Magnetic Sense of Juvenile Gulf Flounder
(
Paralichthys albigutta
)
By
EmmaLi G. Tsai
Senior Honors Thesis
Environment, Ecology, and Energy Program
University of North Carolina at Chapel Hill
(26 November 2019)
Approved:
Dr. Joel Fodrie, Thesis Advisor
ABSTRACT... 3
1. INTRODUCTION... 4
2. METHODOLOGY... 7
2.1 Fish Capture and Husbandry... 7
2.2 Magnetic Coil Structure... 8
2.3 Experimental Conditions... 9
2.4 Experimental Treatments... 10
2.4.1 Magnetic Compass Experiments... 10
2.4.2 Magnetic Map Experiments... 11
2.5 Experimental Procedure... 13
2.6 Data Analysis... 13
3. RESULTS... 15
3.1 Magnetic Compass Results... 15
3.2 Magnetic Map Results... 16
4. DISCUSSION... 17
4.1 Magnetic Compass Experiments... 17
4.2 Magnetic Map Experiments... 18
5. FUTURE STEPS... 19
6. APPLICATIONS & IMPLICATIONS... 21
ABSTRACT
Diverse marine taxa use Earth's magnetic field for navigation. This ability, known as
magnetic navigation, has classically been grouped into two main types – the magnetic “compass”
and “map” sense. Organisms possessing a magnetic compass sense are able to use the Earth’s
magnetic field to gain directional information, similar to a human compass. In contrast, an
organism possessing a magnetic map sense uses the Earth’s magnetic field to determine their
position relative to a goal, similar to a human GPS. We assessed for the presence of both a
magnetic compass and map sense in juvenile Gulf Flounder (Paralichthys albigutta), a species of
migratory flatfish found in coastal waters spanning from North Carolina to the Gulf of Mexico.
In both experiments, a magnetic coil system was used to produce different magnetic field
treatments. To test for a magnetic compass sense, flounder were tested in one of two magnetic
treatments: (1) the ambient magnetic field of the test site; (2) the same magnetic field but with
reversed polarity, where the direction of magnetic north was reversed. To search for the presence
of a magnetic map sense, we tested flounder in one of two magnetic fields that exist at two
locations: (1) north of the test site (near New Jersey); and (2) south of the test site (near
Jamaica). Flounder consistently oriented towards a magnetic direction in our compass
experiments, which suggests the presence of a compass sense. In the magnetic map experiments,
flounder exhibited orientations that were significantly different between northern and southern
treatments, providing evidence for a magnetic map sense. Considering past magnetic navigation
research has mainly focused on long-distance migrants, the discovery of a both a magnetic
compass and map sense in flatfish brings to light how these senses could be useful for organisms
undergoing relatively shorter migrations between nearshore and offshore environments.
1. INTRODUCTION
Many fish species move between ocean habitats that are critical for different stages of
their life history (Aas et al., 2011; Dingle, 2014). In fish, an abundance of these history strategies
involve offshore larval dispersal, a pelagic stage, and subsequent settlement in a coastal nursery
habitat where fish continue their development (Dingle, 2014). As knowledge behind fish
migration has developed, past biotelemetry and stable isotope analysis has discovered these
movements can actually cover significant distances across ocean environments (Bultel et al.,
2014; Madigan et al., 2014). During these long-distance migrations, fish utilize a suite of
navigational systems to successfully arrive at habitats essential for growth and reproduction
(Gould, 1998). The systems must be reliable to account for directional errors that inevitably arise
when navigating an ever-changing ocean environment (Lohmann, 2008). However, obtaining
consistent navigational information is a challenge in the open-ocean environment; a habitat
largely devoid of visual landmarks. How fish are able to achieve these long-distance migrations
can be elucidated by understanding the sensory mechanisms guiding these movements.
Previous research suggests that fish use an amalgamation of acoustic, visual,
hydrodynamic, olfactory, and geomagnetic cues to navigate the ocean environment (Radford et
al., 2010; Gardiner & Atema, 2014; Groves et al., 1968; Lohmann 2008). In such an
environment, information gained from Earth’s magnetic field can be particularly beneficial to
migrating organisms. It is present everywhere on the planet – from the deepest ocean trenches to
the highest mountains, and largely undisturbed by extreme weather events (Lohmann et al.,
2004). Using the Earth’s magnetic field for navigational information is known as “magnetic
navigation”, and is an ability that has been discovered in diverse taxa ranging from birds,
Vácha et al., 2009; Kimchi & Terkel, 2001; Ernst & Lohmann, 2018; Naisbett-Jones et al., 2017;
Putnam et al., 2014; Lohmann et al., 2004). As knowledge behind this ability has developed,
there are two main types of magnetic sense that have been identified by scientists. The first is
known as the “magnetic compass” sense, where an organism is able to use the Earth’s magnetic
field to identify a specific magnetic direction to maintain a constant heading, similar to a human
compass (Lohmann et al., 1993). This ability has been identified in many organisms, such as
lobsters, fish, and mollusks (Ernst & Lohmann, 2018; Bottesch et al., 2016; Cain et al., 2005).
