1535-9778/07/$08.00⫹0 doi:10.1128/EC.00304-07
Copyright © 2007, American Society for Microbiology. All Rights Reserved.
NOTES
Sexual Pheromone Induces Diffusion of the Pheromone-Homologous
Polypeptide in the Extracellular Matrix of
Volvox carteri
䌤
Koichi Ishida*
Lehrstuhl Biochemie I, Universita¨t Regensburg, D-93040 Regensburg, Germany
Received 17 August 2007/Accepted 14 September 2007
The C-terminal domain of pherophorin II is homologous to the sexual pheromone ofVolvox carteriand is released from other domains during sexual induction. Green fluorescent protein fused to the C terminus of pherophorin II was located at the extracellular matrix directly surrounding the gonidium, the final target of the sexual-induction signal.
Volvox carteriis one of the simplest multicellular green algae,
comprising 2,000 to 4,000 biflagellate somatic cells arranged in a monolayer surface around a hollow sphere and 16 reproduc-tive cells called gonidia residing below the somatic cells (13). In the asexual life cycle, the matured gonidium undergoes 11 to 12 rapid cell divisions and gives rise to the somatic cells and gonidia; however, when exposed to a sexual pheromone, gonidia of the male and female spheroids undergo modified cell divisions, and sperm packets and eggs are produced in-stead of the asexual gonidia, respectively (12). The sexual pher-omone ofV. carteriis a 32-kDa glycoprotein that works at a very low concentration of less than 10⫺16M (20, 21), which
implies that a special mechanism amplifies the sexual-phero-mone signaling within the spheroid during the sexual-induc-tion process. Interestingly, this sexual-inducsexual-induc-tion process ne-cessitates a continuous 6- to 8-h exposure of the asexual spheroid to the sexual pheromone before the initiation of gonidial cleavage, and during this period, the sexual-induc-tion signal is assumed to be transmitted to the putative surface receptor of the gonidium (6).
The nature of the sexual-induction mechanism ofV. carteriis largely unknown. However, several molecular clues have been accumulated to date; the earliest biochemical responses of the spheroid to the sexual pheromone occurs in the somatic cells, which is evidenced by the synthesis of an extracellular matrix (ECM) protein, pherophorin II, as well as of many other pro-teins (1, 2, 3, 4, 8, 11, 20, 22). Pherophorin II is also a glyco-protein that consists of three domains: the N-terminal domain, whose sequence is related to a motif of another ECM protein, SSG185 (this motif is shared among other types of pherophor-ins as well); the polyproline spacer; and the C-terminal do-main, whose amino acid sequence is homologous to that of the sexual pheromone (8, 20). An impressive fact is that this C-terminal domain of pherophorin II is liberated from the other domains by proteolytic cleavage slowly after the sexual
induc-tion, with the same kinetics as those of the sexual-induction process (20). This phenomenon invokes the liberated phero-mone-like C-terminal domain to mediate the signaling process between the sexual pheromone and the gonidium. Therefore, in this work, green fluorescent protein (GFP) was fused to the C terminus and/or the N terminus of pherophorin II (5) andV.
carteriwas transformed with these fusion gene constructs (18)
(hereinafter referred to as the PIIC-GFP and PIIN-GFP trans-formants, respectively) to study the behavior of the each do-main of pherophorin II in the sexually induced spheroid.
