R E S E A R C H
Open Access
Multiplex array proteomics detects increased
MMP-8 in CSF after spinal cord injury
Matthew Light
1, Kenneth H Minor
1, Peter DeWitt
2, Kyle H Jasper
1and Stephen JA Davies
1*Abstract
Introduction:A variety of methods have been used to study inflammatory changes in the acutely injured spinal cord. Recently novel multiplex assays have been used in an attempt to overcome limitations in numbers of
available targets studied in a single experiment. Other technical challenges in developing pre-clinical rodent models to investigate biomarkers in cerebrospinal fluid (CSF) include relatively small volumes of sample and low
concentrations of target proteins. The primary objective of this study was to characterize the inflammatory profile present in CSF at a subacute time point in a clinically relevant rodent model of traumatic spinal cord injury (SCI). Our other aim was to test a microarray proteomics platform specifically for this application.
Methods:A 34 cytokine sandwich ELISA microarray was used to study inflammatory changes in CSF samples taken 12 days post-cervical SCI in adult rats. The difference between the median foreground signal and the median background signal was measured. Bonferroni and Benjamini-Hochburg multiple testing corrections were applied to limit the False Discovery Rate (FDR), and a linear mixed model was used to account for repeated measures in the array.
Results:We report a novel subacute SCI biomarker, elevated levels of matrix metalloproteinase-8 protein in CSF, and discuss application of statistical models designed for multiplex testing.
Conclusions:Major advantages of this assay over conventional methods include high-throughput format, good sensitivity, and reduced sample consumption. This method can be useful for creating comprehensive inflammatory profiles, and biomarkers can be used in the clinic to assess injury severity and to objectively grade response to therapy.
Keywords:biomarker, cerebrospinal fluid (CSF), cytokine, inflammation, matrix metalloproteinase-8 (MMP-8), microarray, proteomics, spinal cord injury, tissue inhibitor of metalloproteinase-1 (TIMP-1).
Introduction
Traumatic injury to the spinal cord can result in life-changing neurological deficits. Patients, family members, clinicians, and researchers alike face significant long-term challenges including (but not limited to) pain man-agement, rehabilitation, and functional improvement. According to 2011 estimates, the incidence of spinal cord injury (SCI) is roughly 12,000 cases per year in the USA, 40% resulting from motor vehicle accidents and 27% result from falls, creating a bimodal age distribution in
the patient population [1]. Long-term implications of these debilitating injuries are significant. Advances in pa-tient care, specifically prevention and treatment of urin-ary tract complications and renal failure, have improved longevity such that causes of mortality are now similar to the general population (cardiovascular disease, cancer, and lower respiratory disease) [2,3].
The molecular mechanisms underlying inflammation-induced damage and recovery of function at the acute, subacute and chronic stages after SCI are currently being pieced together. Although trauma to the spinal cord results in rapid loss of neurons and glia within gray and white matter and transection of white matter path-ways, this acute insult also initiates a robust inflamma-tory response [4-6] that continues to damage tissue * Correspondence:[email protected]
1Department of Neurosurgery, University of Colorado School of Medicine,
Building RC-1 North, Room P18-9400, 12800 E 19th Avenue, Aurora, CO 80045, USA
Full list of author information is available at the end of the article
[7,8], creating new deficits at subacute and chronic time points post-injury. Loss of blood-spinal cord barrier (BSCB) [9] and lymphocytic infiltrate into the injury site [10] potentiates inflammation, and reactive gliosis and formation of scar tissue prevent axon regeneration [5,11,12].
With clinical trials testing the effects of new therapies on neurologic outcomes post-SCI already underway (minocycline and erythropoetin to name a few) [13], and more still in the scientific pipeline, the role of cerebro-spinal fluid (CSF) biomarkers as indicators of injury pro-gression and repair is now more important than ever. Although functional improvement remains the gold standard and certainly the most important outcome for patients, Kwonet al.highlight the potential role for bio-markers in assessing initial injury severity and tracking changes in injury status over time. In addition, they emphasize the role of biomarkers in determining thera-peutic response more objectively than current highly subjective functional outcome measures like the Ameri-can Spinal Injury Association (ASIA) Score [14]. Other important potential applications include characterization of molecular signs of neurotoxic damage and endogen-ous neuro-repair, and discovery of new therapeutic tar-gets. What is needed is a protein-based high-throughput assay that can detect biomarkers from a safe and clinic-ally available source. Sensitivity is also critical because CSF (an important source of biomarkers given its pro-ximity to neuronal tissue) contains relatively low protein concentrations even after BSCB disruption [15]. This ideal assay would be of significant value to researchers and clinicians alike, facilitating delivery of new therapies to SCI patients.
It is becoming increasingly apparent that inflammation plays key roles in both acute and secondary damage, and late-phase recovery after traumatic SCI, and that any novel therapy would almost certainly interact with these processes. In fact, much can be learned from the Immunology field, where researchers have adopted many of the latest technologies to characterize inflammatory processes previously too complex for traditional methods of investigation. Where PCR and standard ELISA were once popular for detecting changes in mRNA and pro-tein respectively, newer multiplex arrays have emerged as the current assays of choice. Recently Leeet al. simul-taneously profiled 10 different cytokine responses to H1N1 infection in hospitalized patients using a cyto-metric bead array system [16], and Mizutaniet al. were able to track serial changes in 17 cytokines using a simi-lar suspension array in patients with Crohn’s disease treated with Infliximab [17]. Oncology is another field that has made good on the promise of newer array tech-nology. Tefferi et al. studied the prognostic value of 30 cytokines in primary myelofibrosis by multiplex
sandwich immunoassay [18]: one of many truly compre-hensive cytokine profiling studies. For a thorough review of multiplex sandwich ELISA technology read Nielsen and Geierstanger [19].