The second is known as a “magnetic map” sense, where an organism can use the Earth’s
magnetic field to determine their position relative to a goal, similar to a human GPS (Wiltschko
& Wiltschko, 2005). The magnetic map sense has been discovered in many long-distance
migrants, such as eels, sea turtles, and salmon (Naisbett-Jones et al., 2017; Lohmann et al., 2004;
Putnam et al., 2017). While our understanding of magnetic navigation has increased substantially
over the years, current research has largely focused on iconic long-distance migrants that exhibit
high fidelity to specific locations. Comparatively little research, however, has been conducted on
fish possessing other life history characteristics.
Fish that do not exhibit high fidelity to specific locations
nonetheless experience navigational obstacles where a magnetic sense
could provide useful information. An example of such a case would be
flatfish (Pleuronectiformes spp.); an order of oceanadromous demersal
fish that face the challenge of navigating the ocean floor. Flatfish
species adopt a relatively common life history strategy that requires
migration of 10s-100s km between inshore and offshore habitats (Gibson et al., 2015). Adult
producing small larvae. These pelagic larvae develop in the water column and undergo an
incredible metamorphosis to become demersal juveniles, where one eye fully migrates to the
other side of the head (Figure 1; McMenamin & Parichy, 2013). During the late spring and early
summer months, juveniles settle in continental nursery habitats to continue their development.
During the offseason, flatfish either migrate offshore or southward to reach warmer waters
(Gibson et al., 2015). While previous telemetry studies with European plaice (Pleuronectes
platessa) have demonstrated that this species of flatfish are able to maintain a constant compass
bearing, to our knowledge no study has researched the magnetic sense in any species of flatfish
(Metcalfe et al., 1993).
Here, we investigated for the presence of both a magnetic compass and map sense in
juvenile Gulf Flounder (Paralichthys albigutta). Gulf flounder is a species of flatfish commonly
found in coastal waters across the southeastern United States, ranging from North Carolina to an
undetermined southern limit around the Gulf of Mexico (Figure 2). For the magnetic compass
experiments that I assisted with, we used two separate treatments: (1) the ambient magnetic field
present at Morehead City, NC; and (2) the ambient magnetic field but with reversed polarity. In
order to assess the presence of a magnetic map sense, I modified magnetic field parameters to
represent magnetic fields at two geographic locations: (1) a magnetic field found north near New
2. METHODOLOGY
2.1 Fish Capture and Husbandry
Gulf Flounder ranging from 8-32 cm were obtained via trawling and seining seagrass
beds in Back Sound, North Carolina (Figure 3; 34.68° N, 76.56° W) during July 3rd-10th of
2018 for the compass experiments, and May 24th - June 6th of 2019 for both the compass and
map experiments. After capture in the field, fish were kept in a non-magnetic aerated container
that was replenished with local water every hour. After transportation, fish were maintained in
elevated non-magnetic outdoor tanks located at the University of North Carolina’s Institute of
Marine Science (UNC-IMS; 34.72° N, 76.75° W). These tanks were elevated to reduce the
magnetic influence of potentially buried objects, and isolated to reduce human-generated
magnetic anomalies. During this time in captivity, the fish were fed small anchovies every other
day and supplied with local water from Bogue Sound that mimicked natural conditions. In the
absence of adverse weather, collected flounder were tested between 24-48 hours after capture to
avoid stress associated with prolonged time spent in captivity. Flounder collected from these
Figure 3. Map showing collection site for Gulf Flounder near Morehead City, North Carolina. Dark blue dotted oval displays the sea grass collection sites located in Bogue Sound, North Carolina. The red star indicates the location
where all experiments took place, UNC-IMS.
2.2 Magnetic Coil Structure
In order to produce the magnetic treatments used in both the compass and map
experiments, we utilized a Merritt et al. (1983) 4-Coil design, which is a wooden structure that
supports 6,000 feet of vertically and horizontally wrapped insulated copper wire (Figure 4).
These copper wires were connected to a BK Precision Model 1550 DC Power Supply box and
supplied with constant current in order to produce a constant magnetic field throughout the
duration of our experiments. By running a current through these vertically and horizontally
wrapped wires, the magnetic coil structure can be used to manipulate the ambient magnetic field
present at the UNC-IMS. All of our treatments were based on magnetic parameters produced by
NOAA’s International Geomagnetic Reference Field (IGRF-12) and verified using a FVM400
All fish were tested on the experimental platform located inside of the wooden structure
and within enclosed black-out curtains. Black-curtains were used to remove all celestial cues and
isolate the magnetic response from the flounder. This experimental platform was also centered
inside of the structure where the magnetic field produced was uniform, and lifted to avoid
magnetic disturbance from objects or debris that may be buried under the coil structure.
