Initial Steps in RNA Processing
and Ribosome Assembly Occur
at Mitochondrial DNA Nucleoids
Daniel F. Bogenhagen,1,*Dwight W. Martin,2,3and Antonius Koller31Department of Pharmacological Sciences, Stony Brook University, Stony Brook, NY 11794-8651, USA 2Department of Medicine, Stony Brook University, Stony Brook, NY 11794-8160, USA
3Proteomics Center, Stony Brook University, Stony Brook, NY 11794-8691, USA *Correspondence:[email protected]
http://dx.doi.org/10.1016/j.cmet.2014.03.013
SUMMARY
Mammalian mitochondrial DNA (mtDNA) resides in
compact nucleoids, where it is replicated and
tran-scribed into long primary transcripts processed
to generate rRNAs, tRNAs, and mRNAs encoding
13 proteins. This situation differs from bacteria and
eukaryotic nucleoli, which have dedicated rRNA
transcription units. The assembly of rRNAs into
mi-toribosomes has received little study. We show that
mitochondrial RNA processing enzymes involved
in tRNA excision, ribonuclease P (RNase P) and
ELAC2, as well as a subset of nascent mitochondrial
ribosomal proteins (MRPs) associate with nucleoids
to initiate RNA processing and ribosome assembly.
SILAC pulse-chase labeling experiments show that
nascent MRPs recruited to the nucleoid fraction
were highly labeled after the pulse in a
transcrip-tion-dependent manner and decreased in labeling
intensity during the chase. These results provide
insight into the landscape of binding events required
for mitochondrial ribosome assembly and firmly
establish the mtDNA nucleoid as a control center
for mitochondrial biogenesis.
INTRODUCTION
Evolution of eukaryotes from two cohabiting ancient bacteria re-sulted in organisms capable of generating biochemical energy in a much more effective manner than their prokaryotic ancestors (Lane and Martin, 2010). During this process, the mitochondrial genome lost the vast majority of genes, some of which migrated to the nucleus, leaving a minimal set of genes involved exclu-sively in production of the mitochondrial respiratory complexes. Our 16.6 kb human mitochondrial DNA (mtDNA) genome en-codes only 13 polypeptides and is entirely dependent on prop-erly coordinated expression of over 200 nuclear genes for its replication, transcription, and repair and for translation of the handful of mitochondrial mRNAs. Approximately 80 nuclear-encoded proteins are imported to construct a mitochondrial ribosome using the small 12S and 16S mitochondrial RNAs as
scaffolds. Similar challenges are encountered in assembling respiratory complexes, as NADH dehydrogenase, for example, is built by combining approximately 38 nuclear-encoded sub-units with 7 mtDNA-encoded polypeptides that tether the com-plex to the inner membrane (Mimaki et al., 2012).
The combination of mitochondrial transcript mapping with the sequencing of human mtDNA (Anderson et al., 1981) re-sulted in the tRNA punctuation model of mitochondrial RNA processing in which rRNAs and mRNAs are produced by endo-nucleolytic cleavage from long polycistronic transcripts, often as byproducts of excision of closely flanking tRNA genes (Ojala et al., 1981). We now understand the roles of the mtRNA poly-merase and its accessory factors, TFB2M and TFAM, in tran-scription initiation (Falkenberg et al., 2007). We know far less concerning the pathways for RNA processing events required to generate mitochondrial rRNAs, mRNAs, and tRNAs. The extensive transcription of both strands of mtDNA (Aloni and At-tardi, 1971) creates potential problems, including avoidance of RNA-RNA duplex formation as well as accurate discrimination of functional RNAs from nonfunctional products. Processing of primary transcripts is initiated by the action of mitochondrial ribonuclease P (mtRNase P), which contains three polypep-tides, MRPP1, MRPP2, and MRPP3 (Holzmann et al., 2008). RNase P cleaves the 50end of tRNAs, which are subsequently released by action of ELAC2 (RNaseZL;Brzezniak et al., 2011; Rossmanith, 2012). Other work has characterized a transient complex of SUV3 helicase with polynucleotide phosphorylase (PNPase) that degrades unneeded mtRNAs (Borowski et al., 2013). We still lack an understanding of many aspects of mtRNA metabolism (Rackham et al., 2012; Rorbach and Min-czuk, 2012), including the intramitochondrial location and timing of events in RNA processing and the assembly of rRNAs into ribosomes.
mtDNA circles (5 mm) are folded into nucleoids comprised of the DNA bound with mitochondrial transcription factor A (TFAM), mitochondrial single-stranded DNA binding protein (mtSSB), and associated enzymes that act on the DNA ( Bogen-hagen, 2012). Nucleoids are compact structures with diameters of about 100–120 nm nestled between adjacent infoldings of the inner membrane (Brown et al., 2011; Kukat et al., 2011; Kopek et al., 2012). Our proteomic analysis of biochemically purified native and crosslinked nucleoids revealed association of many of the known proteins involved in mtDNA replication and transcription with these structures (Bogenhagen et al.,
2008; Wang and Bogenhagen, 2006). Proteins involved in mtRNA processing and turnover, as well as mitoribosomal pro-teins (MRPs), were discovered in nucleoid preparations. For example, our previous nucleoid preparations contained pep-tides derived from two subunits of mtRNase P, polynucleotide phosphorylase, and three proteins with RNA helicase do-mains associated with nucleoids, DHX30, DDX28, and SUV3. The apparent yeast homolog of DDX28 has been implicated in mitoribosome assembly (De Silva et al., 2013). Other recent studies identified another nucleoid-associated protein (GRSF1) that we identified as a component of processing bodies (Antonicka et al., 2013; Jourdain et al., 2013; Wang and Bogenhagen, 2006).
Finding MRPs associated with nucleoids suggested that bind-ing of ribosomal proteins to rRNA to initiate ribosome assembly may begin at the nucleoid while transcription and initial RNA inci-sion events continue. This is consistent with other recent reports of the copurification of ribosomal components with nucleoids
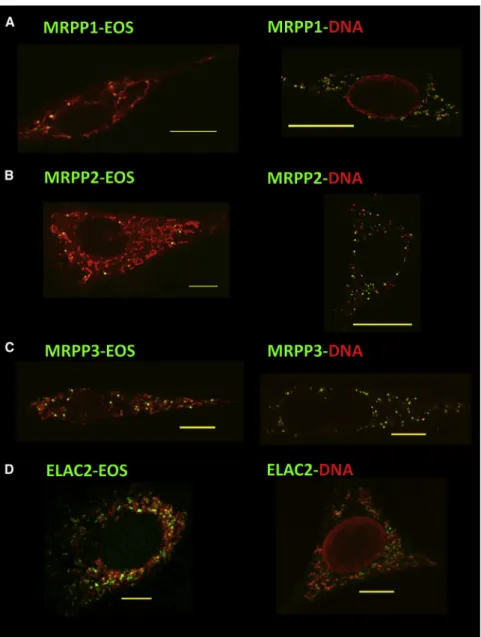
Figure 1. RNase P and ELAC2 Are Localized Near mtDNA Nucleoids, Where They Can Initiate Processing of Primary Transcripts
(A–D) Open reading frames for mouse MRPP1 (A), MRPP2 (B), MRPP3 (C), and ELAC2 (D) were cloned in pGS-Eos, and expression of fusion proteins in mouse 3T3 cells was induced with mifepristone for 7 hr. Fusion proteins were imaged with general mitochondrial staining (left) or with antibodies directed against DNA and Alexa 568-labeled secondary antibodies (right). Individual cells exhibited highly variable numbers of ex-pressed Eos-labeled foci. To visualize mitochon-dria, either cells were stained with MitoTracker Red (A and B), or fixed cells were labeled with antibodies to the 70 kDa subunit of respiratory complex II along with Alexa 568-labeled second-ary antibodies against mouse (C and D). Scale bars represent 10mm.
