The morphological ‘forms’ of
Palpopleura lucia
(Drury)
areseparate speciesas evidenced
by
DNA
sequencing
(Anisoptera: Libellulidae)
A. Mitchell¹ and M.J . Samways²
ReceivedApril2,2004 / Revised andAcceptedOctober22,2004
INTRODUCTION
Intense debateoverthe taxonomicstatusof
Palpopleura
lucia(Drury)
has con-tinued since thedescription
of P. lucia and P.portia
asseparate species in 1773(P1NHEY,
1962).
PINHEY(1951,
1985)
treated “lucia“portia,”
and the West African“graffei”
as “forms” of P. lucia. Currenttaxonomyposits
twosubspecies,
P. I. lucia and P. I.
portia.
While females of thetwosubspecies
aredifficulttodistin-1School of Molecularand CellularBiosciences, UniversityofNatal,PrivateBag X01, Scottsville, 3209,SouthAfrica;— e-mail:[email protected]
2
DepartmentofEntomologyandNematology, FacultyofAgriculturalandForestry Sciences, UniversityofStellenbosch,PrivateBag XI,7602Matieland,SouthAfrica;
-e-mail;[email protected]
P. lucia isawidespreadAfrican sp. withacheckered taxonomichistory.Currently2 ‘forms’orsubspecies,P. l. lucia and P. l.portiaarerecognized, althoughdebateoverthe
taxonomicstatusof these taxahashardlyletupoverthe last 230years.The 2 ‘forms’ showdistinctive wingpatterndifferencesalthoughotheraspectsof theirmorphology
areverysimilar.Theycan occurhighly sympatricallyatsomelocalities in southern
Af-rica,aswell aselsewhere,thusraisingthequestionof whethertheyaretwospeciesor oneperhapswith balancedpolymorphism.DNAsequencedata from the ITS2 and COI geneswerecollected fromspecimensof both these ‘forms’toassess morerigorouslythe taxonomicstatusof thesetaxa.Thecloselyrelated P.deceptor (Calv.)andP.jucunda (Ramb.)wereincluded in the dataset toprovideabaseline forcomparisons. Specimens from all 4taxawerefrompoolsof the floodplainof the SabieR., KrugerNational Park,SouthAfrica,andwerepotentiallyabletointerbreed. Bothphylogenetic
analy-sesandcomparisonsofsequencedivergencelevelsstrongly supportthehypothesisthat the 2 ‘forms’ ofP. luciaarereproductivelyisolatedand should be accorded fullspecies
guish,
the males show distinc-tivepatterning
on thewings
(Fig.
1).
Thesubspecies
aresympatric
overmuch of theirgeographical
ranges, from the EasternCape
toEast and WestAfrica, suggesting
thatthey
are notbiological
sub-species. Although
atsome lo-calities bothoccur as adultsatthesame
pool, although
P. l.lucia overallseemstoprefer
warmerconditions
(PINHEY,
1985).
Thequestion
of their taxonomicstatus is alsoeco-logically significant,
because ifthey
wereindeednonspecific,
this wouldpossibly
be balancedpolymorphism,
unknowntodate in theOdonata,raising
thequestion
of what selection pressuresmight
beresponsible
formaintaining
suchpolymorphism.
Weinvestigate
here the taxonomicstatusof thesetaxausing
DNAsequencesof the second internal transcribed
spacer
region (ITS2)
of the nuclear ribosomal DNA repeatregion,
and the mitochondrialcytochrome
oxidase I(COI)
gene.MATERIAL AND METHODS
WesequencedDNAfrom21malespecimens, includingtenspecimensofP.I. lucia
,fivespecimens of P. I.portia,three specimensofP.jucunda (Ramb.)and threespecimensofP.deceptor (Calv.). Im-portantis that allspecimenswerecollected onthesameday (14March 2002 withinakm of eachother, and thereforehighly sympatric, alongthe floodzoneof the SabieRiver, KrugerNational Park,South Africa(24° 59’S,31°28’E.,320ma.s.1.),andpreserved directlyinto 100% ethanol.