An effort has also been made to validate the reliability of these new assays, focusing mainly on the issues sur-rounding transition from a uniplex to multiplex format. Panget al.studied two commercially available solution-phase microarray platforms against standard ELISA in detecting autoantibodies in human serum [20]. They note similar detection performance, and name high-throughput screening and reduced reagent consumption as major advantages over standard methods. This paper also emphasizes the importance of cutoff values (criteria for determining positive and negative signals in the dataset). In further considering data processing, the multiplex format necessitates different statistical treat-ment of the data: the need for multiple hypothesis test-ing. The concept of the false discovery rate (FDR) comes from earlier work in epidemiology and genome-wide association studies: the idea that as the number of statistical tests performed increases, so does the number of false positives. Hsueh et al. compare strategies like Benjamini-Hochburg [21] for estimating the FDR, and emphasize the importance of controlling the type I error rate in multiplicity testing [22]. Finally, important technical issues such as binding‘inhomogeneity’across immobilized spot-based arrays [23] are currently under investigation. To summarize, multiplex proteomic assays, although rela-tively new, offer many qualities of the ‘ideal assay’ described above, provided there is: correct statistical treat-ment of the data, careful consideration of technical issues, and verification of ‘hits’ with appropriate confirmatory methods.
The challenge of characterizing the inflammatory response to acute SCI has been met using a variety of methods from flow cytometry, to PCR, to ELISA. Streit
24 hours post-injury by mass spectrometry of which 10 correlated with injury severity (ASIA Score) including Ywhaz, Itih4, and Gpx3 [27].
In an effort to characterize inflammatory markers present at subacute time points after traumatic SCI, thought to represent a clinically relevant ‘window of opportunity’ for therapeutic intervention, we have conducted a proteomics analysis of CSF from rats at 12 days after cervical spinal cord contusion for changes in 34 cytokines using a multiplex sandwich ELISA micro-array. Initial statistical analysis yielded seven proteins (IL-1α, Leptin, B7-2/CD86, GM-CSF, IL-1β, matrix metalloproteinase-8 (MMP-8), and tissue inhibitor of metalloproteinase-1 (TIMP-1)), the levels of which were all increased in the CSF from injured animals compared with normal uninjured controls. The FDR was controlled (limit type I errors) by adjusting theP-values using both Benjamini-Hochburg and Bonferroni methods (although we chose to focus our analysis on Benjamini-Hochburg adjustments because this method generally allows more potential hits to reach significance). To account for tech-nical replicates, as well as variation in regional signal intensity within the array, a linear mixed model was ap-plied, which showed only MMP-8 was significantly ele-vated in injured CSF, a finding subsequently confirmed by western blot.
Methods
Spinal cord injury model
Animal care and experimental protocols were carried out according to the guidelines of the National Insti-tutes of Health as well as the policies of our Institu-tional Animal Care and Use Committee at the University of Colorado Anschutz Medical Campus. Female Sprague–Dawley rats (Harlan Laboratories, Denver, CO, USA) aged between 3 months and 5
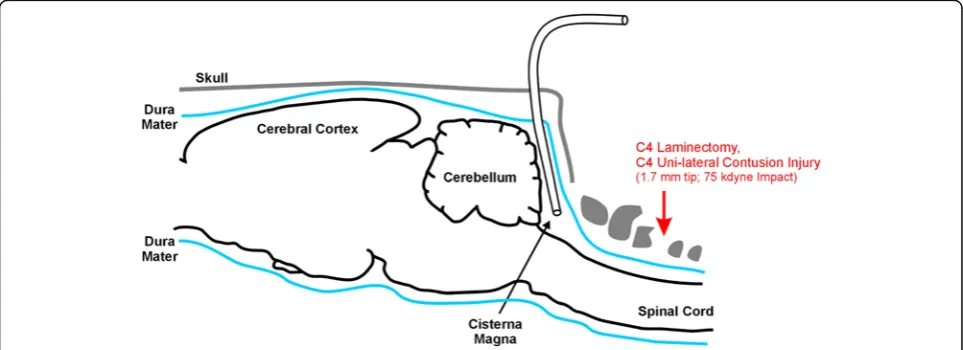
months were used. Animals were anesthetized with an intraperitoneal injection of ketamine and xylazine, shaved and placed in a stereotactic device for small ro-dent surgery (David Kopf Instruments, Tujunga, CA, USA). Lubricating eye gel was applied to prevent drying under anesthesia. After cleaning the surgical site, a mid-line incision was made extending from the shoulder blades to the base of the skull. Another midline incision at the origin of the trapezius muscles was made, and the paraspinal muscles were scraped from the spinous processes and lamina of C2 to C6 with a small scalpel blade, exposing the spinal column. Hemostasis was achieved using gauze with light pressure. Once the spinal column was exposed, the posterior longitudinal ligament was cut above and below C4. Rongeurs were used to perform a C4 laminectomy, keeping the dura intact.
An Infinite Horizon impactor (Precision Systems and Instrumentation, Fairfax, VA, USA) with a 1.7 mm tip was used to create a unilateral dorsolateral funiculus contusion of the cervical cord. After laminectomy the animal was secured into the impactor by clamping the vertebral bodies of C3 and C5 using Adson stabilizing forceps. Contusion was made by dropping a weight with a preset force of 75 kdynes on the lateral aspect of the cord at the C4 level with a dwell time set at zero. Both displacement and force were recorded to minimize injury variation. The animal was then returned to the stereotactic device. The trapezius muscles were approxi-mated using simple interrupted stitches (4–0 size vicryl suture). Skin was closed with surgical staples.