(He et al., 2012; Rorbach et al., 2008). Very little is known concerning the as-sembly of mitoribosomes, which contain a larger number of proteins bound to smaller rRNAs compared to that bound to bacterial ribosomes (Koc et al., 2010). A number of MRPs have identifiable homologs in bacterial ribosomes, while others do not. In both bacteria (Shajani et al., 2011) and in eukaryotic nucleoli (Fatica and Tollervey, 2002; Kressler et al., 2010) ribosome assembly begins with the binding of ribosomal proteins to nascent rRNA while transcription pro-ceeds. The process may be similar in mitochondria. In this paper, we use stable isotope pulse labeling (Ong et al., 2002) to identify a set of newly synthesized polypeptides selectively associated with mtDNA nucleoids. This work provides great insight into the initial dynamics of mitoribosome assembly at the mtDNA nucleoid, substantiating the model that nucleoids are critical centers for mitochondrial biogenesis.
RESULTS
RNase P and ELAC2 Are Localized Near Nucleoids We conducted imaging experiments to determine whether pro-teins observed in biochemical preparations of mtDNA nucleoids were enriched in the vicinity of nucleoids. First, we studied the intramitochondrial locations of RNase P subunits, which do not form a tightly associated complex (Holzmann et al., 2008), and of ELAC2. The open reading frames encoding these four mouse proteins were fused to the fluorescent reporter protein mEos in a mifepristone-inducible expression vector we used to study TFAM distribution (Brown et al., 2010, 2011). All four fusion proteins were observed as punctate foci within mitochondria near nucleoids (Figures 1andS1available online). This nucleoid
proximity resembles that of the helicase DHX30 (Wang and Bogenhagen, 2006) as well as mtDNA degradosome components (Borowski et al., 2013) and GRSF1 (Antonicka et al., 2013;
Jour-dain et al., 2013). The precise spatial relationships among these components of the RNA processing and turnover machinery will require additional studies. The presence of these complexes in the vicinity of a particular nucleoid and their protein composition may be dynamic depending on concurrent transcription activity. Biochemical Separation of Nucleoid- and Ribosome-Enriched Fractions
We refined methods used to purify nucleoids to explore their as-sociation with RNA processing enzymes and with incompletely processed nascent transcripts as well as potential ribosomal assembly intermediates. He et al. (2012) also used nonionic iodixanol buoyant density gradients to show a close association between nucleoids and mitoribosomes. This work, similar to our initial fractionation ofXenopusnucleoids using metrizamide gra-dients (Bogenhagen et al., 2003), is based on initial fractionation studies byBarat et al. (1985). In our hands, these gradients are easily overloaded when a crude mitochondrial lysate is applied. Therefore we developed a two-step fractionation procedure us-ing glycerol gradient sedimentation to separate nucleoids from the bulk of free ribosomes, followed by parallel iodixanol gradient analyses of both fractions. This is essentially a two-dimensional fractionation dependent on sedimentation coefficient and then buoyant density.
Highly purified mitochondria washed with high salt to remove adherent cytoplasmic ribosomes (Kellems et al., 1975) were lysed with 2% Triton X-100, and the lysate was subjected to sedimentation for 3–4 hr through 15%–40% glycerol gradients containing 0.2% Triton X-100. These gradients separate mtDNA-containing nucleoids, collected on a density shelf in frac-tion 1 (Figure 2A), from most ribosomes, which sediment about one-third of the length of the tube, and from free proteins, which remain in the upper third of the gradient, although the brief centri-fugation does not permit separation between the large and small ribosomal subunits (Figures 2B and 2C). Large protein com-plexes and nonribosomal RNA-protein comcom-plexes are also found in the ribosome-enriched fractions.
We consistently observed some mtRNA, including rRNAs, associated with nucleoids in fraction 1 of glycerol gradients (Figure 2). Most ribosomes were physically separated from nucleoids by sedimentation. Mitoribosomes associate with the inner membrane with receptors such as OxaI (Liu and Spremulli, 2000) and are widely distributed within mitochondria, not confined to nucleoids (Vogel et al., 2006). We used quantitative PCR (qPCR) primer pairs to test whether incompletely pro-cessed, newly synthesized RNA precursors would copurify with nucleoids. The tRNA junction primers (Figure S2) were de-signed to flank uncleaved RNase P processing sites for tRNAs bordering 12S and 16S rRNA and ND1 mRNAs. Real-time qPCR analysis of total RNA from gradient fractions revealed greater enrichment of incompletely processed tRNA junctions in the nucleoid fraction compared to that observed for mature RNAs (Figure 2C). A substantial fraction of partially processed mtRNA was separated from nucleoids during glycerol gradient sedimentation. Our results support the implication of the tRNA punctuation model that mtRNA processing by RNase P and ELAC2 begins at the nucleoids, although partially processed RNAs can be physically separated from nucleoids. It is not known how long partially processed mtRNAs remain near the
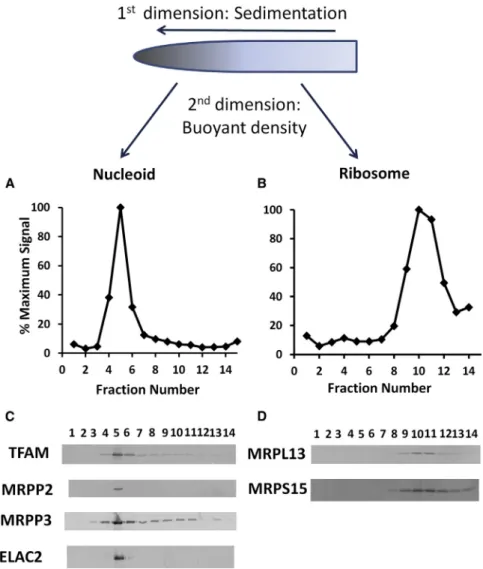
Figure 2. Incompletely Processed mtRNAs Are Enriched in the Nucleoid Fraction
HeLa mitochondria were lysed with Triton X-100 and subjected to sedimen-tation on a 15%–40% glycerol gradient as described (Lee et al., 2013). (A) A sample of fraction 1, which contained the peak of SYBR Green DNA fluorescence, was organically extracted to purify DNA, which was cleaved with HindIII endonuclease and analyzed by agarose gel electrophoresis. The marker lane (M) contains a 1 kb DNA ladder with bands ranging from 0.5 to 10 kb. The sizes of the human mtDNA HindIII fragments in kb are shown on the right.
(B) Samples (150ml) of the indicated fractions (#3 was omitted) were processed to purify RNA for electrophoresis on an agarose gel following denaturation and reaction with formaldehyde. The positions of 16S rRNA, 12S rRNA, and tRNA are indicated on the right. The sizes of RNA mobility markers in lane M are shown on the left.
(C) RT-PCR detection of mtRNA processing intermediates and mature 12S, 16S rRNAs, and ND1 mRNA in glycerol gradient fractions. The brackets over lane 1 and lanes 7–11 indicate fractions enriched in nucleoids and mitochondrial ribosomes, respectively, for the second-dimension analysis in Figure 3.
nucleoid, possibly in processing bodies. Processing body components are not a well-defined set of proteins. While they are found in the nucleoid fraction, as shown below, they do not remain quantitatively associated with mtDNA during our handling procedures.