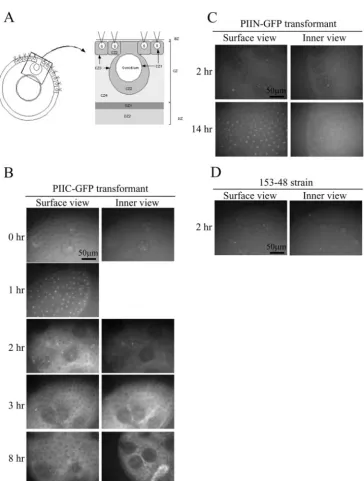
Figure 1A shows the scheme of the PIIC- and PIIN-GFP fusion proteins. Figure 1B shows a Western blot of the soluble fraction of the 153-48 strain (a recipient strain for transforma-tion), in which the PIIC- and PIIN-GFP transformants in 1% sodium dodecyl sulfate (SDS)–0.5 M NaCl were boiled for 5 min and detected by anti-GFP antibody, suggesting that GFP-tagged pherophorin II was synthesized successfully in both transformants. However, the Western blot of the ECM protein fraction, which was insoluble in boiled 1% SDS–0.5 M NaCl solution (called the “ECM ghost”) but thereafter solubilized in 1% Triton X-100–0.1 M EDTA at 65°C for 20 min as detected with anti-GFP antibody, showed that only the PIIC-GFP fusion protein was proteolytically cleaved appropriately (Fig. 1C). This result suggests that the insertion of GFP into the N-terminal domain of pherophorin II, which might cause a steric obstruction, inhibited further processing of this fusion protein. Thus, this transformant can serve as a control in distinguishing the localization of the cleaved C-terminal domain of the PIIC-GFP fusion protein from that of intact pherophorin II, al-though it has to be noted that it was still not a strict control because it is not known whether the PIIN-GFP fusion protein migrates through the ECM in the same way as the intact PIIC-GFP fusion protein. Next, I performed a pulse-chase labeling experiment with35SO
42⫺using the soluble fraction of
the ECM ghosts of the PIIC-GFP transformant after the Tri-ton X-100–EDTA treatment, prepared as described in the work of Wenzl and Sumper (20) (the sugar residues of the glycoprotein ofV. carteriECM is sulfated frequently [19]). As shown in Fig. 1D, the deduced molecular mass suggests that the band [band (b)] represents the cleaved C-terminal domain
* Present address: Nishii Initiative Research Unit, Frontier Research System, RIKEN, Hirosawa 2-1, Wako, 351-0198 Saitama, Japan. Phone: 81-48-467-4834. Fax: 81-48-467-9530. E-mail: [email protected].
䌤Published ahead of print on 28 September 2007.
2157
on September 8, 2020 by guest
http://ec.asm.org/
fused to GFP, although the cleavage process seems to be much faster than the endogenous one [band (d)], possibly because of the structural effect of the recombinant pherophorin II-GFP fusion. However, the result of Western blotting using anti-GFP
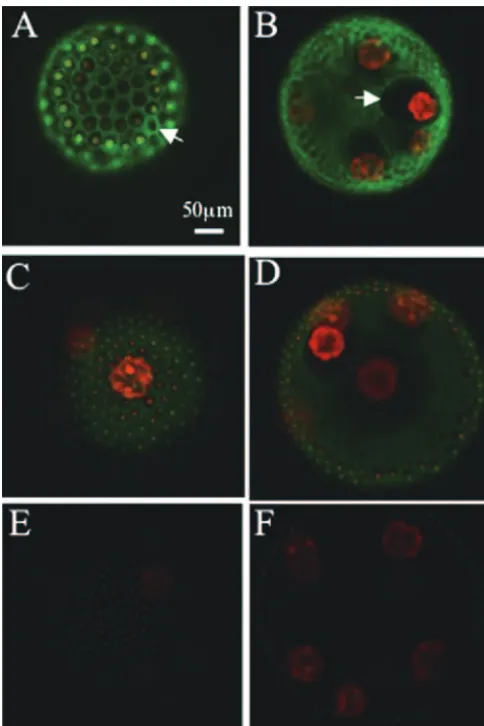
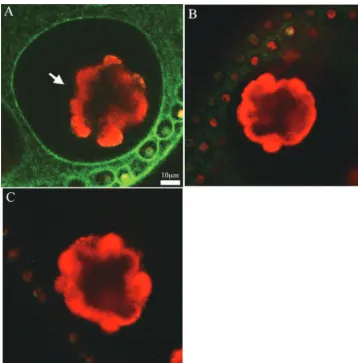
antibody (Fig. 1C) supports the notion that the band of the corresponding molecular mass is the cleaved C-terminal do-main fused to GFP. These results suggest that the PIIC-GFP fusion protein was also cleaved proteolytically, as is the native protein. Next, the GFP signal in the ECM structure (14) (shown in Fig. 2A) of the PIIC-GFP transformant was detected at each time point after the application of the sexual phero-mone with a fluorescence image system (Till Photonics, Gra¨felfing, Germany) as described by Fuhrmann et al. (5). The result in Fig. 2B shows that the GFP signal that was initially seen in the somatic cells (1 h later; bright spots in the surface view) spreads into amorphous cellular zone 2 (CZ2) and the fibrous CZ3 layer of the somatic cells (2 h later, surface view; signals in the compartment surrounding each somatic cell and the boundary of the compartment, respectively). Thereafter, the intense signal was detected at CZ3 of the gonidia (3 h later; signal at the compartment boundary surrounding each gonid-ium in the inner view). However, in the PIIN-GFP transfor-mant, the GFP signal was detected only weakly in CZ2 and -3 of the somatic cells (surface view) and in the inner space of the spheroid (inner view) even at 14 h after pheromone exposure, whereas rather intense signals were found in the somatic cells (Fig. 2C, surface view). These signals in the PIIC- and PIIN-GFP transformants were absent in the 153-48 strain (Fig. 2D). To obtain more-detailed images, the GFP signals of the PIIC-and PIIN-GFP transformants PIIC-and the 153-48 strain at 8 h after the application of the sexual pheromone were captured by a confocal laser scanning microscope (model LSM 5 