Cerebrospinal fluid collection
Animals were again anesthetized at 12 days following contusion and were placed in the stereotactic device. The old incision was extended rostrally between the ears
to expose the occipital bone of the skull. Subcutaneous tissue was scraped off the scull with a scalpel, and a small burr hole (from 1 mm to 2 mm diameter) was drilled through the occipital bone, exposing the menin-ges. A slit was then made in the dura and a small diam-eter cannula (order number 7741, Durect Corp., Cupertino, CA, USA) was carefully guided under the dura (in a caudal direction) along the internal surface of the occipital bone and into the cisterna magna (depth of 7 mm). A syringe was used to draw approximately 200 μL of CSF from each animal, making sure the sample was not bloody. All grossly bloody taps were excluded from the analysis (three in total). Protease inhibitor cock-tail (Sigma-Aldrich, St. Louis, MO, USA) was quickly added and samples were placed on dry ice. Samples were stored at −80°C until further use. Animals were then sacrificed by intrahepatic injection of ketamine and xyla-zine. Control CSF was obtained from age-matched ani-mals (no contusion) in a similar fashion. Figure 1 shows a schematic of injury location and cannula placement for CSF extraction.
Cerebrospinal fluid analysis
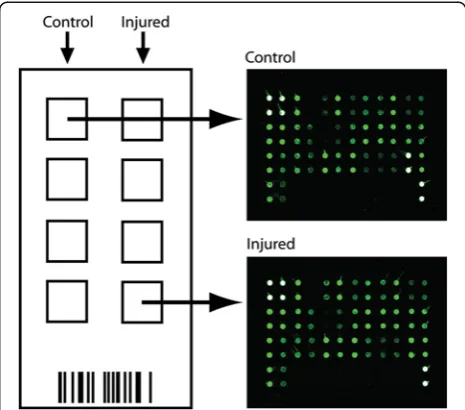
Samples were removed from−80°C, thawed on ice, and briefly centrifuged. CSF from injured animals was com-pared with control CSF (n= 4) using the 34 cytokine preconfigured sandwich ELISA Rat Cytokine Array G2 (RayBiotech, Norcross, GA, USA). To reduce batch variability100μL of each CSF sample (eight total) occu-pied a single array such that only one chip was used for the experiment. CSF samples were not diluted. The chip was read using a GenePix 4000B Microarray Scanner (Molecular Devices, Sunnyvale, CA, USA). Of note, the array was arranged such that each antibody was spotted twice, creating two technical replicates per protein of interest. Any remaining CSF was aliquoted and placed back at−80°C. Specific protocol details can be found at the website of RayBiotech Inc [28]. (http://www.raybio-tech.com). A schematic of the chip format and image of representative control and injured arrays in the Cy3 channel are shown in Figure 2. Layout of spotted pri-mary antibodies is shown in Table 1 (adapted from Ray-Biotech Inc.).
Statistical analysis
Analysis of the data was by the following steps. The dataset was read into R [29] and each of the 96 probe sets (includes cytokines, positive, and negative controls) were tested for significance. A two sample, equal vari-ance t-test was used to test the hypothesis H0:μ1 =μ2 versus H1: μ16¼μ2 where μ1 is the mean response for the injured rats and μ2 is the mean response for the non-injured rats. The response in this case is the differ-ence between the median foreground signal and the
median background signal on the Cy3 channel. RawP-values for each of the 96 probe sets were recorded. Bonferroni and Benjamini-Hochburg multiple testing corrections were used to adjust P-values. Accounting for the technical replicates was done using linear mixed models and done in R via the nlme package [30].
Western blot analysis
MMP-8 western blot
The same CSF samples that had been used for micro-array were removed from −80°C and thawed on ice. Samples (30μL each) from two injured and two control animals were run on a 12.5% polyacrylamide gel (Bio-Rad Life Science, Hercules, CA, USA), and transferred overnight onto PVDF. Westerns were run in duplicate and probed with a polyclonal antibody to MMP-8 (Abcam, Cambridge, MA, USA). Blots were also probed for Transferrin (Santa Cruz Biotechnology, Santa Cruz, CA, USA), previously shown to be unaffected by SCI [27], as a loading control. Rat lung lysate was loaded as a positive control for MMP-8.
TIMP-1 western blot
CSF from injured and control animals (samples not included in multiplex analysis) were in the same way as MMP-8 western blots (above). Lysate from 293 T cells over-expressing TIMP-1 (Santa Cruz) and rat whole brain lysate (Abcam) were used for positive controls. The blot was cut and probed separately for transferrin and TIMP-1 (Santa Cruz Biotechnology).
Results
Microarray data
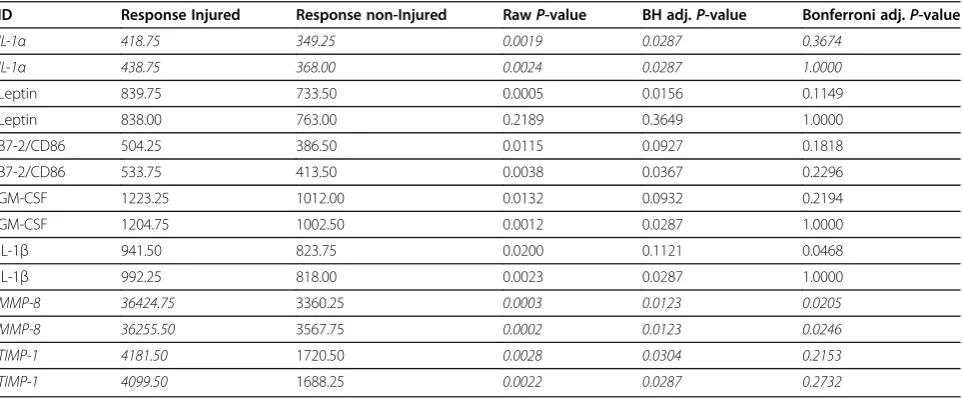
Out of 96 probe sets tested (48 spots with replicates), seven proteins showed a statistically significant difference in median signal intensity (background subtracted) in at least one technical replicate (the response). Proteins with significant results at the 5% level are reported in Table 2. Responses from both technical replicates are shown for comparison along with the corresponding raw and adjusted P-values. IL-1α, Leptin, B7-2/CD86, GM-CSF, IL-1β, MMP-8, and TIMP-1 were all increased in CSF of injured animals compared with controls. Of these results, only IL-1α, MMP-8, and TIMP-1 showed significant increases across both replicate sets after Benjamini-Hochburg adjustment. As expected, there were no statistically significant differences among control datasets (data not shown). Bonferroni adjusted P-values are shown for comparison.
