental infection in mice. Antimicrob. Agents Chemother.34:94–97. 12. Ishiguro, K., and K. Sugimoto.1996. Purification and characterization of the
Proteus vulgarisBlaA protein, the activator of the-lactamase gene. J. Bio-chem.120:98–103.
13. Iyer, R., and V. Iyer.1960. Effect of agar on the inhibition ofMicrococcus pyogenesvar.aureusby chlortetracycline and other antibiotics. Antibiot. Chemother.10:409–413.
14. Jacobs, C., B. Joris, M. Jamin, K. Klarsov, J. van Beeumen, D. Mengin-Lecreulx, J. van Heijenoort, J. T. Park, and S. Normark.1995. AmpD, essential for both-lactamase regulation and cell wall recycling, is a novel cytosolicN-acetylmuramyl-L-alanine amidase. Mol. Microbiol.15:553–559. 15. Jacobs, C., L. Huang, E. Bartowsky, S. Normark, and J. T. Park.1994.
Bacterial cell wall recycling provides cytosolic muropeptides as effectors for
-lactamase induction. EMBO J.13:4684–4694.
16. Kenny, G. E., and F. D. Cartwright.1993. Effect of pH, inoculum size, and incubation time on the susceptibility ofUreaplasma urealyticumto erythro-mycinin vitro. Clin. Infect. Dis.17(Suppl. 1):S215–S218.
17. Kerry, D. W., J. M. T. Hamilton-Miller, and W. Brumfitt.1976. Paradoxical effect of mecillinam onProvidencia stuartii. J. Antimicrob. Chemother.2:
386–388.
18. Lindquist, S., K. Weston-Hafer, H. Schmidt, C. Pul, G. Korfmann, J. Erick-son, C. Sanders, H. H. Martin, and S. Normark.1993. AmpG, a signal trans-ducer in chromosomal-lactamase induction. Mol. Microbiol.9:703–715. 19. Lowry, O. H., N. J. Rosebrough, A. L. Farr, and R. J. Randall.1951. Protein
measurement with the Folin phenol reagent. J. Biol. Chem.193:265–275. 20. National Committee for Clinical Laboratory Standards.1997. Methods for
dilution antimicrobial susceptibility tests for bacteria that grow aerobically, 4th ed. Approved standard. NCCLS document M7-A4. National Committee for Clinical Laboratory Standards, Wayne, Pa.
21. National Committee for Clinical Laboratory Standards.1998. Performance standards for antimicrobial susceptibility testing; eighth information supple-ment. NCCLS document M100-S8. National Committee for Clinical Labo-ratory Standards, Wayne, Pa.
22. Neu, H. C.1976. Mecillinam, a novel penicillanic acid derivative with unusual activity against gram-negative bacteria. Antimicrob. Agents Chemother.9:
793–799.
23. Nightingale, J.1987. Clinical limitations ofin vitrotesting of microorganism susceptibility. Am. J. Hosp. Pharm.44:131–137.
24. Pe´duzzi, J., A. Reynaud, P. Baron, M. Barthe´le´my, and R. Labia.1994. Chromosomally encoded cephalosporin-hydrolyzing-lactamase ofProteus vulgarisRO104 belongs to Ambler’s class A. Biochim. Biophys. Acta1207:
31–39.
25. Phillips, I., and A. King.1997. Standardization of susceptibility testing meth-ods. J. Chemother.9(Suppl. 1):13–18.
26. Price, K. E., Z. Zolli, Jr., J. C. Atkinson, and H. G. Luther.1957. Antibiotic inhibitors. II. Studies on the inhibitory action of selected divalent cations for oxytetracycline. Antibiot. Chemother.7:689–701.
27. Tamaki, M., M. Nukaga, and T. Sawai.1994. Replacement of serine 237 in class A-lactamase ofProteus vulgarismodifies its unique substrate speci-ficity. Biochemistry33:10200–10206.
28. Ward, P., S. Palladino, B. McLaren, R. J. Rathur, and J. C. Looker.1993. The effect of increased agar concentration in susceptibility testing media on MICs of antimicrobials for gram-negative bacilli. J. Antimicrob. Chemother.
31:1005–1007.
29. Wood, G. L.1995.In vitrotesting antimicrobial agents. Infect. Dis. Clin. N. Am.9:463–481.
30. Yeo, S. F., and D. M. Livermore.1994. Effect of inoculum size on the in vitro susceptibility to-lactam antibiotics ofMoraxella catarrhalisisolates of dif-ferent-lactamase types. J. Med. Microbiol.40:252–255.