To further purify nucleoids and ribosomes, we added a second dimension to the fractionation scheme. The DNA- and RNA-en-riched fractions from glycerol gradients were loaded in parallel on separate iodixanol gradients on the same day as the cell frac-tionation to avoid any freeze-thaw steps. Following overnight centrifugation, analysis of the nucleoid-containing gradient frac-tions showed that TFAM, RNase P, and ELAC2 proteins remain associated with the large mtDNA complexes, although some proteins apparently dissociate and begin to float to lower den-sities (Figures 3A and 3C). Parallel analysis of the ribosome-enriched material revealed ribosomal proteins MRPL13 and MRPS15 associated with the peak of SYBR Green II RNA fluo-rescence (Figures 3B and 3D).
SILAC Pulse-Chase Analysis of Mitochondrial Ribosome Assembly
We used proteomics to test the hypothesis that some newly syn-thesized MRPs associate with nascent rRNAs at the nucleoid.
Figure 3. RNase P and ELAC2 Are Persis-tently Associated with mtDNA Nucleoids
(A and B) Fraction 1 from the glycerol gradient in Figure 2 containing mtDNA nucleoids, and pooled fractions 7–11, enriched in mitochondrial ribosomes, were separately loaded on pre-formed gradients containing 20%–40% iodixanol. Following equilibrium centrifugation, gradients were fractionated and samples were analyzed for nucleic acids by staining with SYBR Green I for DNA (A) or SYBR Green II for RNA (B). Samples (10ml) of gradient fractions were analyzed by SDS-PAGE, and indicated proteins were detected by immunoblotting.
(C) TFAM as well as mitochondrial RNase P subunits and ELAC2 were detected in the peak nucleoid fraction.
(D) Ribosomal proteins MRPL13 and MRPS15 found in the initial glycerol gradient fractions 7–11 copurified with rRNA.
Previous studies have documented the orderly association of ribosomal proteins with nascent rRNA in bacteria (Shajani et al., 2011) and eukaryotic nucleoli (Fatica and Tollervey, 2002). Talkington et al. (2005) used ordered addition of bulk 15N-labeled proteins with mass spectrometry analysis to follow the clas-sical in vitro pathway for bacterial ribo-some assembly. A recent publication applied mass spectrometry to study in vivo assembly of bacterial ribosomes as well (Chen and Williamson, 2013).
We devised a pulse-chase labeling approach to ask whether certain MRPs might preferentially associate with nascent rRNA at the nucleoid. Radioactive pulse labeling could be used to address this ques-tion, but identifying a large number of proteins with this approach is cumbersome. As an alternative, we used pulse labeling with stable isotopes detected using mass spectrometry (SILAC;
Ong et al., 2002) to interrogate large numbers of proteins in a sin-gle experiment. Pulse labeling with stable isotopes has been used to measure protein half-lives in eukaryotic cells (Boisvert et al., 2012) but has rarely been used to monitor assembly of proteins into specific large complexes using cell fractionation methods. SILAC is a method to compare two specimens, one density labeled and one unlabeled. In a pulse-labeling SILAC experiment, the preexisting light proteins provide the internal control. We reasoned that, after pulse labeling with a mixture of13C-arginine and -lysine, if an MRP synthesized during the pulse is found preferentially in the nucleoid fraction, we could infer that this protein is involved in ribosome assembly at the nucleoid. For HeLa cells growing with a doubling time of 20– 24 hr, the total protein content may be expected to increase by about 25% in a 5–6 hr labeling period. We incubated cells for 6 hr with13C-arginine and -lysine, each labeled at 6 positions. In duplicate experiments, we found that a 6 hr pulse resulted in heavy/light (H/L) ratios of 0.217 and 0.239, in good agreement
with each other and with our prediction of the overall labeling in-tensity. We then purified mitochondria and employed our two-dimensional fractionation scheme to obtain nucleoid-enriched and ribosome-enriched fractions. In this experimental scheme, it is not necessary that either fraction be highly purified. Proteins were trypsin-digested to obtain peptides that were analyzed us-ing standard liquid chromatography-tandem mass spectrometry (LC/MS-MS) methods to provide information on the content of
isotope labels in the parent molecular ions as well as fragmenta-tion patterns to confirm peptide identity.
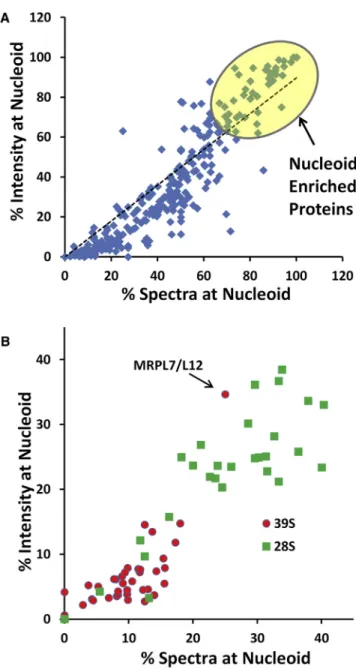
Application of this method provided a deep source of data on the relative abundance of proteins in either the nucleoid-en-riched or ribosome-ennucleoid-en-riched fraction using both spectral counts and extracted ion current intensity (Figure 4A). These two quan-titative parameters showed a strong linear correlation (p < 0.05), permitting us to make a number of observations based only on protein abundance, irrespective of the SILAC labeling analysis below. We identified a number of nucleoid-enriched proteins (Tables 1andS1). This list included all three subunits of RNase P, ELAC2, and other proteins previously identified as having
Figure 4. Identification of Nucleoid-Enriched Proteins
(A) Output from MaxQuant analysis of MS data for proteins in nucleoid-associated and ribosome-enriched fractions was mined to determine the percent of total spectral counts and the percent of total extracted ion current intensity in the nucleoid-associated fraction. Top-scoring proteins identified by both criteria were considered nucleoid-enriched proteins, listed inTables 1 andS2.
(B) Spectral count and intensity data were processed as in (A) are shown for 77 MRPs.