2.3 Experimental Conditions
Our preliminary studies with Gulf Flounder illustrated higher activity levels at night,
which is consistent with previous experiments on similar flatfish species (Miyazaki et al., 1997).
Therefore, all experiments were conducted at night between 20:50-4:20. Four opaque circular
orientation arenas were placed on the raised magnetic coil platform and enclosed with black-out
curtains (Figure 5). These arenas were non-magnetic and isolated from each other to reduce
disruption resulting from adjacent flounder movements. Each orientation arena was filled at
uniform depth with local water obtained from Bogue Sound, and large black arrows were drawn
on the coil platform to reference magnetic north. In order to film flounder behavior, an infrared
Figure 5. Image of the experimental platform where all experiments were filmed. Each of the four orientation arenas are isolated and filled to a certain depth with water obtained
from Bogue Sound. The black arrows on the platform are directed towards magnetic north
and were used for video analysis.
2.4 Experimental Treatments
2.4.1 Magnetic Compass Experiments
All compass experiments were conducted between July 3rd-10th of 2018 and May 24th
-June 6th of 2019 outside of UNC-IMS. Flounder were tested in two experimental treatments: (1)
the ambient magnetic field located at Morehead City, NC, and (2) a magnetic field with reversed
polarity where magnetic north would be present on the opposite end of the experimental
platform. The act of reversing magnetic north is often used as a method for determining the
presence of a compass sense in previous experiments (Lohmann & Lohmann, 1993; Lohmann et
al., 1995). If flounder possess a magnetic compass sense, we would hypothesize that flounder
experiencing the reversed magnetic north treatment would orient in approximately the opposite
aimed to gather a total sample of roughly 20 flounder for each treatment, which is consistent
with previous magnetic compass experiments (Cresci et al., 2017; Durif et al., 2013).
(a) (b)
Figure 6. Our expected results if Gulf Flounder possess a magnetic compass sense. (a) In the ambient Morehead City field, the flounder orient towards a
specific magnetic direction, such as west. (b) Upon reversal of the field, the
flounder should orient towards approximately the opposite direction, but still
towards the same magnetic direction.
2.4.2 Magnetic Map Experiments
All magnetic map experiments occurred between the June 23rd - August 13th of 2019. For
the magnetic map experiments, I modified both magnetic field intensity and the angle the
magnetic field intersects the surface of the Earth (inclination), to represent magnetic fields of
different geographic locations. I generated magnetic fields that would be found at locations
approximately 600km north and south of our experimental location of North Carolina: (1) a
magnetic field of stronger field intensity and larger inclination angle found north near New
Jersey, and (2) a magnetic field of weaker field intensity and smaller inclination angle found
tested organisms in magnetic fields present at different geographic locations (Naisbett-Jones et
al., 2017; Boles & Lohmann, 2003; Putman et al., 2014). If flounder possess a magnetic map
sense, I hypothesize that flounder would respond to changes in magnetic field intensity and
inclination and exhibit significantly different orientations between experimental treatments. It is
possible they would orient in approximately opposite directions in order to return to the magnetic
field found near their caught location of North Carolina (Figure 7). I aimed to gather a sample
size of approximately 50 fish per treatment, which is consistent with previous magnetic map
experiments (Fischer et al., 1999).
Figure 7. My expected results if Gulf Flounder possess a magnetic map sense. The northern New Jersey treatment has a magnetic field intensity of approximately 51.1 µT and inclination of 65°, and the southern Jamaica treatment
has a magnetic field intensity of approximately 38.4 µT and inclination of 46°. Flounder should sense these changes
treatments. The fish could orient where they would hypothetically return to the magnetic field intensity of 48.6 µT
and inclination of 61° present at their caught location of Morehead City, NC.
2.5 Experimental Procedure
The experimental procedure was similar for both the magnetic compass and magnetic
map experiments. In both experiments, treatments were interspersed throughout the night to
control for temporal variation, such as tidal cycle, which has been shown to previously influence
magnetic orientation in other aquatic species (Cresci et al., 2017). Prior to the start of an
experiment, the four orientation arenas were filled at uniform depth with local water from Bogue
Sound and the particular magnetic treatment was generated. Fish were transported from their
outdoor tank in a non-magnetic bucket to the coil structure, carefully placed into an orientation
arena, and given a 10 - minute acclimation period. Flounder behavior was filmed for a total
period of 2 hours for the compass experiments, and 1.5 hours for the map experiments. The
magnetic compass experiments were filmed for a longer duration than the map experiments to be
consistent with experiments conducted in 2018, which used generally larger and more sedentary
flounder. Between experiments, the orientation arenas were emptied and replenished with local
water, the coil was configured to generate the magnetic field for the other treatment, and four
new fish were placed into each orientation arena. Each fish was only tested once in response to
an experimental treatment and a single individual was either used for the magnetic compass or
map experiments, but not both. After testing, flounder were released into Bogue Sound.