Microarray data
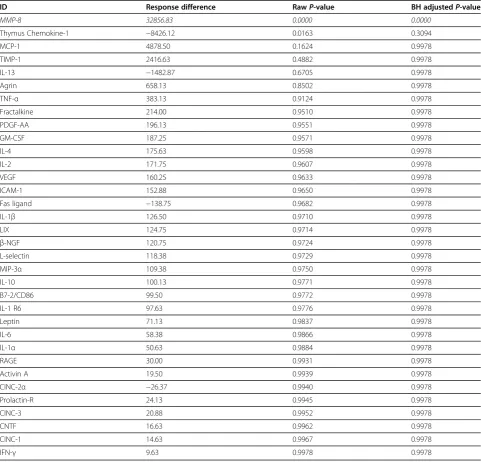
To account for technical replicates, as well as regional signal variability across the array as a whole, a linear mixed model was applied to the data. Table 3 displays the full table of results from the mixed model. The value displayed is the difference between the average of median signal intensity minus the median background intensity (across all technical replicates) for injured and control animals. Positive values indicate an increase in protein in injured animals, and negative values indicate a decrease. Results found to have statistically different sig-nals between the injured and non-injured animals based on raw P-values are highlighted. In this model, only MMP-8 and Thymus Chemokine-1 showed statistically significant changes based on raw P-values. MMP-8 was elevated in injured animals: the same conclusion as from the probe set level tests (simple model). Thymus Chemokine-1 did not reach significance in the simple Table 1 Primary antibody key
Pos Pos Pos Neg Act A Agrin B7-2/CD86 β-NGF CINC-1 CINC-2α CINC-3 CNTF
Pos Pos Pos Neg Act A Agrin B7-2/CD86 β-NGF CINC-1 CINC-2α CINC-3 CNTF
Fas-L Fract GM-CSF ICAM-1 IFN-γ IL-1α IL-1β IL-1 R6 IL-2 IL-4 IL-6 IL-10
Fas-L Fract GM-CSF ICAM-1 IFN-γ IL-1α IL-1β IL-1 R6 IL-2 IL-4 IL-6 IL-10
IL-13 Leptin LIX L-Sel MCP-1 MIP-3α MMP-8 PDGF-AA PRL- R RAGE TK-1 TIMP-1
IL-13 Leptin LIX L-Sel MCP-1 MIP-3α MMP-8 PDGF-AA PRL-R RAGE TK-1 TIMP-1
TNF-α VEGF Neg Neg Neg Neg Neg Neg Neg Neg Neg Neg
TNF-α VEGF Neg Neg Neg Neg Neg Neg Neg Neg Neg Pos
Antibody layout for each array on microarray chip AAR-CYT-G2 (image adapted from RayBiotech). Each primary antibody is spotted twice per array; eight arrays are spotted per chip. Positive and negative controls determine signal variation between arrays.
Table 2 Simple model: two sample equal variancet-tests
ID Response Injured Response non-Injured RawP-value BH adj.P-value Bonferroni adj.P-value
IL-1α 418.75 349.25 0.0019 0.0287 0.3674
IL-1α 438.75 368.00 0.0024 0.0287 1.0000
Leptin 839.75 733.50 0.0005 0.0156 0.1149
Leptin 838.00 763.00 0.2189 0.3649 1.0000
B7-2/CD86 504.25 386.50 0.0115 0.0927 0.1818
B7-2/CD86 533.75 413.50 0.0038 0.0367 0.2296
GM-CSF 1223.25 1012.00 0.0132 0.0932 0.2194
GM-CSF 1204.75 1002.50 0.0012 0.0287 1.0000
IL-1β 941.50 823.75 0.0200 0.1121 0.0468
IL-1β 992.25 818.00 0.0023 0.0287 1.0000
MMP-8 36424.75 3360.25 0.0003 0.0123 0.0205
MMP-8 36255.50 3567.75 0.0002 0.0123 0.0246
TIMP-1 4181.50 1720.50 0.0028 0.0304 0.2153
TIMP-1 4099.50 1688.25 0.0022 0.0287 0.2732
model, but showed a statistically significant decrease in the linear mixed model based on the raw P-value; a re-sult that did not hold up after Benjamini-Hochburg ad-justment. This discrepancy is the result of significant variability within the Thymus Chemokine-1 dataset com-bined with large differences in the magnitude of signal between injured and control animals. After Benjamini-Hochburg correction of mixed model data, MMP-8 was the only cytokine with a significant change between injured and non-injured animals (P<0.0001). Figure 3
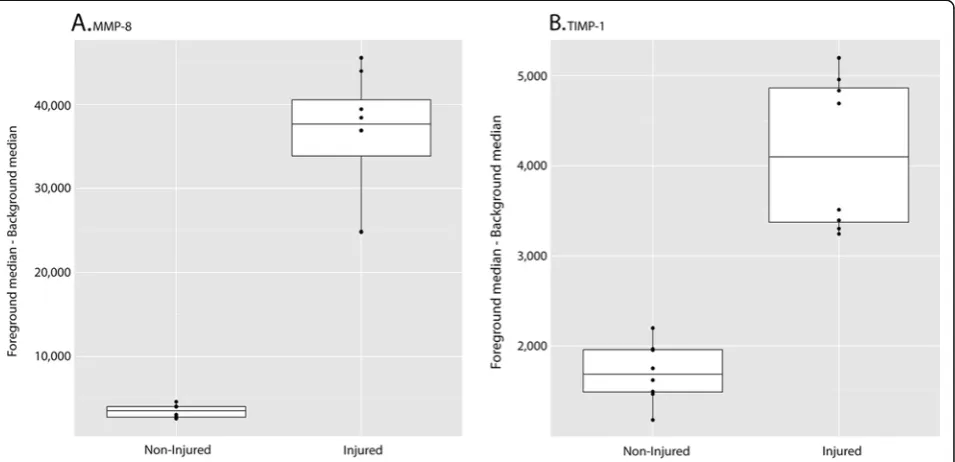
shows all median response data (n= 8) from the MMP-8 and TIMP-1 datasets in box plot format.