Table 1. Nucleoid-Enriched Proteins with Roles in Replication, Transcription, and RNA Processing
Protein ID Total Spectra Spectra at Nucleoid (%) Intensity at Nucleoid (%) Mito Nucleic Acid Synthesis
spjQ96RR1jPEO1_HUMAN 12 100.0 100.0 spjQ9UHN1jDPOG2_HUMAN 10 100.0 100.0 spjP54098jDPOG1_HUMAN 8 100.0 100.0 spjQ96QE5jTEFM_HUMAN 17 100.0 100.0 spjQ00059jTFAM_HUMAN 72 98.6 100.0 spjQ9H5Q4jTFB2M_HUMAN 74 68.9 75.2 spjQ04837jSSBP_HUMAN 134 77.6 69.8 spjO00411jRPOM_HUMAN 135 74.1 69.8 spjQ8WVM0jTFB1M_HUMAN 56 66.1 62.2
Mito Nucleic Acid Processing
spjQ96CM3jRUSD4_HUMAN 12 100.0 100.0 spjQ8WV60jPTCD2_HUMAN 10 100.0 100.0 spjQ9BQ52jRNZ2_HUMAN 26 92.3 98.8 spjO95900jTRUB2_HUMAN 18 94.4 98.2 spjQ96E29jMTER1_HUMAN (mTERF3) 37 94.6 95.5 spjQ7L0Y3jMRRP1_HUMAN 35 88.6 92.3 spjQ99714jHCD2_HUMAN (MRPP2) 77 75.3 71.1 spjQ9NVV4jPAPD1_HUMAN 193 77.2 81.1 spjO75127jPTCD1_HUMAN 47 83.0 80.4
Helicases and/or Processing Body
spjQ8IYB8jSUV3_HUMAN 50 100.0 100.0 spjQ8TCS8jPNPT1_HUMAN 127 97.6 97.6 spjQ7L2E3jDHX30_HUMAN 423 91.0 94.7 spjQ9NUL7jDDX28_HUMAN 33 93.9 91.0 spjQ12849jGRSF1_HUMAN 36 88.9 90.6 Chaperones spjP08238jHS90B_HUMAN 46 100.0 100.0 spjP07900jHS90A_HUMAN 13 100.0 100.0 spjP08107jHSP71_HUMAN 16 81.3 94.7 spjP11142jHSP7C_HUMAN 42 83.3 72.4 spjQ96EY1jDNJA3_HUMAN 172 69.2 53.6 spjP61604jCH10_HUMAN 14 71.4 12.8
Detailed information on these and other enriched proteins can be found in
roles in mtDNA replication, transcription, and RNA processing, with abundant hits on proteins considered to be part of the degradosome and of processing bodies such as SUV3L1, PNPT1, and GRSF1. These proteins confirm the efficiency of our rapid separation method. In a series of experiments, we ob-tained MS spectral data on 77 of the approximately 80 MRPs, although some small basic MRPs do not yield large numbers of diagnostic tryptic peptides. As expected, only a minor frac-tion of MRPs was found in the nucleoid fracfrac-tion (Figure 4B). Unexpectedly, polypeptides of the 28S subunit were consider-ably more abundant in the nucleoid fraction than those of the 39S subunit. Interestingly, one exception to this is provided by MRPL7/MRPL12, which may have a greater presence at
Figure 5. SILAC Proteomic Analysis of Proteins in the Nucleoid and Ribosome-Enriched Fractions
(A) An example is shown for a doubly charged peptide derived from MRPL28 detected in both the nucleoid and ribosome fractions. Two forms of this peptide were observed as separate nested sets of molecular ions. Those containing lysine (12C-6), beginning at m/z 1266.63*, are resolved from those containing lysine (13
C-6), beginning at m/z 1269.64#. Integration of the area under the curve for the extracted ion current is shown in the lower panels ofFigure 5A.
(B) Proteins detected by proteomics in both the nucleoid and ribosomal enriched fractions (111; Table S2) were ranked according to the overall ratio of H/L amino acid content in the nucleoid and ribosome fractions after the 6 hr pulse. The inset shows a box-and-whisker plot using the quartile method to indicate that the 39S, 28S, and non-ribosomal (other) protein groups display statisti-cally distinct pulse label ratios by ANOVA.
the nucleoid based on its role in mitochondrial transcription (Surovtseva et al., 2011). Certain 28S subunit poly-peptides not well represented at the nucleoid may join assembly at later stages.
SILAC permitted us to identify newly synthesized proteins enriched in the nucleoid fraction in comparison with the ribosome-enriched fraction. Our study was designed to compare proteins well represented in both the nucleoid and ribosome fractions. Therefore, proteins largely confined to the nucleoid (Table 1), as well as any ribosomal proteins that are not abundant in the nucleoid, do not figure in the ratiometric analysis. We found 111 proteins in both the nucleoid- and ribosome-enriched frac-tions with sufficient coverage to permit calculation of the relative content of heavy and light peptides using MaxQuant analysis of MS spectra (Cox and Mann, 2008). An example of the MS/MS data obtained for one peptide derived from MRPL28 is shown in Fig-ure 5A, and additional examples are shown inFigure S3. As in conventional peptide MS experiments, more than one molecular ion is detected for each peptide in this analysis, due to the natural abundance of13C, resulting in a nested set of molecular ions identifying each peptide. For MRPL28, as for other 39S subunit polypeptides, the striking result is that the intensity of the13 C-labeled peaks generated during the pulse (# in Figure 5A) is slightly greater than that of the12C-labeled peaks attributable to the preexisting protein (* inFigure 5A), but only for the protein observed in the nucleoid fraction. Averaging over all peptides observed for MRPL28, we found the H/L ratio was 6.3-fold higher in the nucleoid fraction than in the ribosome-enriched fraction,
consistent with the hypothesis that newly synthesized MRPL28 accumulates for some duration in the vicinity of the nucleoid. In a brief pulse, any newly synthesized MRPL28 incorporated into a ribosome that completes assembly and dissociates from the nucleoid is diluted by the large pool of ‘‘light’’ protein in the fraction enriched in preexisting ribosomes.
Unsupervised ranking of all 111 proteins identified in our anal-ysis according to their nucleoid/ribosome labeling ratios re-vealed 31 MRPs among those proteins with the highest nucleoid enrichment of newly synthesized polypeptides (Figure 5B). Of the top 14 ranked proteins with the highest nucleoid enrichment factors, 13 were 39S subunit proteins. The 17 28S subunit pro-teins identified had significantly lower H/L ratios, but this was still over 1.73 times the average ratio of the nonribosomal proteins. The inset inFigure 5B shows that the means and 95% confi-dence limits of the distributions of 39S, 28S, and nonribosomal proteins define three distinct distributions. These polypeptides showing a high abundance of newly synthesized protein in the nucleoid fraction most likely bind nascent mtRNA at an early stage in ribosome assembly, making them candidates for pri-mary ribosomal proteins that bind early during assembly, in contrast to secondary or tertiary proteins that join the ribosome assembly process later.
To further test this model, we conducted two additional inde-pendent labeling experiments. First, we conducted pulse-chase experiments in which cells pulse labeled for 6 hr in13C-arginine and -lysine were switched back to normal medium for a 4 hr chase. With this labeling scheme, many newly synthesized ‘‘heavy’’ proteins found at the nucleoid immediately after a pulse would be expected to be in ribosomes that complete assembly during the chase. Simultaneous MaxQuant analysis was performed on eight data sets from the nucleoid and ribosome fractions from two pulse-chase experiments along with two pulse-labeling experiments. The H/L density ratios observed for the 111 proteins represented by multiple peptides in both the nucleoid- and ribosome-enriched fractions in at least three of the four experiments are shown in Table S2
and in the heatmap inFigure 6A. We found that the density labeling of the 14 39S subunit proteins decreased by over 60% during the chase, while that of the 17 small subunit proteins decreased by about 33%. In contrast, no systematic ratio changes were observed for the 80 nonribosomal proteins (chase/pulse ratio average 0.97). The 39S and 28S subunit polypeptides generally cluster in distinct patterns in this pre-sentation, since the specific activity of the 39S subunit proteins, which were more heavily labeled during the pulse, drops off more dramatically during the chase.
As a second test of our model, we sought to determine whether the accumulation of nascent MRPs at nucleoids was dependent on active mitochondrial transcription. In the experi-ment shown inFigure 6B, we pulse labeled cells for only 3 hr in the presence or absence of 20-C-methyladenosine, a specific inhibitor of mitochondrial transcription (Arnold et al., 2012). Con-trol experiments confirmed that this compound inhibited mito-chondrial RNA synthesis as effectively as 400 ng/ml ethidium bromide in our hands. Analysis of nucleoid and ribosomal fractions by LC-MS/MS provided ratiometric labeling data on 14 39S subunit proteins, 19 28S subunit proteins, and over 100 nonribosomal proteins. In most cases, the MRPs observed
with this shorter pulse were the same proteins well represented at the nucleoid after a 6 hr pulse. When transcription was in-hibited, the H/L ratios were decreased by 39% and 18% for the 39S and 28S polypeptides, respectively, but were not signif-icantly different for the nonribosomal proteins (Figure 6B). These results are consistent with our model, showing that a subset of ribosomal proteins interacts with nascent rRNA at the nucleoid (Figure 6C).