TotalgenomicDNAwasextracted fromapproximately20 mgofflightmusclesbymeansof the QiagenDNeasy™Tissue Kit.
For the ITS2gene,initial PCRamplificationsusedtheprimersITS5 (5’-GGAAGTAAAAGTCG-TAACAAGG-3’)and 28S-25R(5’-TATGCTTAAAYTCAGCGGGT-3’),yieldingPCRfragmentsof
ap-proximately900bpinlength.EachPCRfragmentwasfractionatedon a1%agarosegel,2-5piofgel
con-tainingthe PCRfragmentwasremoved withasterilemicropipette tipand used inareamplification
reac-tionusinganinternalupstreamprimer,ITS2-1F(5’-CATGAACATCGACATYTTGAACGC-3’),and theoriginaldownstreamprimer (28S-25R) yieldingaPCRfragmentofapproximately520bp.Theprimers
used for the COIgenewereJerry/Cl-J-2183(5’-CAACATTTAliltGATTTTTTGG-3’)andGeorgina/ Cl-N-2786(5’-GGATAATCTGAATAWCGWCG-3’), yieldingPCRfragments647bpinlength.
PCRwasperformedon aPerkin ElmerGeneAmpPCRSystem2400 under thefollowingconditions:
94°C for 3minutes,32cycles (or24cycles forreamplifications)of(94°Cfor 30sec.,55°C for 30sec,72°C for 60-90sec),72°C for 7min,4°C hold. Reaction volumes were30-50pl,and the reaction mixture
con-tained:IXPCRbuffer,2mMMgCl,, 200pMof eachdNTP,10pmolof each PCRprimer,0.7 units of RocheTaqDNApolymerase,andapproximately 250ngofgenomicDNA/RNA.PCR reactionswere
cleanedusing QiagenPCRpurificationcolumns and directly sequencedin both directions usingthe ABIBigDye™Terminatorv3.0Cycle SequencingKit, followingthe manufacturer’srecommended Fig. 1. WingpatterningofPalpopleural.lucia(above)and
conditions.Sequenceswerevisualized on anABI 3100 GeneticAnalyser.
DNAsequencechromatogramswereedited andcontigsassembledusingthe Stadenpackage (STA-DEN, 1996).Consensus sequences wereautomatically aligned usingClustalX(THOMPSONetah, 1997)and then manuallycorrectedusingSe-Al v2.0a7(RAMBAULT, 2001). Phylogenetic analyses
wereperformedunder both the maximumparsimony(MP)and maximum likelihood(ML)criteria
us-ingPAUP—4.0M0(SWOFFORD, 1998).A10,000iterationpartition homogeneitytestwasfirst per-formed. Each datasetwasanalysed separately,andfinally,the combineddatasetwasanalysed.Branch and bound searcheswereperformedunderMP,while200-replicaterandom addition sequence heuristic searcheswereperformedunder ML. Theextentofsupportfor internal nodeswasestimatedbymeans
of bothparsimony-basedanddistance-based (ML-model) bootstrap analyses,eachemploying 1,000 iterations.FollowingFRATIetal.(1997),16 different models of sequence evolutionwerecomparedfor their fittoeach dataset(ITS2,COI and combineddata)bymeansof likelihood ratio tests.The model chosenforMLanalysiswastheonewith the least number of freeparameters,and whichwasnot sig-nificantlydifferent from themostcomplexmodel,thegeneraltime-reversiblemodel(GTR +1+G).For all three data sets, the best modelprovedtobe theHasegawa,Kishino and Yano model(HASEGAWA etal., 1985),withgamma-distributedrates(HKY85+G).Modelparameterswereestimated from the
data for each datasetseparately.Alternativephylogenetic hypotheseswereassessedby performingML constraint searchesandcomparingtheresultingtreeswith the ML tree,usingKishino-Hasegawa (K-H)andShimodaira-Hasegawa(S-H)tests,asimplementedin PAUP*4.