2.6 Data Analysis
We used flounder “oriented swimming movement” as the behavioral assay to determine
that caused the flounder to swim vertically or come in contact with the perimeter of the
orientation arena for an extended period of time. Fish that did not move for the duration of the
experiment were not included in the analysis. Following the acclimation period, the beginning of
the first 10 directional movements made by each fish were documented, and the angle of this
movement relative to magnetic north was measured using ImageJ software. Any movements
made after the first 10 were determined to be more exploratory rather than directional, and
therefore excluded from the analysis. The angles of movement for a single flounder were
averaged to compute a mean angle of directional preference for a single fish throughout the
duration of the experiment, and graphed using the circular statistics software, Oriana. Using
Oriana, it was possible to conduct a Rayleigh test to search for significant unimodal orientation
within experimental group, and a Mardia-Watson-Wheeler test to assess significance between
experimental groups (Wilkie 1983; Batschelet 1981). Across all tests, a P-value less than 0.05
3. RESULTS
3.1 Magnetic Compass Results
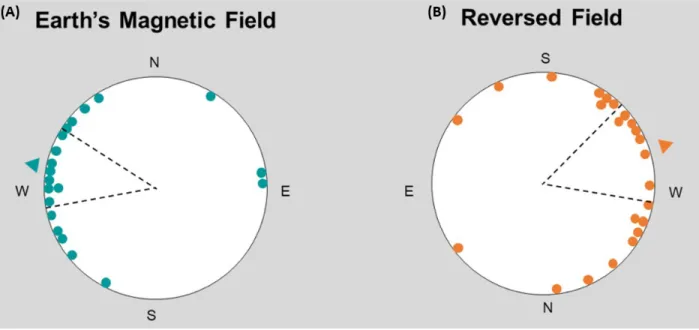
Figure 8. Each dot represents the mean angle of movement for a single flounder and the dashed black lines display 95% confidence intervals. (a) In the Earth’s magnetic field treatment, flounder significantly oriented west (N=20;
Mean Angle= 282°; Rayleigh P= 2.09E-4). (b) In the reversed magnetic north field, flounder significantly oriented
in the approximately the opposite direction, but towards the same magnetic direction (N=23; Mean Angle=72.2°;
Rayleigh P= 5.70E-4).
In the ambient magnetic field present at the experimental location, flounder significantly
oriented towards magnetic west (Figure 8a). Upon reversal of the field, flounder significantly
oriented in approximately the opposite direction, but still towards the same magnetic direction
(Figure 8b). The orientation responses of these two treatments were significantly different from
each other (Mardia-Watson-Wheeler P= 5.63E-5). In both treatments, flounder orientation data
gathered from 2018 and 2019 were not significantly different from one another, and tidal state
3.2 Magnetic Map Results
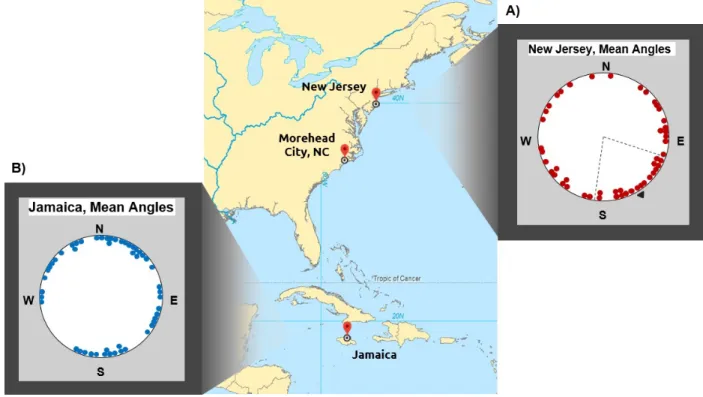
Figure 9. Each dot represents the mean angle of movement for a single fish, and 95% confidence intervals are displayed as dashed black lines. The northern New Jersey magnetic field treatment is illustrated above in red, and
the southern Jamaica magnetic field treatment in blue. Average direction of movement for each treatment is
indicated by a black arrow. (a) In the northern New Jersey treatment, fish significantly oriented towards magnetic
southeast (N=48; Mean Angle= 147°; Rayleigh P= 0.022). (b) In the southern Jamaica treatment, fish exhibited
orientations that could not be distinguished from random (N=51; Rayleigh P=0.148).