Western blot analysis
To confirm that microarray samples contained a protein with the correct molecular weight for MMP-8, CSF from injured and control animals were run and probed for MMP-8. A probe for Transferrin was used as a loading control. Westerns for both MMP-8 and Transferrin were aligned to create Figure 4(a). One representative set of Table 3 Linear mixed model results
ID Response difference RawP-value BH adjustedP-value
MMP-8 32856.83 0.0000 0.0000
Thymus Chemokine-1 −8426.12 0.0163 0.3094
MCP-1 4878.50 0.1624 0.9978
TIMP-1 2416.63 0.4882 0.9978
IL-13 −1482.87 0.6705 0.9978
Agrin 658.13 0.8502 0.9978
TNF-α 383.13 0.9124 0.9978
Fractalkine 214.00 0.9510 0.9978
PDGF-AA 196.13 0.9551 0.9978
GM-CSF 187.25 0.9571 0.9978
IL-4 175.63 0.9598 0.9978
IL-2 171.75 0.9607 0.9978
VEGF 160.25 0.9633 0.9978
ICAM-1 152.88 0.9650 0.9978
Fas ligand −138.75 0.9682 0.9978
IL-1β 126.50 0.9710 0.9978
LIX 124.75 0.9714 0.9978
β-NGF 120.75 0.9724 0.9978
L-selectin 118.38 0.9729 0.9978
MIP-3α 109.38 0.9750 0.9978
IL-10 100.13 0.9771 0.9978
B7-2/CD86 99.50 0.9772 0.9978
IL-1 R6 97.63 0.9776 0.9978
Leptin 71.13 0.9837 0.9978
IL-6 58.38 0.9866 0.9978
IL-1α 50.63 0.9884 0.9978
RAGE 30.00 0.9931 0.9978
Activin A 19.50 0.9939 0.9978
CINC-2α −26.37 0.9940 0.9978
Prolactin-R 24.13 0.9945 0.9978
CINC-3 20.88 0.9952 0.9978
CNTF 16.63 0.9962 0.9978
CINC-1 14.63 0.9967 0.9978
IFN-γ 9.63 0.9978 0.9978
BH, Benjamini-Hochburg; MMP, matrix metalloproteinase; TIMP-1, tissue inhibitor of metalloproteinase-1.
data is shown. Western blot for MMP-8 shows a band at approximately 53 kD (predicted size for rat MMP-8) in the lung positive control, as well as bands of the same size in each CSF sample. A clear increase in MMP-8 pro-tein was observed in samples from injured animals com-pared with controls, but more importantly there was no
evidence of immune-reactive proteins or breakdown ducts. The Transferrin blot shows comparable total pro-tein concentrations in each sample as indicated by relative band size/intensity.
Western blot for TIMP-1 was also performed to test for type II errors in the statistical analysis. Specifically we were interested in testing whether there were hits present in the simple model that did not reach signifi-cance in the mixed model. The western blot for TIMP-1 along with Transferrin loading controls are shown in Fig-ure 4(b). Bands at approximately 25 kD are seen in posi-tive controls (cells overexpressing TIMP-1 and brain extracts) but not in CSF samples. This result is not unex-pected given the relatively weak signal intensity for TIMP-1 in the multiplex array compared with MMP-8, which indicated lower protein concentrations (or weaker antigen-antibody affinity) that were likely below the limit of detection by our western blot system.
Discussion
The primary objective of this study was to characterize the inflammatory profile present in CSF at a subacute time point in a clinically relevant rodent model of trau-matic SCI. Our other aim was to test a microarray pro-teomics platform specifically for this application. Studying inflammation at 12 days post-SCI is highly rele-vant in that it falls within a clinical‘window of opportun-ity’where patients have been stabilized and are generally more receptive to therapy. We also agree with the SCI field in general that CSF is an important source of Figure 3Array data for MMP-8 (A) and TIMP-1 (B) represented in box plot format.The plots illustrate the distribution of the median background subtracted signal for the eight measurements (four rats × two reps) in each of the Injured/Control groups. The height of the box plot gives a measure of spread MMP, matrix metalloproteinase; TIMP-1, tissue inhibitor of metalloproteinase-1.
Figure 4CSF western blot analysis. (a)MMP-8 levels are elevated in injured animals (I) compared with controls (C) 12 days post-contusion (n= 4 for each condition). Rat lung lysate (L) was loaded as a positive control. Blots were probed for Transferrin as a loading control.(b)TIMP-1 is observed in over-expressing 293 cells (293) and rat brain lysate (B), but not in CSF CSF, cerebrospinal fluid; MMP, matrix metalloproteinase; TIMP-1,tissue inhibitor of
potential biomarkers given that it is safe to obtain, read-ily available, and in close proximity to neuronal tissue of interest. Using small volumes of CSF we were able to study 34 cytokines/growth factors simultaneously, detecting significant changes in seven inflammatory mar-kers. After applying statistical models designed for multi-plex testing, we also found robust increases in MMP-8 within CSF, which to our knowledge represents a novel subacute phase biomarker for SCI.