RESULTS AND DISCUSSION
All DNA sequences
reported
herearedeposited
in GenBank(accession
numbers ITS2: AY582759-AY582777;COI;AY582778-AY582796).
The datasetconsisted of 474 nucleotides of ITS2 sequence and 595 nucleotides of COI sequence, fora total of 1069 sites. For ITS2 therewere59 variable sites(12.4%
ofthetotal)
while for COI therewere 143 variable sites(24%).
Of the 21specimens
sampled,
two lacked ITS2 sequence and anothertwo lacked COI sequence, duetosequencing
difficulties.Analyses
of the combined datasettherefore usedonly
the 17complete
Comparison ITS 2 COI
mean s min max n1 mean s min max n
Withindeceptor - - ... 0.78 0.09 0.67 0.84 3 Withinjucundu 3.40 2.43 0.68 5.34 3 Withinportia 0.30 0.13 0.28 0.46 3 2.72 1.29 0.67 4.37 10 Within lucia 0.86 0.46 0.22 1.59 15 0.74 0.36 0.17 1.51 28 luciavsportia 3.66 0.18 3.28 3.91 18 10.82 0.32 10.25 11.61 40 jucundavslucia 4.35 0.16 4.14 4.60 6 10.17 0.47 9.16 11.24 24 jucundavsportia 3.75 0.00 3.75 3.75 3 10.37 0.45 9.67 11.11 15 deceptorvsjucunda 9.76 0 9.76 9.76 1 10.71 0.43 10.09 11.27 9 deceptorvslucia 9,17 0.36 8.83 9.72 6 13.46 0.26 12.94 14.02 24 deceptorvsportia 9.48 0.17 9.40 9.61 3 12.07 0.50 11.09 12.77 15
1For ITS2anumber ofspecimensshowed identical sequences (e.g,withinP.deceptorandP.jucunda) and allduplicatesequenceswereeliminated beforecomparisonofdivergencelevels.
TableI
Uncorrectedpairwise divergencevalues(%)
Comparison
mean s
ITS 2
min max n1 mean s
COI min max n Withindeceplor _ _ _ _ - 0.78 0.09 0.67 0.84 3 Withinjucunda - - - 3.40 2.43 0.68 5.34 3 Withinportia 0.30 0.13 0.28 0.46 3 2.72 1.29 0.67 4.37 10 Within lucia 0.86 0.46 0.22 1.59 15 0.74 0.36 0.17 1.51 28 luciavsportia 3.66 0.18 3.28 3.91 18 10.82 0.32 10.25 11.61 40 jucundavslucia 4.35 0.16 4,14 4.60 6 10.17 0.47 9.16 11.24 24
jucundavsportia 3.75 0.00 3.75 3.75 3 10.37 0.45 9.67 II.11 15
deceptorvsjucunda 9.76 0 9.76 9.76 1 10.71 0.43 10.09 11.27 9 deceptorvslucia 9.17 0.36 8.83 9.72 6 13.46 0.26 12.94 14.02 24 deceptorvsportia 9.48 0.17 9.40 9.61 3 12.07 0.50 11.09 12.77 15
sequences.The
partition homogeneity
testwasnon-significant (p
=0.095),
there-fore the datawereanalysed
bothseparately
and in combination.Table I summarizes the uncorrected
divergence
values observed inpairwise
com-parisons
among all sequences. For ITS2 sequences, the averagepairwise divergence
levels between P. l. lucia and P. l.portia
arenotstatistically
differenttothose be-tween thesesubspecies
and P.jucunda.
Infact,
the maximumdivergence
observed betweenspecimens
of P I. lucia and P. I.portia (3.91%)
isgreater than the maxi-mumdivergence
observed between P.jucunda
and P. I.portia (3.75%).
Themoredistantly
related P.deceptor
shows 8.8- 9.7%divergence
from all othertaxa. Asimilarpatternisseenfor COI sequences, with themeanand maximum
divergenc-es between P. I. lucia and P. I.
portia (10.8%
and 11.6%respectively) being
greater than thecorresponding
values forcomparisons
between eithersubspecies
and P.jucunda (10.3%
and 11.1%respectively). Again, comparisons involving
Pdeceptor
give larger
values(up
to14.2%).