Not all mitochondrial ribosomal proteins exhibit high nucleoid enrichment scores. For those ribosomal proteins not well repre-sented in the nucleoid fraction, the newly synthesized copies are immediately diluted among bulk ribosomal proteins in our experiments. These proteins may be incorporated into mito-chondrial ribosomes at later stages in assembly. These second-ary steps in ribosome assembly may occur near the nucleoid, but late assembly intermediates have a decreasing probability of maintaining sufficient physical linkage to mtDNA to survive our handling procedures. To test this model further, we tagged four additional MRPs with the Eos fluorescent protein tag to interrogate the location of newly synthesized fusion proteins following brief (7 hr) induction of each protein. Three apparent primary MRP constructs, MRPL13-Eos, MRPLS17-Eos, and MRPS15-Eos, were found in punctate foci near nucleoids, while MRPL10-Eos, a fusion to a secondary MRP that is known to reside in the periphery of the large subunit (Sharma et al., 2003), was found delocalized within the mitochondrial matrix (Figure S4).
DISCUSSION
Nucleoids as Centers for Mitochondrial Biogenesis Nucleoids are substructures within mitochondria where mtDNA genomes are replicated and transcribed, since these processes must occur on the DNA template. Nucleoids are tightly packed in structures smaller than the resolution limit of a conventional light microscope (Brown et al., 2011; Iborra et al., 2004; Kukat et al., 2011). The size of a single nucleoid is sufficient to nearly fill the space of a single infolding of the mitochondrial inner membrane (Kopek et al., 2012). The major components of nucleoids that lack enzymatic activity are the DNA binding proteins TFAM and mtSSB. Other proteins associate with nucleoids in order to participate in replication and transcription (Tables 1 and S1). We now extend this paradigm to show that factors involved in mtRNA processing and initial steps in assembly of mitoribo-somes are also closely associated with nucleoids. Due to steric constraints, some of the participants in mtRNA processing, ribo-some assembly, and turnover of unused nascent mtRNA apparently cannot be accommodated in the tight confines of a 100 nm diameter nucleoid core structure. This may help explain the apparent slight spatial separation between nucleoids and processing granules or degradosomes (Antonicka et al., 2013; Borowski et al., 2013; Jourdain et al., 2013). We propose that one can consider many, perhaps most, processing bodies as dynamic physical extensions of nucleoids. Several proteins involved in RNA processing and degradation, such as SUV3, PNPT1, and GRSF1, are well represented in nucleoid prepara-tions (Tables 1 and S1). We have not efficiently separated nucleoids and processing granules using biochemical methods. When mitochondrial membranes are dissolved using nonionic
Figure 6. Large and Small Subunit MRPs Found in the Nucleoid Fraction Show Reduced Labeling after a Chase or when Transcription Is Inhibited, Consistent with Assembly into Nascent Ribosomes at the Nucleoid
(A) Effects of chase incubation following the pulse label. Heatmap showing nucleoid:ribosome H/L label ratios for 111 proteins after pulse labeling and pulse-chase labeling. High ratiosR4 are shown in red, lower ratios%1 are shown in blue.
(B) Effects of transcription inhibition. HeLa cells were incubated for 3 hr in either the presence or absence of 100mM 20-C-methyladenosine as an inhibitor of mtRNA polymerase during pulse labeling with13
C-lysine/arginine. Nucleoid-enriched and ribosome-enriched fractions were processed to determine H/L peptide ratios. The boxplot shows median and quartile levels for the distributions as analyzed by ANOVA.
(C) mtRNA polymerase (R) is shown engaged in transcription at a mtDNA nucleoid with nascent RNA extending from the nucleoid core. RNase P (P) and various RNA binding proteins and newly synthesized ribosomal proteins may encounter the nascent RNA while transcription continues. Further maturation of the 12S rRNA involves cleavage by RNase P and ELAC2 (E) and the binding of secondary ribosomal proteins as the rRNA folds into a mature secondary and tertiary structure.
detergents, RNA complexes no longer covalently linked to mtDNA can be separated using gentle sedimentation methods, as shown inFigure 2C. Incompletely processed mtRNAs may remain largely confined to the vicinity of the nucleoid in vivo.
Early observation of very long mtRNAs (Aloni and Attardi, 1971) raised the possibility that primary transcripts must be released from mtDNA to be delivered to distant sites to initiate processing. The tRNA punctuation model for mtRNA suggested the alternative possibility that processing of primary transcripts might begin even while transcription of an individual genome is still underway. While this concept was implied by a cartoon illus-trating the tRNA punctuation model (Figure 4 of Ojala et al., 1981), this was not established at the time. This model stimulated a search for mtRNase P, also unknown at the time, as an early participant in mtRNA processing. We detected all three subunits of mtRNase P and ELAC2 enriched in nucleoid preparations (Tables 1,S1, andS2) and localized all four of these tRNA cleav-age proteins at or near nucleoids using fluorescent reporter fusions (Figures 1andS1).
Many other RNA binding proteins are not localized exclusively to nucleoids. For example, LRPPRC is abundant in nucleoid preparations but is not confined to the nucleoid. This protein participates with SLIRP in binding and stabilizing mitochondrial mRNAs and facilitating polyadenylation (Ruzzenente et al., 2012; Chujo et al., 2012; Gohil et al., 2010; Sasarman et al., 2010). LRPPRC/SLIRP is distributed essentially throughout the mitochondrial matrix. Our data show that both LRPPRC and SLIRP occur with essentially equal H/L isotope ratios in both the nucleoid- and ribosome-enriched fractions, and mass spectrometry intensity data show that only about 40% of each protein is associated with nucleoids (Table S3). This is the result expected for proteins that move readily in and out of the nucleoid domain. By stabilizing mRNAs against turnover, LRPPRC/SLIRP may assist in the exchange of genetic information between mitochondria to protect against mutations that may arise in indi-vidual mtDNA genomes. This sort of exchange is necessary to permit the genetic complementation documented in a number of studies, beginning with early somatic cell hybridization exper-iments showing translation of mRNAs encoded in one genome on ribosomes generated from another (Oliver and Wallace, 1982).
Mitochondrial Ribosome Assembly Begins at Nucleoids Mitochondrial ribosomes have been difficult to study, in part because they are not highly abundant. A HeLa cell contains only about 34,000 mitoribosomes, but about 6 million cyto-plasmic ribosomes (Attardi and Schatz, 1988). Understanding how approximately 80 nuclear-encoded MRPs are indepen-dently imported into mitochondria and assembled with nascent mitochondrial rRNA to build a ribosome has been an elusive goal. The assembly process may be relatively slow and may involve a significant amount of wastage or turnover of polypep-tides that arrive at assembly sites when they are not specifically required.