As with any new technology, comparison of results to previous data is crucial in assessing the reliability of the new system. It is generally accepted that a multiphasic inflammatory response occurs after SCI: an early phase peaking at 24 hours and extending from 9 to 10 days, and a later phase between 14 and 180 days post-injury [31]. Studies of inflammation in the setting of SCI have primarily focused their analysis on this early phase be-tween 0 and 72 hours post-injury when neuroprotective strategies are most effective, making comparisons to this study somewhat difficult. Changes in mRNA from rat cord lysate observed by Streitet al. (IL-1β, TNF-α, IL-6, and M-CSF) were transient, returning to normal levels by 24 hours post-injury, and remaining at these levels for the remainder of their analysis (10 days) [24]. In this study, TGF-βlevels started to rise at 24 hours, peaked at 7days, and remained elevated at 10 days; TGF-βwas not included in our analysis. The study by Wang et al. showed elevated IL-1β protein in CSF at one hour [25]. Their result differed from Streit et al. in that protein levels remained elevated at 7 days, but fits nicely with our IL-1βsimple model result. Although changes in IL-1β did not hold up to mixed model analysis, it is quite possible that this result is real, and that we are observing a lull in protein concentration between early- and late-phase inflammation. Further analysis is needed to con-firm this result. Yanget al. showed elevated levels of IL-1β, IL-6, and TNF-αby western blot from cord lysates, a result that peaked at 6 hours, returned to normal by 24 hours, and was dependent on injury severity [6]. IL-6 and TNF-α levels in our study, although elevated in injured animals, did not reach statistical significance. Finally, the human CSF study by Kwon and colleagues did not show any changes in IL-1βor GM-CSF protein over a 5-day period [26] (both elevated in our simple model at 12 days), a difference that may be explained by the observation window, and/or cross-species differences in the inflammatory response.
MMPs and their endogenous inhibitors (TIMPs) have been studied extensively in the setting of SCI. de Castro
et al.showed increases in gelatinases 9 and MMP-2 at 1MMP-2 to14 hours, and 5 days respectively in injured rat spinal cord by zymography. They suggest (using neutral-izing antibody studies) that these MMPs are released by infiltrating neutrophils responding to inflammation [32].
Wells et al. performed a comprehensive MMP/TIMP mRNA profile in an acute murine model of SCI. They showed an increase in MMP-3, 7, 10, 11, 19, and 20 at 24 hours post-SCI, and elevated levels of MMP-2, 12, and 13, but not MMP-8 at 5 days, colocalizing MMP-12 mRNA to Iba1 positive macrophages in the lesion epi-center [33].Using microarray proteomics, we present a new finding, an increase in MMP-8 protein, also known as neutrophil collagenase or collagenase-2, in CSF at 12 days after SCI in rats. This result is consistent with find-ings of Veeravalli et al. who showed an increase in MMP-8 mRNA following SCI between 3 and 21 days post-injury in mouse cord lysate samples [34]. Increases in TGF- β mRNA 10 days post-SCI reported by Streit
et al. [24] may in part explain the elevated levels of MMP-8 in our study, as TGF- β is a known inducer of other collagenases like MMP-13, through the Smad3 pathway [35], as well as MMP-2, and MMP-9 though ERK-1/2, and p38 MAPK [36]. In the Streit study [24], TGF-β levels started to rise at 24 hours, peaked at 7 days, and remained elevated at 10 days; TGF-βhowever was not included in our analysis. Our finding also corre-lates with data from the stroke field, where Cuadrado and colleagues reported upregulation of MMP-8 protein (and other MMPs) in ischemic brain tissue [37]. Finally, the fact that increases in CSF MMP-8 protein levels were not reported by Lubienieckaet al.[27], despite the use of sensitive mass spectrometry analysis at time points ran-ging from 12 to 72 hours after injury, indicates that our observed increase in MMP-8 protein in CSF represents a novel subacute-phase SCI biomarker. Increases in a var-iety of MMPs within the spinal cord following SCI are thought to significantly contribute to further tissue dam-age particularly at acute time points after SCI. At present, however, relatively little is known about CSF levels of specific MMP mRNAs and proteins or their im-pact on tissue damage at subacute time points after SCI. Elevated levels of MMP-12 mRNA in spinal cord injured tissue observed by Wells and colleagues [33] have been shown to correlate with disruption of the BSCB, however this study did not extend beyond 5 days post-SCI. Whether the significant increase in MMP-8 protein we have observed in the CSF contributes to BSCB disruption and other mechanisms of tissue damage post-SCI requires further investigation beyond the scope of the present study.
verifying protein size when using an ELISA-based system because other immunoreactive proteins may disrupt the analysis.
TIMP-1, by contrast, only reached statistical signifi-cance in our simple model. Lack of signifisignifi-cance in the linear mixed model indicates that this result is more likely to be a type 1 error than a true positive. Western blot was performed to test this hypothesis. Although it is possible that our inability to detect TIMP by western blot reflects the ability of the mixed model to control the FDR, it is also possible that TIMP-1 is truly elevated fol-lowing SCI and, as discussed above, the microarray was just more sensitive at identifying biologically significant changes in this molecule.
Clearly, uncertainty exists when interpreting the results of multiplex testing. Different statistical treatment of the data gives us clues, but follow-up testing is often required by conventional methods. This is especially true when using the Benjamini-Hochburg correction, accept-ing a certain number of false positives per assay. How-ever, there are ways, within the confines of the multiplex format, to reduce uncertainty. One obvious option would be to increase the sample size of each dataset. If we were to run two chips instead of one would we not increase our power to detect differences in the data? This brings up the concept of batch variability: the observation of significant variation between batches (chips in our ex-periment). This variability commonly offsets any statis-tical ‘gain’ from the corresponding increase in sample size. A better alternative would be to run sequential arrays, one for screening, as presented here, and another ‘confirmation array’, where only hits from the screening array are spotted, in greater numbers. This sequential ap-proach, although more expensive and time consuming, avoids the problem of batch variation, and prevents con-trols for the FDR from making costly type II errors.