For COI data
alone, parsimony analysis
ofthe 17taxa datasetproduced
30 short-est treesoflength
=202steps,Cl(excluding
uninformativecharacters)
=0.77,and RI=0.91.
For ITS2 dataalone, parsimony analysis
of the 17taxadatasetproduced
78 shortesttreesoflength
=72steps,Cl(excluding
uninformativecharacters)
=0.96,
and RI =0.98. Bothanalyses
recovered the four nominatetaxawith >90%bootstrap
supportin everycase. The branches thatcollapsed
in the strict consen-sus treeswerethoseindicating relationships
within the fourtaxa.Analyses
of thecomplete
19taxadatasetsfor eachgenewereinagreementbutgavenoadditionalinformation,
thereforethey
arenotconsidered further here. Maximum likelihoodanalyses
gavealmost identical resultstothe MPanalyses,
theonly
differencesbe-ing
in theweakly supported relationships
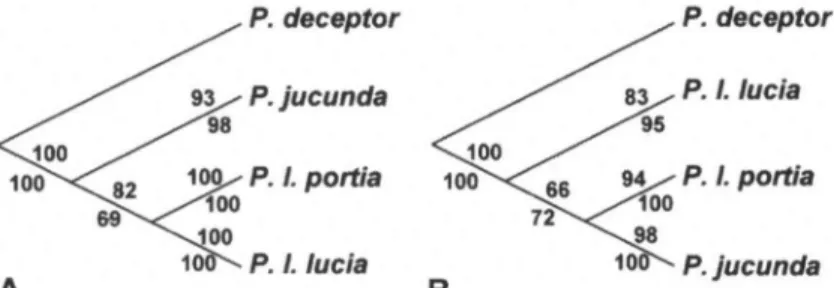
within the fourtaxa.Figure
2A summarizes therelationships
recovered among the fourtaxafor the COI dataalone,
under both MP and MLcriteria,
whileFigure
2B shows thesameforthe ITS2 data alone. Thesetreesdiffer in their
placement
of P. I.portia.
The COI dataplaces
P. I. lucia and P. I.portia
assister groups, withbootstrap
support of 82% under ML and 69% underMP,while the ITS2 dataplaces
P I.portiaand Fig.2.Summaryofrelationshipsamongthe foursympatric taxarecovered under both MLand MP criteria.Bootstrap supportlevels underMLandMPcriteriaareshown above andbe-lowbranches, respectively: (A)COI dataonly;
-(B)ITS2 dataonly. Palpopteura
P.
jucunda
as sister groups, withbootstrap
supportof 66% under ML and72%under MRThe combined dataset recov-ered thesame
relationships
among taxa as the COI
analysis,
withFigure
3clearly showing
theun-ambiguous
and broadseparation
of thesubspecies
of P. lucia. Vis-ualinspection
of internalbranchlengths
confirms the initialfind-ings suggested by
examinationofpairwise divergence levels,
thatthe distance between P. I. lucia and P. l.portia
isatleastasgreatasthat between either taxonand P. ju-cunda. Themonophyly
of eachsubspecies
also isstrongly
sup-ported.
Totestthe
significance
of dif-ferences in MLscorebetween thecompeting hypotheses,
heuristic searcheswere conductedon the COI(and combined)
datasets whileconstraining
thetopology
tothat shown inFigure
2B(ob-tained with the ITS2
data),
and vice versa.Resulting
trees werecompared
tothe MLtreefor each datasetby
meansof K-H andS--F1tests
(Tab. II).
All of theteststatisticswerenon-significant. Thus, despite
seem-ingly
reasonablebootstrap
supportvalues for both of thecompeting topologies,
noneof the datasetshas theresolving
powertodistinguish
among them. Further-more,reverse constraint searches in which either P. l. luciaorP. l.portia
were con-strainedtobenotmonophyletic yielded
treeswhichweresignificantly
different from the MLtrees(K-H
tests, p=0.000-0.003;
S-H tests, p=0.001-0.018).
