Adapting to a changing world: RAGgenomics and evolution
Full text
Figure
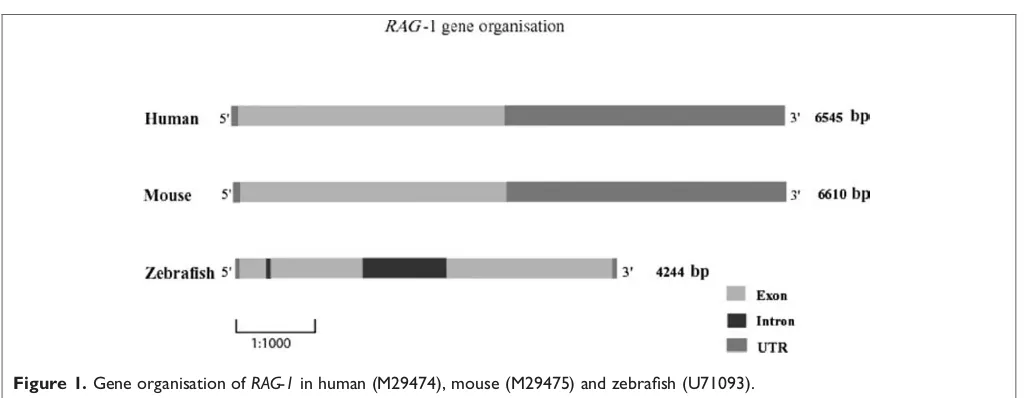
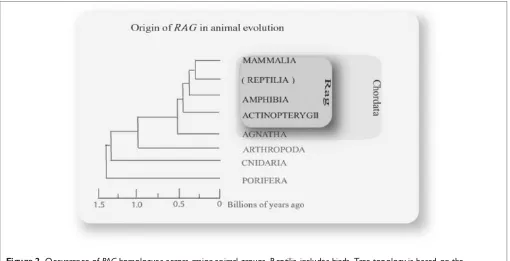
Related documents
It was about the disputes between the Client, Putrajaya Holdings Sdn Bhd (PJH) and the Contractor, Peremba Jaya Sdn Bhd. The disputes has been kept in silence for the last
In their study, Keefer et al found that between 11 and 27 IBS patients experienced sleep quality impairment, 3 to 29 IBS patients had dif fi culty falling asleep, about 11 to 25
The following corollary gives us a fixed point theorem with a contractive condition of integral type for cyclic contractions.... Then T has a unique
expression of monocyte chemoattractant protein 1 in cultured human endothelial
When compared with the results obtained from single fibroblast samples (in single female fibroblast cells, XIST transcripts were detected in 90 per cent and HPRT in 65
Using the results of the 2008 Keeping Cool Survey, we estimate the relative economic disadvantage faced by MS households in trying to keeping cool.. We find that MS households
cfDNA: Cell-free DNA; CMA: Chromosomal microarray analysis; CNV: Copy number variation; CPM: Confined placental mosaicism; CVS: Chorionic villi sampling; NGS:
The proposed cosine similarity clustering algorithm is used to cluster these selected URLs which infers user search goals.. These clustered search results are