Some insight into mitoribosome assembly may be gleaned from a comparison with bacterial ribosome assembly, since nearly half of mitochondrial ribosomal proteins have recogniz-able bacterial homologs (Koc et al., 2010). Although bacterial ribosomal proteins are able to self-assemble with rRNA
in vitro to form functional ribosomes, this process is more effi-cient in vivo (Shajani et al., 2011; Woodson, 2008). Several poly-peptides of the bacterial small ribosomal subunit are considered to bind as primary RNA-interacting proteins, while others asso-ciate as secondary or tertiary proteins dependent on prior bind-ing of the primary proteins. However, these prokaryotic maps are only a rough guide to potential mammalian mitochondrial ribosome assembly, since the mitochondrial rRNAs have under-gone extensive deletion and new proteins with no clear rela-tionship to prokaryotic ribosomal proteins have been added. Mutations in some of the mitochondrial ribosomal proteins lack-ing bacterial homologs, such as MRPS22, a protein we found enriched at nucleoids (Figure 6A), can lead to fatal disorders (Smits et al., 2011).
Our SILAC experiments provided data on over 75 MRPs. We focused on those proteins that were observed in both the nucleoid- and ribosome-enriched fractions in at least three of four experiments. The incomplete data obtained for those MRPs excluded from the main analysis indicate that they did not show high H/L peptide ratios, which is consistent with their assignment as secondary proteins. Some of these proteins may be amenable to additional analysis in future studies. We consider that the data reported here on 31 nucleoid-associated MRPs provides robust support for the conclusion that these proteins tend to be assembled into mitoribosomes at or near the nucleoid. MRPs considered to have bacterial homologs (28S proteins numbered 21 and below, 39S proteins numbered 36 and below; Koc et al., 2010) are not overly represented in our list of candidate primary ribosomal proteins. This may be expected, since, by definition, not all prokaryotic ribosomal pro-teins bind as primary propro-teins. In fact, many primary bacterial RPs do not appear to have mitochondrial homologs, consistent with the extensive deletion of bacterial rRNA segments in the mitochondrial rRNAs. If we consider only the set of bacterial pri-mary RPs that have mitochondrial homologs (S17*, S15*, S7*, S18*, and S9*; L20, L4*, L13*, L2, L1*, L9, L3*), we find that the majority of these MRPs (those indicated with asterisks) behave as primary binding proteins in our SILAC study. We obtained extensive coverage (33%–45%) for MRPL2, L9, and L20 in the ribosome fraction, but these were absent or underrepresented in the nucleoid fraction, implying that they participate in later stages of assembly.
We were surprised to find that the relative intensity of the13C label for nucleoid-associated 39S subunits is significantly higher than that for 28S subunit proteins inFigure 5B. Since this statistic involves calculation of an H/L ratio, we reasoned that this could reflect the presence of a larger steady-state pool of preexisting 28S subunits at the nucleoid, a larger denominator in the ratio. By accessing spectral count and signal intensity data in the Max-Quant analysis of our MS data for those MRPs represented at the nucleoid, we estimated the pool of each protein observed at the nucleoid relative to the ribosome-enriched fraction (Figure 4B). We found that only about 3.7% of the pool of 39S proteins was observed in the nucleoid fraction, while approximately 15% of the 28S protein signal intensity was found at the nucleoid. Thus, the smaller pool of 39S subunits observed at the nucleoid is more intensely labeled with13C or has a higher specific activity than the 28S MRPs, possibly because the pool of 39S MRP pro-teins at the nucleoid turns over more rapidly, as if these propro-teins
spend less time engaged in assembly than the 28S MRPs. This in turn suggests that the 12S rRNA and associated proteins may not be released from the vicinity of the nucleoid as quickly as the nascent large subunit. This may be expected, since the 12S rRNA is transcribed by mtRNA polymerase before the 16S rRNA so that it is available to interact with nascent ribosomal proteins at an earlier stage. The facile detection of a long RNA termed RNA 4, containing both 12S and 16S rRNA, indicates that this precursor has a relatively long half-life, as if cleavages surrounding tRNAValmay not occur rapidly, providing time for the RNA to bind proteins (Ojala et al., 1981). Our results may reflect a mechanism for coordinate quality control of mitochon-drial ribosome biogenesis at the nucleoid, possibly involving fac-tors such as mTERF3, which is significantly nucleoid associated (Table 1) and has been shown to have a role in ribosome biogen-esis (Wredenberg et al., 2013). rRNA modification events likely occurring at the nucleoid also appear to play a role in quality control of mitoribosome assembly (Metodiev et al., 2009), with important human health implications (Raimundo et al., 2012). A recent analysis of lifespan in the inbred mouse BXD panel has revealed a number of MRPs as primary determinants of longevity (Houtkooper et al., 2013). The majority of MRPs included in this study are among the nucleoid-enriched proteins we identified, suggesting that properly coordinated mitoribosome assembly may be a factor in the mitonuclear protein imbalance proposed to play a role in aging. Further tests of this model are beyond the scope of our study. We conclude that SILAC pulse-labeling experiments provide robust support for our main hypothesis, that mitoribosome assembly begins at the nucleoid. This work has established SILAC pulse labeling as a valuable approach for further studies of the kinetics and mechanism of mitoribo-some assembly.
EXPERIMENTAL PROCEDURES
Plasmids, Protein Expression, and Microscopy
cDNA clones for mouse MRPP1, MRPP2, MRPP3, ELAC2 (RNase ZL),
MRPL13, MRPS15, MRPS17, and MRPL10 were generated using first-strand cDNA (Clontech) as template and using High-Fidelity Phusion Reverse Tran-scriptase (New England Biolabs). Primers added AscI restriction sites at the 50end and NotI sites at the 30end. Sequences were first cloned using TOPO cloning in pCRII vector (Life Technologies), sequenced to confirm the con-struction, and transferred as AscI/NotI fragments to pGS-Eos vector (Brown et al., 2010). Expression constructs were transfected into 3T3-switch cells, and stably transformed cell lines were generated by selecting for Zeocin resis-tance. For microscopy, fusion protein expression was induced by addition of 100 pM mifepristone for 7 hr. Where indicated, cells were stained with 50 nM MitoTracker Red CM-H2XRos for 90 min prior to fixation and imaging. Zeocin, mifepristone, and MitoTracker were obtained from Life Technologies. Additional imaging methods are described inSupplemental Experimental Procedures.
Quantitative Real-Time PCR
RNA was prepared from total cells or purified mitochondria or from glycerol gradient fractions using TRIzol (Life Technologies). RNA samples were treated with TURBO DNase (Ambion, Life Technologies) to remove contaminating DNA and were converted to first-strand cDNA using a High-Capacity cDNA Reverse Transcription Kit from Applied Biosystems (Life Technologies). Quan-titative real-time PCR was done using SYBR Green detection in a StepOne instrument (Applied Biosystems; Life Technologies) with theDDCt method (Schmittgen and Livak, 2008). Primer pairs are indicated inSupplemental Experimental Proceduresand were validated in reactions using mtDNA puri-fied from HeLa cells as template.