In the northern New Jersey treatment with a stronger magnetic field intensity and larger
inclination angle, flounder significantly oriented towards magnetic southeast (Figure 9a). In the
Jamaica treatment with a weaker magnetic field intensity and smaller inclination angle, fish
exhibited orientations that could not be distinguished from random (Figure 9b). Orientations
between these two experimental treatments were significantly different from one another
4. DISCUSSION
Our results suggest that Gulf Flounder possess the ability to use Earth’s magnetic field
for both directional and positional information. In the magnetic compass experiments, fish
oriented towards magnetic west in both the ambient and reversed magnetic field treatments.
Maintaining consistent magnetic orientation even upon reversal of the field illustrates the
presence of a magnetic compass sense, where an organism can use the magnetic field for
directional information. In my magnetic map experiments, flounder exhibited significantly
different orientations between the northern and southern magnetic map treatments. Considering
flounder responded differently to changes in field intensity and inclination angle, these results
suggest that Gulf Flounder possess the ability to determine positional information from Earth’s
magnetic field, or a magnetic map sense.
4.1 Magnetic Compass Experiments
The results of our magnetic compass experiments with Gulf Flounder were consistent
with previous research, where an organism possessing a compass sense was able to consistently
orient towards the same magnetic direction (Lohmann & Lohmann, 1993; Lohmann et al., 1995).
This sense has been discovered in diverse marine organisms, such as reef fish and isopods, and
may provide sufficient directional information to guide organisms between habitats (Bottesch et
al., 2016; Ugolini & Pezzani, 1995). Building off of existing literature, our discovery of a
compass sense in Gulf Flounder illuminates how this sense could also be useful to migratory
flatfish that travel between nearshore and offshore environments. In particular, our results could
explain the consistent directional heading of European plaice, a genetically similar flatfish
There are a couple of possible interpretations of the western directional preference
flounder exhibited in the magnetic compass experiments. First, it is possible that this westerly
preference may be a recruitment response. Considering we collected these flounder during the
summer when they would normally be recruiting, orienting westward may be sufficient to guide
flounder to coastal nursery habitats. However, flounder may also be responding to this cue on a
more local scale. It is possible they may be orienting westward in order to travel along Bogue
Sound, a body of water located near the experimental location of UNC-IMS. Future experiments
should be conducted to determine the scale that flounder are responding to magnetic information.
4.2 Magnetic Map Experiments
In the magnetic map experiments, I found that fish experiencing the northern New Jersey
magnetic field significantly oriented towards magnetic southeast, a direction that would
hypothetically allow them to return to their caught location of Morehead City, NC. Flounder
experiencing the southern magnetic field treatment near Jamaica exhibited orientations that could
not be distinguished from random. I believe these different responses could be explained by the
known distribution of Gulf Flounder, which extends from North Carolina to an undetermined
southern limit near the Gulf of Mexico. Since the New Jersey magnetic field treatment extends
far north above their natural range, flounder would recognize this magnetic field as present in an
uninhabitable environment. The New Jersey coastline is colder than the locations Gulf Flounder
usually occupy, and it would come at a high cost for an individual to dwell in this location.
Therefore, flounder exhibited a strong southern response to leave this magnetic field and return
to a location they could inhabit. This is consistent with previous map experiments, where testing
elicit a strong directional response (Putnam et al., 2014). Additionally, given there is anecdotal
evidence of Gulf Flounder near Jamaica, our southern magnetic field is present in a location that
flounder could feasibly occupy. It is possible that our southern Jamaica field may not have been
outside of the southernmost limit of Gulf Flounder’s range to elicit a northern response, leading
to random orientation.
5. FUTURE STEPS
Previous magnetic navigation research has largely focused on discrete life history stages
of an organism. However, to date, no research has ever been conducted that offered a more
comprehensive view of how the use of this sense transforms throughout ontogeny. Considering
fish at varying have history stages have different objectives, the magnetic sense is likely used
differently throughout an individual’s lifetime. For flounder, it would be fascinating to illustrate
how this sense could be recapitulated in younger life history stages, especially in light of new
empirical evidence of magnetic navigation in Atlantic Haddock larvae and reef fish larvae
(Cresci et al., 2019; O'Connor & Muheim, 2017; Bottesch et al., 2016). Searching for the
presence of a magnetic sense in flounder larvae could illustrate how they may have a more active
role in controlling the trajectory of their recruitment, thereby enhancing in our understanding of
larval dispersal and population connectivity. In the adults, investigating the transformation of the
magnetic sense could elucidate how this sense may be used when navigating offshore towards
spawning locations. Ultimately, understanding the use of the magnetic sense throughout
ontogeny could help us better understand the senses guiding flatfish movements during different
While examining different stages of life history would illustrate the transformation of the
magnetic sense through an individual’s lifetime, another fascinating avenue of research would be
to assess the influence of seasonality on directional preference. Seasonal shifts in magnetic
directional preference have previously been discovered in migratory bird species, such as the
European robin, and migrating moths (Wiltschko & Wiltschko, 2001; Chapman et al., 2008).