Pascal; Zeiss, Jena, Germany). The surface view of the PIIC-GFP transformant showed the clear GFP signal in the honeycomb structure of CZ3 of the somatic cells (Fig. 3A), but the view of the inner section of the spheroid exhibited the GFP signal at CZ3 of the gonidia (Fig. 3B). Furthermore, a significantly intense signal was found in CZ4 and the deep zone of the PIIC-GFP transformant. In the PIIN-GFP transformant, how-ever, only a weak signal was detected in CZ2, -3, and -4 of the somatic cells (Fig. 3C) and the deep zone (Fig. 3D), in addition to the point-like signal in the somatic cells. The signal detected in strain 153-48 shows the background signal mainly from the endogenous chloroplast (Fig. 3E and F). A more important result was obtained from a magnified image of the gonidium of the PIIC-GFP transformant, in which the GFP signal was found at CZ1 (Fig. 4A). In this picture, cleaving gonidium was already detached from CZ1, but CZ1 is directly juxtaposed to the plasma membrane of the precleavage gonidium (14), which is supposed to receive the sexual-induction signal. Neither the PIIN-GFP transformant nor strain 153-48 showed such signals at CZ1s of their gonidia, although weak signals were seen in CZ2, -3, and -4 of the somatic cells of the PIIN-GFP transfor-mant (Fig. 4B and C).
The sexual-induction signal received first by the somatic cells must be transmitted to its final target, namely, the gonidium. Therefore, inevitably, the induction signal has to pass through CZ1, -2, and -3 of both the somatic cell and the gonidium. The results in this study showed that PIIC-GFP liberated from its parental molecule diffused gradually to CZ1 of the gonidium, which implies the role of pherophorin II as a transmitter/ amplifier of the sexual-induction signal. In addition, the results of the confocal images of the PIIN-GFP transformant, in which the PIIN-GFP fusion protein cannot be cleaved proteolytically
FIG. 1. (A) Schematic picture of the pherophorin II-GFP fusion proteins. Stippled boxes, signal peptide; hatched boxes, polyproline motif; arrow, proteolytic cleavage site. (B) Western blot of the phero-phorin II-GFP fusion proteins detected by the anti-GFP antibody, which was kindly provided by the Hegemann lab. Loading samples were prepared by boiling the spheroid in 1% SDS and 0.5 M NaCl for 5 min and centrifuging the solution at 13,200 rpm, and then the su-pernatants were subjected to SDS-polyacrylamide gel electrophoresis. The first lane contains a sample from strain 153-48, a recipient strain for transformation; the second and third lanes contain samples from the PIIC- and PIIN-GFP transformants, respectively. The arrow indi-cates the signal from the intact pherophorin II-GFP fusion protein. All samples were prepared 30 min after the application of the sexual pheromone. (C) Western blot of the pherophorin II-GFP fusion pro-teins detected by the anti-GFP antibody. Loading samples were pre-pared by incubating the insoluble pellet obtained after the sample preparation described for panel B in 0.1 M EDTA–10% Triton X-100 at 65°C for 20 min. All samples were prepared 10 h after the applica-tion of the sexual pheromone. The signal from the cleaved C-terminal domain of the PIIC-GFP fusion protein is marked by the letter a within the blot. The signal from the intact PIIC-GFP fusion protein is also seen above this band. (D) Results of SDS-polyacrylamide gel electro-phoresis showing fluorograms loaded with extracts from the PIIC-GFP transformant, which was pulse-chase labeled with35SO
4
2⫺. Briefly, the
pulse-labeling of the PIIC-GFP transformant with35SO
42⫺was
per-formed for a 1-h time period between 1 and 2 h after the addition of the sexual pheromone to the synchronized culture of the spheroids enclosing the precleavage gonidia. The samples were harvested after the chase period represented above each lane. The harvested samples were prepared as described for panel C. (a), intact PIIC-GFP fusion protein; (b), cleaved C-terminal domain of PIIC-GFP; (c), intact en-dogenous pherophorin II; (d), cleaved C-terminal domain of nous pherophorin II; (e), cleaved N-terminal domain of the endoge-nous pherophorin II and PIIC-GFP. K, thousand.