Protein Labeling
For stable isotope labeling, HeLa cells grown in monolayer culture in Dulbec-co’s modified Eagle’s medium (DMEM) were rinsed with Hank’s balanced salt solution (HBSS) for 5 min at 37C prior to addition of DMEM F12 medium con-taining 10% dialyzed fetal bovine serum (FBS) lacking12
C-arginine/lysine sup-plemented with 30 mg/l13
C-arginine and 50 mg/l13
C-lysine (Pierce SILAC kits, Thermo Scientific). For chase experiments, the13
C medium was removed, cells were rinsed with HBSS and then further incubated in HBSS at 37C for 5 min, rinsed with normal DMEM/10% FBS with12
C amino acids, and finally incubated in the same DMEM/10% FBS for 4 hr. Where indicated, cells were incubated with 100mg/ml 20-C-methyladenosine (Santa Cruz) to inhibit mitochondrial transcription (Arnold et al., 2012). A typical preparation used 6–10 15 cm tissue culture plates and began with 83107
to 103107
cells. Cell fractionation in mannitol-sucrose-HEPES (MSH) buffer (pH 8.0) was con-ducted as described (Lee et al., 2013). Briefly, cell homogenates were centri-fuged twice at 1,2003gto remove nuclei, and mitochondria were pelleted at 15,0003gfor 15 min, washed with MSH buffer containing 1 M KCl, treated with DNase I andSerratia marcescansendonuclease, and purified using sucrose gradients. Mitochondrial lysis with 1.5% Triton X-100 and preparation of nucleoid- and ribosome-enriched fractions were performed as described (Lee et al., 2013), except that glycerol and iodixanol gradients used 113 60 mm Beckman SW 60Ti tubes to minimize volumes. Nucleoids and ribo-somes were routinely identified in the final fractions using nucleic acid fluores-cence and immunoblotting with antibodies directed against TFAM and MRPL13. Methods for protein analysis by LC-MS/MS are inSupplemental Information.
SUPPLEMENTAL INFORMATION
Supplemental Information includes Supplemental Experimental Procedures, four figures, and three tables and can be found with this article online at http://dx.doi.org/10.1016/j.cmet.2014.03.013.
ACKNOWLEDGMENTS
This work was supported by a Senior Scholar Award from the Ellison Medical Foundation to D.F.B. Received: October 8, 2013 Revised: December 17, 2013 Accepted: February 6, 2014 Published: April 1, 2014 REFERENCES
Aloni, Y., and Attardi, G. (1971). Symmetricalin vivotranscription of mitochon-drial DNA in HeLa cells. Proc. Natl. Acad. Sci. USA68, 1757–1761. Anderson, S., Bankier, A.T., Barrell, B.G., de Bruijn, M.H.L., Coulson, A.R., Drouin, J., Eperon, I.C., Nierlich, D.P., Roe, B.A., Sanger, F., et al. (1981). Sequence and organization of the human mitochondrial genome. Nature 290, 457–465.
Antonicka, H., Sasarman, F., Nishimura, T., Paupe, V., and Shoubridge, E.A. (2013). The mitochondrial RNA-binding protein GRSF1 localizes to RNA gran-ules and is required for posttranscriptional mitochondrial gene expression. Cell Metab.17, 386–398.
Arnold, J.J., Sharma, S.D., Feng, J.Y., Ray, A.S., Smidansky, E.D., Kireeva, M.L., Cho, A., Perry, J., Vela, J.E., Park, Y., et al. (2012). Sensitivity of mitochondrial transcription and resistance of RNA polymerase II dependent nuclear transcription to antiviral ribonucleosides. PLoS Pathog.8, e1003030. Attardi, G., and Schatz, G. (1988). Biogenesis of mitochondria. Annu. Rev. Cell Biol.4, 289–333.
Barat, M., Rickwood, D., Dufresne, C., and Mounolou, J.-C. (1985). Characterization of DNA-protein complexes from the mitochondria of Xenopus laevis oocytes. Exp. Cell Res.157, 207–217.
Bogenhagen, D.F. (2012). Mitochondrial DNA nucleoid structure. Biochim. Biophys. Acta1819, 914–920.
Bogenhagen, D.F., Wang, Y., Shen, E.L., and Kobayashi, R. (2003). Protein components of mitochondrial DNA nucleoids in higher eukaryotes. Mol. Cell. Proteomics2, 1205–1216.
Bogenhagen, D.F., Rousseau, D., and Burke, S. (2008). The layered structure of human mitochondrial DNA nucleoids. J. Biol. Chem.283, 3665–3675. Boisvert, F.-M., Ahmad, Y., Gierlinski, M., Charrie`re, F., Lamont, D., Scott, M., Barton, G., and Lamond, A.I. (2012). A quantitative spatial proteomics analysis of proteome turnover in human cells. Mol. Cell. Proteomics11, 011429. Borowski, L.S., Dziembowski, A., Hejnowicz, M.S., Stepien, P.P., and Szczesny, R.J. (2013). Human mitochondrial RNA decay mediated by PNPase-hSuv3 complex takes place in distinct foci. Nucleic Acids Res.41, 1223–1240.
Brown, T.A., Fetter, R.D., Tkachuk, A.N., and Clayton, D.A. (2010). Approaches toward super-resolution fluorescence imaging of mitochondrial proteins using PALM. Methods51, 458–463.
Brown, T.A., Tkachuk, A.N., Shtengel, G., Kopek, B.G., Bogenhagen, D.F., Hess, H.F., and Clayton, D.A. (2011). Superresolution fluorescence imaging of mitochondrial nucleoids reveals their spatial range, limits, and membrane interaction. Mol. Cell. Biol.31, 4994–5010.
Brzezniak, L.K., Bijata, M., Szczesny, R.J., and Stepien, P.P. (2011). Involvement of human ELAC2 gene product in 30end processing of mitochon-drial tRNAs. RNA Biol.8, 616–626.
Chen, S.S., and Williamson, J.R. (2013). Characterization of the ribosome biogenesis landscape in E. coli using quantitative mass spectrometry. J. Mol. Biol.425, 767–779.
Chujo, T., Ohira, T., Sakaguchi, Y., Goshima, N., Nomura, N., Nagao, A., and Suzuki, T. (2012). LRPPRC/SLIRP suppresses PNPase-mediated mRNA decay and promotes polyadenylation in human mitochondria. Nucleic Acids Res.40, 8033–8047.
Cox, J., and Mann, M. (2008). MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide pro-tein quantification. Nat. Biotechnol.26, 1367–1372.
De Silva, D., Fontanesi, F., and Barrientos, A. (2013). The DEAD box protein Mrh4 functions in the assembly of the mitochondrial large ribosomal subunit. Cell Metab.18, 712–725.
Falkenberg, M., Larsson, N.-G., and Gustafsson, C.M. (2007). DNA replication and transcription in mammalian mitochondria. Annu. Rev. Biochem. 76, 679–699.
Fatica, A., and Tollervey, D. (2002). Making ribosomes. Curr. Opin. Cell Biol. 14, 313–318.
Gohil, V.M., Nilsson, R., Belcher-Timme, C.A., Luo, B., Root, D.E., and Mootha, V.K. (2010). Mitochondrial and nuclear genomic responses to loss of LRPPRC expression. J. Biol. Chem.285, 13742–13747.
He, J., Cooper, H.M., Reyes, A., Di Re, M., Sembongi, H., Litwin, T.R., Gao, J., Neuman, K.C., Fearnley, I.M., Spinazzola, A., et al. (2012). Mitochondrial nucleoid interacting proteins support mitochondrial protein synthesis. Nucleic Acids Res.40, 6109–6121.
Holzmann, J., Frank, P., Lo¨ffler, E., Bennett, K.L., Gerner, C., and Rossmanith, W. (2008). RNase P without RNA: identification and functional reconstitution of the human mitochondrial tRNA processing enzyme. Cell 135, 462–474.
Houtkooper, R.H., Mouchiroud, L., Ryu, D., Moullan, N., Katsyuba, E., Knott, G., Williams, R.W., and Auwerx, J. (2013). Mitonuclear protein imbalance as a conserved longevity mechanism. Nature497, 451–457.
Iborra, F.J., Kimura, H., and Cook, P.R. (2004). The functional organization of mitochondrial genomes in human cells. BMC Biol.2, 9.
Jourdain, A.A., Koppen, M., Wydro, M., Rodley, C.D., Lightowlers, R.N., Chrzanowska-Lightowlers, Z.M., and Martinou, J.C. (2013). GRSF1 regu-lates RNA processing in mitochondrial RNA granules. Cell Metab. 17, 399–410.
