Hmga2is required for canonical WNT signaling during lung development
Full text
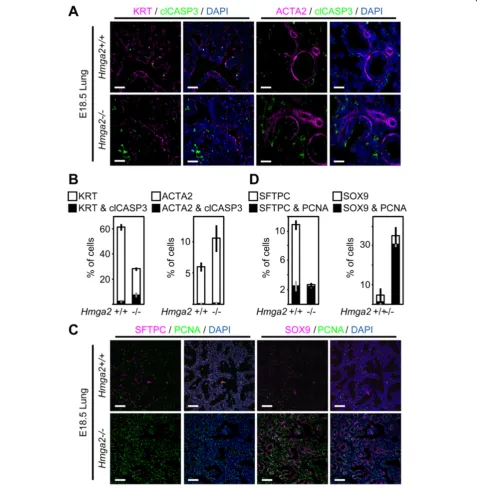
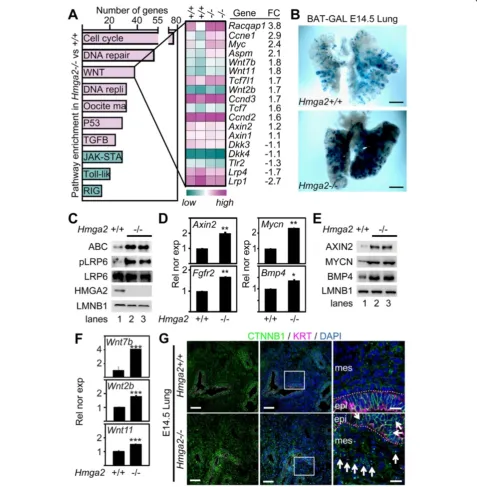
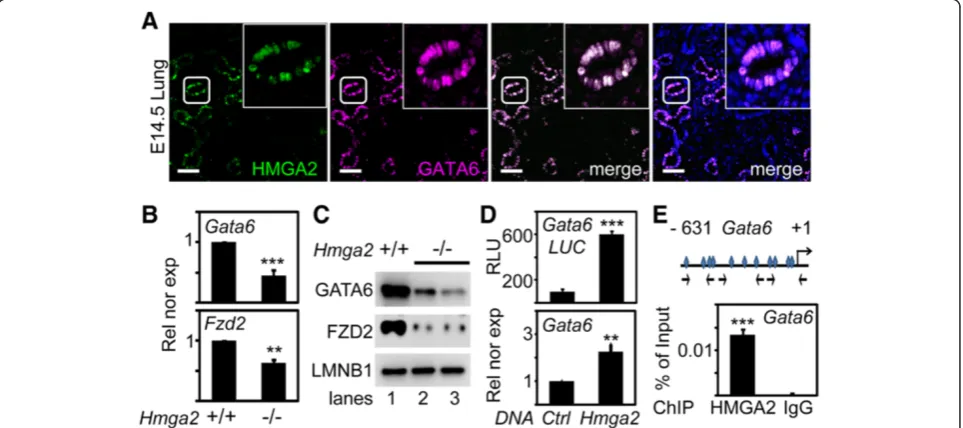
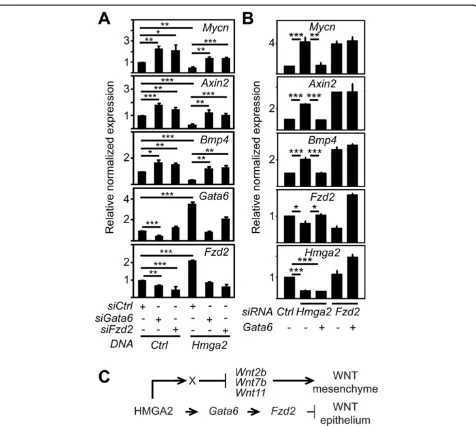
Figure



Related documents
H.pylori : Helicobacter pylori ; IEE: Image-enhanced endoscopy; LCI: Linked color imaging; LH-RH: Luteinizing hormone-releasing hormone; MALT: Mucosa- associated lymphoid tissue;
INSULATION COORDINATION Steve Lambert Insulation element and design factors to handle overvoltages (60 Hz, lightning, and switching surges) and respective insulation
• Be explicit about spillover effects when designing appraising projects (such as administrative burdens, skills constraints), for example by appraising the impact on a sector of
Conclusions and Recommendations From our Interviews with grain farmers and distributers, we learned that while there is no Suitable small-scale grain harvesting
investigating the effect of different parameters on the model reaction, we synthesize efficient route for tetrahydro benzo [ b ] pyran derivatives (4a-m), using
A meta-synthesis of existing qualitative research on early professional socialization and career choice in nursing has the potential to inform future research and
We hypothesized that an evidence-based PA intervention – GEhen, Spielen und Tanzen Als Lebenslange Tätigkeiten – kompakt [walking, playing and dancing as lifelong