Conclusions
To our knowledge our study is the first to demonstrate significant increases in MMP-8 protein in CSF after trau-matic injury to the spinal cord. We propose that MMP-8 is a novel subacute-phase CSF biomarker for SCI. In addition, our study supports the use of sandwich micro-array proteomics as a valid approach for studying CSF in the setting of SCI in rats, and potentially human SCI patients. This assay reliably detects low levels of CSF protein, and has several advantages over conventional methods including quick and easy protocol, high-throughput format, and use of relatively small amounts of precious sample. Disadvantages include relative cost compared with traditional methods, and the need to con-firm hits (either with repeat microarray analysis or other assays). Moreover our study stresses the importance of controlling the FDR given the multiplex format. In
summary, this assay has the potential to detect new tar-get molecules, characterize complex time-dependent pathophysiologic responses to SCI (like inflammation) and, as more therapies reach the clinic, identify biomar-kers with which to assess injury severity and grade re-sponse to treatment.
Abbreviations
ASIA: American Spinal Injury Association; BH: Benjamini-Hochburg multiple testing correction; BSCB: blood-spinal cord barrier; CNS: central nervous system; CSF: cerebrospinal fluid; ELISA: enzyme-linked immunosorbent assay; FDR: false discovery rate; IL: interleukin; MMP: matrix metalloproteinase; mRNA: messenger RNA; PCR: polymerase chain reaction; SCI: spinal cord injury; TIMP: tissue inhibitor of metalloproteinase; TNF: tumor necrosis factor.
Competing interests
The authors declare that they have no competing interests.
Authors’contributions
ML, KM, and SD designed the study. KM conducted all animal injuries and KM and ML performed CSF extractions. ML and KJ performed the microarray and western blot analysis. PD performed all statistical analysis. Manuscript was written by ML, PD, KM and SD. All authors have read and approved the final manuscript.
Acknowledgements
Funding for the study was provided in part by the Lone Star Paralysis Foundation, Lloyd and Floyd Holman, the David Van Wagener SCI Fund, and the CareCure SCI community.
We acknowledge Martha Miles and Sean Carriedo of Molecular Devices Corp. who provided technical assistance.
Author details
1Department of Neurosurgery, University of Colorado School of Medicine,
Building RC-1 North, Room P18-9400, 12800 E 19th Avenue, Aurora, CO 80045, USA.2Colorado Biostatistics Consortium, Department of Biostatistics and Informatics, University of Colorado Denver, 12477 E 19th Avenue, Room 102, Aurora, CO 80045, USA.
Received: 23 November 2011 Accepted: 4 May 2012 Published: 11 June 2012
References
1. National Spinal Cord Injury Statistical Center:Spinal cord injury: facts and figures at a glance.J Spinal Cord Med2005,28:379–380.
2. Miniño AM:Death in the United States, 2009. NCHS data brief, no. 64. Hyattsville, MD: National Center for Health Statistics; 2011. http://www.cdc. gov/nchs/data/databriefs/db64.pdf.
3. van den Berg ME, Castellote JM, de Pedro-Cuesta J, Mahillo-Fernandez I: Survival after spinal cord injury: a systematic review.J Neurotrauma2010, 27:1517–1528.
4. Fleming JC, Norenberg MD, Ramsay DA, Dekaban GA, Marcillo AE, Saenz AD, Pasquale-Styles M, Dietrich WD, Weaver LC:The cellular inflammatory response in human spinal cords after injury.Brain: J Neurol2006, 129:3249–3269.
5. Leal-Filho MB:Spinal cord injury: from inflammation to glial scar.Surg Neurol Int2011,2:112.
6. Yang L, Jones NR, Blumbergs PC, Van Den Heuvel C, Moore EJ, Manavis J, Sarvestani GT, Ghabriel MN:Severity-dependent expression of pro-inflammatory cytokines in traumatic spinal cord injury in the rat.J Clin Neurosci2005,12:276–284.
7. Blight AR:Macrophages and inflammatory damage in spinal cord injury.J Neurotrauma1992,9(Suppl. 1):S83–S91.
8. Pruss H, Kopp MA, Brommer B, Gatzemeier N, Laginha I, Dirnagl U, Schwab JM:Non-Resolving Aspects of Acute Inflammation after Spinal Cord Injury (SCI): Indices and Resolution Plateau.Brain Pathol2011,21:652–660. 9. Cohen DM, Patel CB, Ahobila-Vajjula P, Sundberg LM, Chacko T, Liu SJ,
10. Ankeny DP, Popovich PG:Mechanisms and implications of adaptive immune responses after traumatic spinal cord injury.Neuroscience2009, 158:1112–1121.
11. Fehlings MG, Hawryluk GW:Scarring after spinal cord injury.J Neurosurg Spine2010,13:165–167. discussion 167–168.
12. Silver J, Miller JH:Regeneration beyond the glial scar.Nat Rev Neurosci
2004,5:146–156.
13. Gensel JC, Donnelly DJ, Popovich PG:Spinal cord injury therapies in humans: an overview of current clinical trials and their potential effects on intrinsic CNS macrophages.Exp Opin Therapeutic Targets2011, 15:505–518.
14. Kwon BK, Casha S, Hurlbert RJ, Yong VW:Inflammatory and structural biomarkers in acute traumatic spinal cord injury.Clin Chem Lab Med: CCLM/FESCC2011,49:425–433.
15. Reiber H:Cerebrospinal fluid-physiology, analysis and interpretation of protein patterns for diagnosis of neurological diseases.Mult Scler1998, 4:99–107.