Kellems, R.E., Allison, V.F., and Butow, R.A. (1975). Cytoplasmic type 80S ribosomes associated with yeast mitochondria. IV. Attachment of ribosomes to the outer membrane of isolated mitochondria. J. Cell Biol.65, 1–14.
Koc, E.C., Haque, M.E., and Spremulli, L.L. (2010). Current Views of the Structure of the Mammalian Mitochondrial Ribosome. Isr. J. Chem. 50, 45–59.
Kopek, B.G., Shtengel, G., Xu, C.S., Clayton, D.A., and Hess, H.F. (2012). Correlative 3D superresolution fluorescence and electron microscopy reveal the relationship of mitochondrial nucleoids to membranes. Proc. Natl. Acad. Sci. USA109, 6136–6141.
Kressler, D., Hurt, E., and Bassler, J. (2010). Driving ribosome assembly. Biochim. Biophys. Acta1803, 673–683.
Kukat, C., Wurm, C.A., Spa˚hr, H., Falkenberg, M., Larsson, N.-G., and Jakobs, S. (2011). Super-resolution microscopy reveals that mammalian mitochondrial nucleoids have a uniform size and frequently contain a single copy of mtDNA. Proc. Natl. Acad. Sci. USA108, 13534–13539.
Lane, N., and Martin, W. (2010). The energetics of genome complexity. Nature 467, 929–934.
Lee, K.W., Okot-Kotber, C., LaComb, J.F., and Bogenhagen, D.F. (2013). Mitochondrial ribosomal RNA (rRNA) methyltransferase family members are positioned to modify nascent rRNA in foci near the mitochondrial DNA nucleoid. J. Biol. Chem.288, 31386–31399.
Liu, M., and Spremulli, L. (2000). Interaction of mammalian mitochondrial ribo-somes with the inner membrane. J. Biol. Chem.275, 29400–29406. Metodiev, M.D., Lesko, N., Park, C.B., Ca´mara, Y., Shi, Y., Wibom, R., Hultenby, K., Gustafsson, C.M., and Larsson, N.-G. (2009). Methylation of 12S rRNA is necessary for in vivo stability of the small subunit of the mamma-lian mitochondrial ribosome. Cell Metab.9, 386–397.
Mimaki, M., Wang, X., McKenzie, M., Thorburn, D.R., and Ryan, M.T. (2012). Understanding mitochondrial complex I assembly in health and disease. Biochim. Biophys. Acta1817, 851–862.
Ojala, D., Montoya, J., and Attardi, G. (1981). tRNA punctuation model of RNA processing in human mitochondria. Nature290, 470–474.
Oliver, N.A., and Wallace, D.C. (1982). Assignment of two mitochondrially synthesized polypeptides to human mitochondrial DNA and their use in the study of intracellular mitochondrial interaction. Mol. Cell. Biol. 2, 30–41.
Ong, S.-E., Blagoev, B., Kratchmarova, I., Kristensen, D.B., Steen, H., Pandey, A., and Mann, M. (2002). Stable isotope labeling by amino acids in cell culture, SILAC, as a simple and accurate approach to expression proteomics. Mol. Cell. Proteomics1, 376–386.
Rackham, O., Mercer, T.R., and Filipovska, A. (2012). The human mitochon-drial transcriptome and the RNA-binding proteins that regulate its expression. Wiley Interdiscip Rev RNA3, 675–695.
Raimundo, N., Song, L., Shutt, T.E., McKay, S.E., Cotney, J., Guan, M.-X., Gilliland, T.C., Hohuan, D., Santos-Sacchi, J., and Shadel, G.S. (2012). Mitochondrial stress engages E2F1 apoptotic signaling to cause deafness. Cell148, 716–726.
Rorbach, J., and Minczuk, M. (2012). The post-transcriptional life of mamma-lian mitochondrial RNA. Biochem. J.444, 357–373.
Rorbach, J., Richter, R., Wessels, H.J., Wydro, M., Pekalski, M., Farhoud, M., Ku¨hl, I., Gaisne, M., Bonnefoy, N., Smeitink, J.A., et al. (2008). The human mito-chondrial ribosome recycling factor is essential for cell viability. Nucleic Acids Res.36, 5787–5799.
Rossmanith, W. (2012). Of P and Z: mitochondrial tRNA processing enzymes. Biochim. Biophys. Acta1819, 1017–1026.
Ruzzenente, B., Metodiev, M.D., Wredenberg, A., Bratic, A., Park, C.B., Ca´mara, Y., Milenkovic, D., Zickermann, V., Wibom, R., Hultenby, K., et al. (2012). LRPPRC is necessary for polyadenylation and coordination of transla-tion of mitochondrial mRNAs. EMBO J.31, 443–456.
Sasarman, F., Brunel-Guitton, C., Antonicka, H., Wai, T., and Shoubridge, E.A.; LSFC Consortium (2010). LRPPRC and SLIRP interact in a ribonucleoprotein complex that regulates posttranscriptional gene expression in mitochondria. Mol. Biol. Cell21, 1315–1323.
Schmittgen, T.D., and Livak, K.J. (2008). Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc.3, 1101–1108.
Shajani, Z., Sykes, M.T., and Williamson, J.R. (2011). Assembly of bacterial ribosomes. Annu. Rev. Biochem.80, 501–526.
Sharma, M.R., Koc, E.C., Datta, P.P., Booth, T.M., Spremulli, L.L., and Agrawal, R.K. (2003). Structure of the mammalian mitochondrial ribosome reveals an expanded functional role for its component proteins. Cell115, 97–108. Smits, P., Saada, A., Wortmann, S.B., Heister, A.J., Brink, M., Pfundt, R., Miller, C., Haas, D., Hantschmann, R., Rodenburg, R.J.T., et al. (2011). Mutation in mitochondrial ribosomal protein MRPS22 leads to Cornelia de Lange-like phenotype, brain abnormalities and hypertrophic cardiomyopathy. Eur. J. Hum. Genet.19, 394–399.
Surovtseva, Y.V., Shutt, T.E., Cotney, J., Cimen, H., Chen, S.Y., Koc, E.C., and Shadel, G.S. (2011). Mitochondrial ribosomal protein L12 selectively associ-ates with human mitochondrial RNA polymerase to activate transcription. Proc. Natl. Acad. Sci. USA108, 17921–17926.
Talkington, M.W.T., Siuzdak, G., and Williamson, J.R. (2005). An assembly landscape for the 30S ribosomal subunit. Nature438, 628–632.
Vogel, F., Bornho¨vd, C., Neupert, W., and Reichert, A.S. (2006). Dynamic sub-compartmentalization of the mitochondrial inner membrane. J. Cell Biol.175, 237–247.
Wang, Y., and Bogenhagen, D.F. (2006). Human mitochondrial DNA nucleoids are linked to protein folding machinery and metabolic enzymes at the mito-chondrial inner membrane. J. Biol. Chem.281, 25791–25802.
Woodson, S.A. (2008). RNA folding and ribosome assembly. Curr. Opin. Chem. Biol.12, 667–673.
Wredenberg, A., Lagouge, M., Bratic, A., Metodiev, M.D., Spa˚hr, H., Mourier, A., Freyer, C., Ruzzenente, B., Tain, L., Gro¨nke, S., et al. (2013). MTERF3 reg-ulates mitochondrial ribosome biogenesis in invertebrates and mammals. PLoS Genet.9, e1003178.