This has also demonstrated with aquatic migratory species, where temperature was discovered to
be a large determinant in the strength and directional preference of a species of eel (Durif et al.,
2013). Considering Gulf Flounder either migrate offshore or southwards during the offseason, it
is possible that seasonality and thermal change could transform their summer western response
into a southern response during the fall and winter months. Future research should be conducted
to further investigate the possibility of this seasonal behavior in Gulf Flounder.
Aside from season, the tidal cycle is another possible parameter that has been shown to
influence the directional preference in magnetoreceptive organisms. Many aquatic organisms
utilize tidal change as a method of transport between near and offshore environments
(Tankersley et al., 1998; Gibson, 2003). Recent empirical research that suggests that this method
of transport may be linked to the magnetic compass sense (Cresci et al., 2017). Given tidal
transport has also been discovered in previous tracking studies with European plaice, a
genetically similar flatfish species, it is possible that Gulf Flounder may possess a similar system
(Metcalfe et al., 1990). Understanding how the magnetic compass sense is influenced by tidal
change would provide insight on how flounder may use the tidal cycle to conserve energy and be
6. APPLICATIONS & IMPLICATIONS
Past research on magnetic navigation has largely focused on organisms that undergo
long-distance migrations along complex migratory routes. Discovering a magnetic sense in
flatfish demonstrates how this sense could provide useful navigational information for organisms
that undergo shorter migrations between nearshore and offshore environments. Moreover,
flatfish exhibit a rather common life history strategy comprised of larval dispersal and
subsequent recruitment to coastal habitats. Finding a magnetic sense in flatfish illustrates how
this sense may be present in other organisms that possess similar life history strategies. On a
broader scale, this information serves as a small puzzle piece to the larger developing field of
magnetic sensory research in attempt to deepen the understanding of how sensory cues are
involved in long-distance migrations.
As magnetic sensory research has developed, the processes behind how organisms are
able to detect the Earth’s magnetic field is still a question that still remains unanswered by
scientists. Since the magnetic field passes through the whole organism, the processes behind this
sense could theoretically be present anywhere in biological tissue (Johnsen & Lohmann, 2005).
A couple of hypotheses have been developed in an effort to explain this phenomenon from
observing a number of different organisms. Scientists have found evidence that birds are able to
determine their geographic position by using radical pair-forming molecules in their eyes that
communicate with cryptochrome contained in their beaks (Wiltschko & Wiltschko, 2005).
Elasmobranchs, such as rays and sharks, detect magnetic variations by using an induction-based
method that involves monitoring fluctuations in current generated through their electric organs
(Akoev, 1990). Bats have also been hypothesized to be able to use magnetite and a series of
Flounder’s transparent larval stage provides an interesting opportunity for scientists to search for
these processes within a smaller organism. Their transparency is particularly useful because it
allows for easier scanning for magnetic particles, such as magnetite, that may aid in this sensing
ability (Holland et al., 2008). Overall, the lifecycle and physiological characteristics of Gulf
Flounder allow them to serve as an ideal mechanistic model to resolve the enigma of the physical
processes behind magnetoreception.
Besides being a model organism to search for the magnetoreceptor, many species of
flatfish support important commercial fisheries. In North Carolina, both summer and southern
flounder were found by the Division of Marine Fisheries to be two of the top five fished species
in 2017 and their decline prompted stronger fishing regulations (“NC Flounder Limits”, 2018).
Understanding the senses that guide flatfish movements and migrations could allow for the
construction of successful fishery management decisions, thereby investing in the long-term
prosperity of the flatfish stock and the economies that depend on it.
Gulf flounder possessing both a magnetic compass and map sense could help explain
how flounder navigate the environment they occupy, but also highlights the possible influence
that human magnetic equipment can have on magnetoreceptive organisms. Offshore wind energy
transfers large amounts of current through undersea cables back to the coast, which creates
anomalies in the magnetic field surrounding them (Öhman et al., 2007). This is of particular
concern for flounder, who occupy the ocean floor and could be strongly influenced by these
magnetic anomalies during their nearshore and offshore migrations. Aside from flounder, these
disruptions in the magnetic field have an unknown influence on the orientation, behavior, and
ramifications need to be further explored by scientists and should be taken into consideration
concerning future developments.
REFERENCES
Aas, Ø., Einum, S., Klemetsen, A., Skurdal, J., (2011). Atlantic salmon ecology. Chichester: Wiley-Blackwell.