2158 NOTES EUKARYOT. CELL
on September 8, 2020 by guest
http://ec.asm.org/
and thus is expected to exhibit the same localization pattern as the intact PIIC-GFP fusion protein, suggests that intact phero-phorin II can diffuse into CZ2, -3, and -4 of the somatic cells as well as the deep zone of the spheroid; however, proteolytic cleavage is necessary for further diffusion of the C-terminal domain to CZ1 of the gonidium. This result correlates well with the fact that the inhibition of pherophorin II cleavage suppresses sexual induction (7).
Images of the sexually induced spheroid expressing the pherophorin-deep zone 1 (DZ1)-GFP fusion protein showing
the GFP signal at CZ1 of the gonidium, similar to Fig. 3B, were observed although not mentioned previously (4). The sexual induction ofV. carteritriggers a series of genes in addition to pherophorin II and pherophorin-DZ1, as mentioned earlier. Therefore, it is probable that the concerted actions of these multiple gene products is necessary to amplify and transmit the sexual-induction signal within the spheroid. This hypothesis explains that the applications of the artificially synthesized C-terminal domain of pherophorin II alone cannot induce the sexuality ofV. carteri(7, 20). However, the fact that there are
FIG. 2. (A) Schematic picture of theVolvoxECM structure. S, somatic cells; BZ, boundary zone; CZ, cellular zone; DZ, deep zone. (B to D) Fluorescence microscopy of the PIIC-GFP transformant, PIIN-GFP transformant, and 153-48 strain (negative control), respectively. GFP signals were detected at each indicated time point after the application of the sexual pheromone. Only parts of the spheroids are shown. The bright spots in the surface view of the PIIC-GFP transformant at 1 h shows the positions of the somatic cells (the inner view was omitted because the image was the same as at 0 h). The large dark balls in the pictures are gonidia. Initially the GFP signal was seen in the somatic cells (1 h), but later it spread to CZ2 and -3 of the somatic cells (2 h) and then to CZ3 of the gonidia (3 h). The scale bar applies to all panels.
on September 8, 2020 by guest
http://ec.asm.org/
two types of mutant phenotype, one refractory to the sexual induction and another constitutively sexually induced, corre-sponds well to the expression status of the pherophorin II transcript, which implies an important role of pherophorin II in the sexual induction ofV. carteri(7, 20).
In this study, the GFP signals detected in CZ3, CZ4, and the deep zone of the PIIC-GFP transformant were much more intense than that of CZ1 of the gonidium. One possibility is that the GFP moiety of the cleaved C-terminal domain of the PIIC-GFP fusion protein slowed the migration through CZ3 of both the somatic cell and the gonidium into CZ1 of the gonidium. CZ3 is composed of a coherent network of ECM filament (16), and therefore it might work as a barrier which restricts the migration of the molecules into the CZ3 compart-ment and might have caused the accumulation of PIIC-GFP at CZ3 (Fig. 4A). In contrast, the ECM structures of the amor-phous zones (CZ2, CZ4, and DZ4) and looser fibrous layer (DZ1) do not seem to impede the migration of this molecule significantly (Fig. 3B and 4A) (16).
Even small accumulations of pherophorins at CZ1 of the gonidium seem to amplify the initial signal from the quite low concentration (below 10⫺16M) of the sexual pheromone by
orders of magnitude; however, the localizations of the cleaved C-terminal domain of pherophorin II in the deep zones and CZ4 are conspicuous and thus indicate a certain function of this molecule in these regions. One of the hypotheses is that the C-terminal domain of pherophorin II is involved in the remodeling of the internal ECM structure of the sexual spher-oid, as shown in the case of pherophorin-DZ1 (4), which might generate some signal to be transmitted to gonidia, although the existence of such a signal driven by ECM remodeling has not yet been clarified (9).