These resultssuggestthat thetwo
subspecies
of P. lucia each should be accorded fullspecies
status.Specimens
ofthesetaxawerecollectedfromsympatric
popula-tions, apparently
with fullopportunity
forinterbreeding. Indeed,
somespecimens
of P. I. lucia and P. I.portia
were evencollected withinafew metres ofeachother from thesamepool. Sampling
of suchspecimens provides
avery conservativetestof the taxonomicstatusof thesetaxa. Thatis,
while limitedgeneflow between thetaxa Fig. 3. MLtreeforthe combined data set, -In L=
2,949.27958.HKY85 modelparameterswere asfollows: A=0.25575,C= 0.22220,G=0.22755,T=0.29450; Ts/Tv ratio=3.54019(k=7.1506344); shape parameter
atthis
locality
wouldnotnecessarily
indicate thatthey
wereconspecific,
theapparentlack ofgeneflow between thetaxaprovides
very strongevidence thatthey
arereproductively
isolated,
and therefore fullspecies.
Instead of
sampling specimens
fromacross thegeographical
range of thespecies,
we choseto concentrateonspecimens
thathad been collected insympatryin ordertoavoid theconfounding
effectsofgeographical
variation. Inessence, we have examined the “worst case scenario” ofsympatric populations
andyetwestill obtained very clear results thatPalpopleura
l. lucia and P. l.portia
arereproductively
isolatedfrom oneanother,and should beregarded
asseparatespecies, Palpopleura
lucia(Drury,
1773)
and P.portia (Drury, 1773).
ACKNOWLEDGEMENTS
We thank KLAAS-DOUWE DIJKSTRA and VIOLA CLAUSN1TZER fordiscussion,and STUART TAYLOR forscanningin thefigureof thewingpatterning.Financial supportwasfrom the National ScienceFoundation,South Africa.
REFERENCES
FRAT1, E,C.SIMON,J, SULLIVAN & D.L.SWOFFORD, 1997. Evolution of the mitochondrial cytochromeoxidaseII genein Collembola. J. mol Evol. 44; 145-158.
HASEGAWA, M.,H. KISHINO & T.YANO, 1985.Datingof thehuman-ape splitting bya molecu-lar clock of mitochondrial DNA. J. mol. Evol.21: 160-174.
PINHEY, E.C.G.,1951. Thedragonfliesof Southern Africa. Transv. Mus. Mem. 5; 1-335.
P1NHEY, E.C.G.,1962. Adescriptive catalogueof the Odonataof the African continent(upto De-cember 1959). Publfoescult. Co. Diam. Angola59: 1-323.
PINHEY, E.C.G., 1985. Asurveyof thedragonflies (Odonata)of South Africa,2:Anisoptera,J.ent. Soc. sth.Afr.48: 1-48
RAMBAULT, A., 2001. Se-Al.SequenceAlignmentEditor. hnp.llevolve.zoo.ox.ac.uklSoftwarelSe-AUmain.html
STADEN, R„1996. The StadenSequence Analysis Package.Mol. Biotech. 5;233-241.
SWOFFORD, D.L., 1998. PAUP*Phylogeneticanalysis using parsimony *(amlothermethods).
Sin-auerAssociates,Sunderland/MA.
THOMPSON, J.D.,T.J.GIBSON,F.PLEWNIAK,F.JEANMOUGIN &D.G.HIGGINS,1997. The ClustalX windows interface:flexiblestrategiesformultiplesequencealignmentaidedby quality analysistools. Nucl. Acids Res. 24: 4876-4882.
TableII
Summaryoftestsfor thesignificanceof like-lihood differences between trees with the
topologiesshown inFigures2A and 2B
K-Htest S-H test
Combined data p=0.770 p=0.803 ITS2only p=0.624 p=0.778 COIonly p=0.849 p=0.824