Akoev, G. N. (1990). Temperature Sensitivity of the Ampullae of Lorenzini of Elasmobranchs. Thermoreception and Temperature Regulation,44-52. doi:10.1007/978-3-642-75076-2_6
Batschelet, E. (1981). Circular statistics in biology. New York: Academic Press.
Boles, L. C., & Lohmann, K. J. (2003). True navigation and magnetic maps in spiny lobsters. Nature, 421(6918), 60–63. doi: 10.1038/nature01226
Bottesch, M., Gerlach, G., Halbach, M., Bally, A., Kingsford, M. J., & Mouritsen, H. (2016). A magnetic compass that might help coral reef fish larvae return to their natal reef. Current Biology, 26(24). doi: 10.1016/j.cub.2016.10.051
Bultel, E., Lasne, E., Acou, A., Guillaudeau, J., Bertier, C., & Feunteun, E. (2014). Migration behaviour of silver eels (Anguilla anguilla) in a large estuary of Western Europe inferred from acoustic telemetry. Estuarine, Coastal and Shelf Science, 137, 23–31. doi: 10.1016/j.ecss.2013.11.023
Cain, S. D., Boles, L. C., Wang, J. H., & Lohmann, K. J. (2005). Magnetic Orientation and Navigation in Marine Turtles, Lobsters, and Molluscs: Concepts and Conundrums. Integrative and Comparative Biology, 45(3), 539-546. doi:10.1093/icb/45.3.539
Chapman, J. W., Reynolds, D. R., Hill, J. K., Sivell, D., Smith, A. D., & Woiwod, I. P. (2008). A seasonal switch in compass orientation in a high-flying migrant moth. Current Biology, 18(19). doi: 10.1016/j.cub.2008.08.014
Cresci, A., Paris, C. B., Durif, C., Shema, S., Bjelland, R. M., Skiftesvik, A. B., & Browman, H. I. (2017). Glass eels (Anguilla anguilla) have a magnetic compass linked to the tidal cycle. Science advances, 3(6), e1602007. doi:10.1126/sciadv.1602007
Gould, J. L. (1998). Sensory bases of navigation. Current Biology, 8(20), R731–R738. https://doi.org/https://doi.org/10.1016/S0960-9822(98)70461-0
Dingle, H. (2014). Migration the biology of life on the move. Oxford: Oxford University Press.
Durif, C. M., Browman, H. I., Phillips, J. B., Skiftesvik, A. B., Vøllestad, L. A., & Stockhausen, H. H. (2013). Magnetic Compass Orientation in the European Eel. PLoS ONE, 8(3). doi:10.1371/journal.pone.0059212
Fischer, J., Freake, M., Borland, S., & Phillips, J. (2001). Evidence for the use of magnetic map information by an amphibian. Animal Behaviour, 62(1), 1–10. doi: 10.1006/anbe.2000.1722
Gardiner, J. M., & Atema, J. (2014). Flow Sensing in Sharks: Lateral Line Contributions to Navigation and Prey Capture. Flow Sensing in Air and Water, 127–146. doi: 10.1007/978-3-642-41446-6_5
Gibson, R. N. (2003). Go with the flow: tidal migration in marine animals. Migrations and Dispersal of Marine Organisms, 153–161. doi: 10.1007/978-94-017-2276-6_17
Gibson, R. N., Nash, R. D., Geffen, A. J., & W., V. D. (2015). Flatfishes: Biology and exploitation. Chichester, West Sussex, UK: Wiley Blackwell.
Gould, J. L., & Gould, C. G. (2012). Natures compass: The mystery of animal navigation. Princeton: Princeton University Press.
Groves, A. B., Collins, G. B., & Trefethen, P. S. (1968). Roles of Olfaction and Vision in Choice
of Spawning Site by Homing Adult Chinook Salmon (Oncorhynchus tshawytscha). Journal
of the Fisheries Research Board of Canada, 25(5), 867–876. doi: 10.1139/f68-082
Holland, R. A., Kirschvink, J. L., Doak, T. G., & Wikelski, M. (2008). Bats Use Magnetite to Detect the Earths Magnetic Field. PLoS ONE,3(2). doi:10.1371/journal.pone.0001676
Johnsen, S., & Lohmann, K. J. (2005). The physics and neurobiology of magnetoreception. Nature Reviews Neuroscience, 6(9), 703-712. doi:10.1038/nrn1745
Kimchi, T., Terkel, J. (2001). Magnetic compass orientation in the bling mole rat Spalax ehrenbergi. Journal of Experimental Biology, 204, 751-758.