Another hypothesis is related to the wound-healing process of the spheroid, which is inferred from two considerations of the evolution of the sexual-induction system ofV. carteri, de-scribed blow. First, in the volvocine-lineage algae, the unicel-lular Chlamydomonas reinhardtii and other colonial species, such asPleodorina starrii, are induced sexually by the lack of a nitrogen source in the environment but not by a factor such as sexual pheromone (15, 16). However, V. carterideveloped a more intricate ECM structure than those species in the course of multicellular evolution (14, 19), and the sexual-induction system ofV. carteri seems to have evolved in relation to the establishment of such a complex ECM structure, considering that the sexual pheromone triggers the expression of several of the ECM proteins, including several types of the pherophorin family proteins (9, 10). In reference to this, many genes en-coding pherophorin-related proteins have been isolated from the volvocine-lineage algae ranging fromC. reinhardtii to V.
carteri(10, 17), which implies that the sexually induced
phero-phorins ofV. carteriwere evolutionarily derived from the an-cestral genes of these pherophorin-related ECM proteins (no-tably, one of them, GAS28 of C. reinhardtii, is expressed specifically in the gamete [17]), which originally would be the fundamental components of the ECM architecture and unre-lated to the pheromone-based sexual-induction process.
Second, the expression of all of the sexually induced genes of
V. carteritested to date are also induced by the wounding of the
spheroid (1, 3, 4, 9, 10, 11). Therefore, one possible scenario is that the sexual-induction system ofV. carterihas evolved from the fundamental ECM components which were originally used in the wound-healing process of the spheroid to facilitate their survival; that is, the same set of genes as those expressed in the wound-healing process have become utilized in the sexual-induction process in the course of the evolution of the elabo-rate ECM architecture required for more-sophisticated multi-cellularity. Several unknown extra factors must be involved in the sexual-induction process because the wounding of the spheroid per se does not induce the sexuality of the spheroid (1), but at least both of these processes appear to share a common signal transduction pathway, which makes sense given that both wound healing and durable zygote formation, the ultimate consequence of sexual induction, occur as the results of a response to an adverse environment (1). In fact, it is realistically presumable that both of these processes occur at the same time in the severely desiccating pond in nature in order to carry out sexual induction in the unwounded spheroid and to bring up the gametes within the normal sexual spheroid
FIG. 3. Surface and inner views of the PIIC-GFP transformant (A and B), the PIIN-GFP transformant (C and D), and strain 153-48 (E and F) obtained by confocal laser scanning microscopy are shown. The green signals indicate the GFP signal, whereas the red signals are derived from the autofluorescence of the chloroplast. The scale bar applies to all panels. GFP signals at CZ3 are indicated by white arrows.
2160 NOTES EUKARYOT. CELL
on September 8, 2020 by guest
http://ec.asm.org/
of the next generation, which will raise the adaptive value of this species.
Taking the two considerations mentioned above into ac-count, GFP signals detected in CZ3, CZ4, and the deep zone of the PIIC-GFP transformant (Fig. 3A and B and 4A) can be interpreted as follows. These ECM regions are the fundamen-tal extracellular framework and the interstitial tissues that sup-port the multicellular architecture; therefore, the signals in these regions might represent the function of this molecule in the wound-healing process rather than in the sexual-induction process. Such a versatile function of this molecule would be of great use for the survival of the spheroid in nature, which requires both of these processes at the same time, as men-tioned above. Nevertheless, the wound-healing mechanism would be of great advantage also for other colonial species of volvocine-lineage algae even if they do not have the phero-mone-based sexual-induction system. Although the existence of such a wound-healing mechanism in those colonial species has not yet been examined, at least it is apparent that there is a molecular link between the wound-healing and sexual-induc-tion processes of V. carteri, and therefore, the evolutionary
study which connects these two processes might shed light on the riddle of the sexual-induction mechanism ofV. carteri.
I thank all the people in Manfred Sumper’s and Peter Hegemann’s laboratory for the discussions, technical support, and miscellaneous work that helped me during my stay in Germany. I am also much obliged to Peter Hutzler of the Max Planck Institute, Munich, Ger-many, for generous support with laser scanning microscopy.
This work was supported by the Deutsche Forschungsgemeinschaft (SFB521).
REFERENCES
1.Amon, P., E. Haas, and M. Sumper.1998. The sex-inducing pheromone and wound trigger the same set of genes in the multicellular green algaVolvox. Plant Cell10:781–789.
2.Aono, N., T. Inoue, and H. Shiraishi.2005. Genes specifically expressed in sexually differentiated female spheroids ofVolvox carteri. J. Biochem.138: 375–382.
3.Ender, F., A. Hallmann, P. Amon, and M. Sumper.1999. Response to the sexual pheromone and wounding in the green algaVolvox: induction of an extracellular glycoprotein consisting almost exclusively of hydroxyproline. J. Biol. Chem.274:35023–35028.