16. Lee N, Wong CK, Chan PK, Chan MC, Wong RY, Lun SW, Ngai KL, Lui GC, Wong BC, Lee SK, Choi KW, Hui DSC:Cytokine response patterns in severe pandemic 2009 H1N1 and seasonal influenza among hospitalized adults.
PLoS One2011,6:e26050.
17. Mizutani T, Akasaka R, Tomita K, Chiba T:Serial changes of cytokines in Crohn's disease treated with infliximab.Hepatogastroenterol2011,58(110– 111):1523–1526.
18. Tefferi A, Vaidya R, Caramazza D, Finke C, Lasho T, Pardanani A:Circulating interleukin (IL)-8, IL-2R, IL-12, and IL-15 levels are independently prognostic in primary myelofibrosis: a comprehensive cytokine profiling study.J Clin Oncol2011,29:1356–1363.
19. Nielsen UB, Geierstanger BH:Multiplexed sandwich assays in microarray format.J Immunol Methods2004,290:107–120.
20. Pang S, Smith J, Onley D, Reeve J, Walker M, Foy C:A comparability study of the emerging protein array platforms with established ELISA procedures.J Immunol Methods2005,302:1–12.
21. Benjamini Y, Hochburg Y:Controlling the false discovery rate: a practical and powerful approach to multiple testing.J Royal Stat Soc Series B (Methodol)1995,57:289–300.
22. Hsueh HM, Chen JJ, Kodell RL:Comparison of methods for estimating the number of true null hypotheses in multiplicity testing.J Biopharm Stat
2003,13:675–689.
23. Zhao M, Wang X, Nolte D:Mass-transport limitations in spot-based microarrays.Biomed Opt Express2010,1:983–997.
24. Streit WJ, Semple-Rowland SL, Hurley SD, Miller RC, Popovich PG, Stokes BT: Cytokine mRNA profiles in contused spinal cord and axotomized facial nucleus suggest a beneficial role for inflammation and gliosis.Exp Neurol
1998,152:74–87.
25. Wang CX, Olschowka JA, Wrathall JR:Increase of interleukin-1beta mRNA and protein in the spinal cord following experimental traumatic injury in the rat.Brain Res1997,759:190–196.
26. Kwon BK, Stammers AM, Belanger LM, Bernardo A, Chan D, Bishop CM, Slobogean GP, Zhang H, Umedaly H, Giffin M, Street J, Boyd MC, Paquette SJ, Fisher CG, Dvorak MF:Cerebrospinal fluid inflammatory cytokines and biomarkers of injury severity in acute human spinal cord injury.J Neurotrauma2010,27:669–682.
27. Lubieniecka JM, Streijger F, Lee JH, Stoynov N, Liu J, Mottus R, Pfeifer T, Kwon BK, Coorssen JR, Foster LJ, Grigliatti TA, Tetzlaff W:Biomarkers for severity of spinal cord injury in the cerebrospinal fluid of rats.PLoS One
2011,6:e19247.
28. RayBiotech, Inc:, . http://www.raybiotech.com (November 12, 2011). 29. Team RDC:R: A Language and Environment for Statistical Computing. Vienna,
Austria: R Foundation for Statistical Computing; 2011.
30. Pinheiro J, Bates D, DebRoy S, Sarkar D, nlme:Linear and Nonlinear Mixed Effects Models. R package version 3.1-102 edition. Vienna, Austria: R Foundation for Statistical Computing; 2011.
31. Beck KD, Nguyen HX, Galvan MD, Salazar DL, Woodruff TM, Anderson AJ: Quantitative analysis of cellular inflammation after traumatic spinal cord injury: evidence for a multiphasic inflammatory response in the acute to chronic environment.Brain2010,133:433–447.
32. De Castro RC Jr, Burns CL, McAdoo DJ, Romanic AM:Metalloproteinase increases in the injured rat spinal cord.NeuroReport2000,11:3551–3554.
33. Wells JE, Rice TK, Nuttall RK, Edwards DR, Zekki H, Rivest S, Yong VW:An adverse role for matrix metalloproteinase 12 after spinal cord injury in mice.J Neurosci2003,23:10107–10115.
34. Veeravalli KK, Dasari VR, Tsung AJ, Dinh DH, Gujrati M, Fassett D, Rao JS: Human umbilical cord blood stem cells upregulate matrix metalloproteinase-2 in rats after spinal cord injury.Neurobiol Dis2009, 36:200–212.
35. Leivonen SK, Chantry A, Hakkinen L, Han J, Kahari VM:Smad3 mediates transforming growth factor-beta-induced collagenase-3 (matrix metalloproteinase-13) expression in human gingival fibroblasts. Evidence for cross-talk between Smad3 and p38 signaling pathways.J Biol Chem
2002,277:46338–46346.
36. Kim ES, Kim MS, Moon A:TGF-beta-induced upregulation of MMP-2 and MMP-9 depends on p38 MAPK, but not ERK signaling in MCF10A human breast epithelial cells.Int J Oncol2004,25:1375–1382.
37. Cuadrado E, Rosell A, Penalba A, Slevin M, Alvarez-Sabin J, Ortega-Aznar A, Montaner J:Vascular MMP-9/TIMP-2 and neuronal MMP-10 up-regulation in human brain after stroke: a combined laser microdissection and protein array study.J Proteome Res2009,8:3191–3197.
doi:10.1186/1742-2094-9-122
Cite this article as:Lightet al.:Multiplex array proteomics detects increased MMP-8 in CSF after spinal cord injury.Journal of Neuroinflammation20129:122.
Submit your next manuscript to BioMed Central and take full advantage of:
• Convenient online submission
• Thorough peer review
• No space constraints or color figure charges
• Immediate publication on acceptance
• Inclusion in PubMed, CAS, Scopus and Google Scholar
• Research which is freely available for redistribution