Lohmann, K. J., & Lohmann, C. (1993). A Light-Independent Magnetic Compass in the Leatherback Sea Turtle. The Biological Bulletin, 185(1), 149-151. doi:10.2307/1542138
Lohmann, K. J., Lohmann, C. M., Ehrhart, L. M., Bagley, D. A., & Swing, T. (2004). Animal behaviour: Geomagnetic map used in sea-turtle navigation. Nature,428(6986), 909-910. doi:10.1038/428909a
Lohmann, K. J., Putman, N. F., & Lohmann, C. M. (2008). Geomagnetic imprinting: A unifying hypothesis of long-distance natal homing in salmon and sea turtles. Proceedings of the National Academy of Sciences, 105(49), 19096-19101. doi:10.1073/pnas.0801859105
Lohmann, K. J., Pentcheff, N. D., Nevitt, G. A., Stetten, G. D., Zimmer-Faust, R. K., Jarrard, H. E., & Boles, L. C. (1995). Magnetic Orientation of Spiny Lobsters in the Ocean: Experiments with Undersea Coil Systems. The Journal of Experimental Biology. Retrieved December 12, 2018.
Mcmenamin, S. K., & Parichy, D. M. (2013). Metamorphosis in Teleosts. Current Topics in Developmental Biology Animal Metamorphosis, 127–165. doi: 10.1016/b978-0-12-385979-2.00005-8
Merritt, R., Purcell, C., & Stroink, G. (1983). Uniform magnetic field produced by three, four, and five square coils. Review of Scientific Instruments, 54(7), 879-882. doi:10.1063/1.1137480
Metcalfe, J. D., Arnold, G. P., & Webb, P. W. (1990). The energetics of migration by selective tidal stream transport: an analysis for plaice tracked in the southern North Sea. Journal of the Marine Biological Association of the United Kingdom, 70(1), 149–162. doi: 10.1017/s0025315400034275
Metcalfe, J. D., Holford, B. H., & Arnold, G. P. (1993). Orientation of plaice (Pleuronectes platessa) in the open sea: Evidence for the use of external directional clues. Marine Biology, 117(4), 559-566. doi:10.1007/bf00349766
Miyazaki, T., Masuda, R., Furuta, S., & Tsukamoto, K. (1997). Laboratory. Observation on the Nocturnal Activity of Hatchery-reared Juvenile Japanese Flounder Paralichthys olivaceus. Fisheries Science, 63(2), 205–210. doi: 10.2331/fishsci.63.205
Naisbett-Jones, L. C., Putman, N. F., Stephenson, J. F., Ladak, S., & Young, K. A. (2017). A Magnetic Map Leads Juvenile European Eels to the Gulf Stream. Current Biology,27(8), 1236-1240. doi:10.1016/j.cub.2017.03.015
NC Flounder Limits Have Changed. (2018, June 12). Retrieved from https://islandlifenc.com/nc-flounder-limits-changed/
O’Connor, J. O., & Muheim, R. (2017). Pre-settlement coral-reef fish larvae respond to magnetic field changes during the day. 2874–2877. https://doi.org/10.1242/jeb.159491
Öhman, M. C., Sigray, P., & Westerberg, H. (2007). Offshore Windmills and the Effects of Electromagnetic Fields on Fish. AMBIO: A Journal of the Human Environment, 36(8), 630-633. doi:10.1579/0044-7447(2007)36[630:owateo]2.0.co;2
Putman, N., Scanlan, M., Billman, E., O’Neil, J., Couture, R., Quinn, T., . . . Noakes, D. (2014). An Inherited Magnetic Map Guides Ocean Navigation in Juvenile Pacific Salmon. Current Biology,24(4), 446-450. doi:10.1016/j.cub.2014.01.017
Radford, C. A., Stanley, J. A., Simpson, S. D., & Jeffs, A. G. (2011). Juvenile coral reef fish use sound to locate habitats. Coral Reefs, 30(2), 295–305. doi: 10.1007/s00338-010-0710-6
Tankersley, R. A., Wieber, M. G., Sigala, M. A., & Kachurak, K. A. (1998). Migratory Behavior of Ovigerous Blue Crabs Callinectes sapidus: Evidence for Selective Tidal-Stream Transport. The Biological Bulletin, 195(2), 168–173. doi: 10.2307/1542824
Vacha, M., Puzova, T., & Kvicalova, M. (2009). Radio frequency magnetic fields disrupt magnetoreception in American cockroach. Journal of Experimental Biology, 212(21), 3473– 3477. doi: 10.1242/jeb.028670
Wilkie, D. (1983). Rayleigh Test for Randomness of Circular Data. Applied Statistics, 32(3), 311. doi:10.2307/2347954
Wiltschko, W., Gesson, M., & Wiltschko, R. (2001). Magnetic compass orientation of European robins under 565 nm green light. Naturwissenschaften, 88(9), 387-390. doi:10.1007/s001140100248