4.Ender, F., K. Godl, S. Wenzl, and M. Sumper.2002. Evidence for autocat-alytic cross-linking of hydroxyproline-rich glycoproteins during extracellular matrix assembly inVolvox. Plant Cell14:1147–1160.
5.Fuhrmann, M., W. Oertel, and P. Hegemann.1999. A synthetic gene coding
FIG. 4. (A) Inner view showing the gonidium of a highly magnified confocal image of the PIIC-GFP transformant. The GFP signal at CZ1 is indicated by the white arrow. The scale bar applies to all panels. (B and C) Magnified confocal images of the gonidia of the PIIN-GFP transformant and strain 153-48, respectively. The signal output levels of these two panels were enhanced using Adobe Photoshop CS (Adobe Systems Inc., San Jose, CA) to exhibit the similar autofluorescence intensity of the chloroplast to that of panel A for better comparisons.
on September 8, 2020 by guest
http://ec.asm.org/
for the green fluorescent protein (GFP) is a versatile reporter in Chlamydo-monas reinhardtii. Plant J.19:353–361.
6.Gilles, R., C. Gilles, and L. Jaenicke.1984. Pheromone-binding and matrix-mediated events in sexual induction ofVolvox carteri. Z. Naturforsch.39c: 584–592.
7.Godl, K., A. Hallmann, A. Rappel, and M. Sumper.1995. Pherophorins: a family of extracellular matrix glycoproteins fromVolvoxstructurally related to the sex-inducing pheromone. Planta196:781–787.
8.Godl, K., A. Hallmann, S. Wenzl, and M. Sumper.1997. Differential target-ing of closely related ECM glycoproteins: the pherophorin family from
Volvox. EMBO J.16:25–34.
9.Hallmann, A.2003. Extracellular matrix and sex-inducing pheromone in
Volvox. Int. Rev. Cytol.227:131–182.
10.Hallmann, A.2006. The pherophorins: common, versatile building blocks in the evolution of extracellular matrix architecture in Volvocales. Plant J. 45:292–307.
11.Hallmann, A., P. Amon, K. Godl, M. Heitzer, and M. Sumper.2001. Tran-scriptional activation by the sexual pheromone and wounding: a new gene family fromVolvoxencoding modular proteins with (hydroxyl)proline-rich and metalloproteinase homology domains. Plant J.26:583–593.
12.Hallmann, A., K. Godl, S. Wenzl, and M. Sumper.1998. The highly efficient sex-inducing pheromone system ofVolvox. Trends Microbiol.6:185–189. 13.Kirk, D. L.1998.Volvox: molecular-genetic origins of multicellularity and
cellular differentiation. Cambridge University Press, Cambridge, United Kingdom.
14.Kirk, D. L., R. Birchem, and N. King.1986. The extracellular matrix of
Volvox: a comparative study and proposed system of nomenclature. J. Cell Sci.80:207–231.
15.Matsuda, Y., T. Shimada, and Y. Sakamoto.1992. Ammonium ions control gametic differentiation and dedifferentiation inChlamydomonas reinhardtii. Plant Cell Physiol.33:909–914.
16.Nozaki, H., T. Mori, O. Misumi, S. Matsunaga, and T. Kuroiwa.2006. Males evolved from the dominant isogametic mating type. Curr. Biol.16: R1018–R1020.
17.Rodriguez, H., M. A. Haring, and C. F. Beck.1999. Molecular characteriza-tion of two light-induced, gamete-specific genes from Chlamydomonas rein-hardtii that encode hydroxyproline-rich proteins. Mol. Gen. Genet.261:267– 274.
18.Schiedlmeier, R., R. Schmitt, W. Mu¨ller, M. M. Kirk, H. Gruber, W. Mages, and D. L. Kirk.1994. Nuclear transformation ofVolvox carteri. Proc. Natl. Acad. Sci. USA91:5080–5084.
19.Sumper, M., and A. Hallmann.1998. Biochemistry of the extracellular ma-trix ofVolvox. Int. Rev. Cytol.180:51–85.
20.Sumper, M., E. Berg, S. Wenzl, and K. Godl.1993. How a sex pheromone might act at a concentration below 10⫺16M. EMBO J.12:831–836. 21.Tschochner, H., F. Lottspeich, and M. Sumper.1987. The sexual inducer of
Volvox carteri: purification, chemical characterization and identification of its gene. EMBO J.6:2203–2207.
22.Wenzl, S., and M. Sumper.1986. Early event of sexual induction inVolvox: chemical modification of the extracellular matrix. Dev. Biol.115:119–128.
2162 NOTES EUKARYOT. CELL