P
ATRIKF
ORSZT1, A
GNIESZKAP
ILECKA2, 3, M
AŁGORZATAM
AŁODOBRA2, 4,
J
OANNAM
ARKOWSKA3, K
RZYSZTOFM
AKSYMOWICZ3, T
ADEUSZD
OBOSZ2Single−Nucleotide Polymorphism Association Study
of
VDR
and
CDH1
Genes and the Risk of Prostate Cancer
Wpływ polimorfizmu typu SNP w genach
VDR
i
CDH1
na ryzyko rozwoju raka prostaty
1Regional Specialist Hospital, Research and Development Center, Wroclaw, Poland
2Department of Forensic Medicine, Molecular Technique Unit, Wroclaw Medical University, Wroclaw, Poland 3Department and Unit of Forensic Medicine, Wroclaw Medical University, Wroclaw, Poland
4Warsaw University of Medicine, Postgraduate School of Molecular Medicine, Warsaw, Poland
Adv Clin Exp Med 2009, 18, 3, 215–220 ISSN 1230−025X
ORIGINAL PAPERS
© Copyright by Wroclaw Medical University
Abstract
Background. Prostate cancer (PC) is considered the most common cause of male cancer mortality. A positive fam− ily history is one of the strongest risk factors for prostate cancer. Numerous data indicate that PC has a genetic background; however, it cannot be explained as a single−gene disease but as a multigenetic disorder.
Objectives. The aim of this study was to search for genetic correlation between single−nucleotide polymorphisms (SNPs) in the VDR and CDH1genes and the risk of PC.
Material and Methods. One hundred PC patients and 100 control subjects were investigated. The SNPs rs2107301 and rs2238135 in VDRand rs16260 in CDH1were detected by minisequencing followed by capillary electrophoresis. Hardy−Weinberg equilibrium, the chi−squared test, and non−parametric tests (Wald−Wolfowitz and Mann−Whitney U) were used for statistical analyses.
Results. Two of the three tested SNPs, i.e. rs2238135 in VDRand rs16260 in CDH1, displayed statistically sig− nificant differences in frequency between the two groups (p= 0.0266 and p= 0.0123 for rs2238135 and rs16260, respectively). The C/C genotype of rs2107301 in VDRgene positively correlated with increased prostrate−specific antigen (PSA) level (p= 0.0073).
Conclusions. The results provide unique data and show strong association between the tested SNPs in the VDR
and CDH1genes and malignancy and progression of prostate cancer (Adv Clin Exp Med 2009, 18, 3, 215–220).
Key words: prostate cancer, SNP, VDRgene, CDH1gene.
Streszczenie
Wprowadzenie. Rak prostaty (r.p.) jest obecnie jedną z najczęstszych przyczyn śmierci z powodu nowotworów wśród mężczyzn. Jednym z czynników ryzyka jest wywiad rodzinny w kierunku raka prostaty. Wiele danych prze− mawia za tym, iż r.p. ma podłoże genetyczne o wielogenowym charakterze dziedziczenia.
Cel pracy.Poszukiwanie korelacji między polimorfizmem typu SNP w genach VDRiCDH1a ryzykiem rozwoju raka prostaty.
Materiał i metody. Zbadano 100 pacjentów z rakiem prostaty oraz 100 zdrowych mężczyzn w podobnym przedziale wiekowym. Analizie poddano rs2107301 oraz rs2238135 znajdujące się w genie VDRoraz rs16260 w genie CDH1. Uzyskane wyniki poddano analizie statystycznej z zastosowaniem testu χ2oraz testów niepara−
metrycznych (Wald−Wolfowitza i U Mann−Whitneya).
Wyniki. Dwa z trzech analizowanych polimorfizmów: rs2238135 w genie VDRoraz rs16260 w genie CDH1
wykazało istotne statystycznie różnice w częstości występowania między badanymi grupami (p = 0,0266 i p = 0,0123 dla rs2238135 i rs16260, odpowiednio). Poza tym genotyp C/C rs2107301 dodatnio korelował z podwyższonym poziomem PSA u chorych (p = 0,0073).
Wnioski. W przestawionej pracy wykazano korelację między polimorfizmami w genach VDRiCDH1a rozwojem raka prostaty wśród pacjentów z regionu Dolnego Śląska (Adv Clin Exp Med 2009, 18, 3, 215–220).
Prostate cancer (PC) is considered, after lung cancer, the most common cause of male cancer mortality [1]. The mechanism of its pathogenesis is still unknown; however, the risk factors are well known, one of the strongest being a positive fami− ly history of prostate cancer. Approximately 10–15% of men with prostate cancer have at least one relative who is also affected. Steinberg et al. [2] estimated that the risk of prostate cancer rises by a factor of two for patients with a positive fam− ily history and by five with more than one affect− ed first−degree relatives. This suggests that PC has a genetic background; however, it cannot be explained as a single−gene disease, but rather as a multigenetic disorder [3, 4]. The list of candidate genes correlating with prostate cancer covers many, including genes implicated in testosterone biosynthesis, degradation, and distribution [5]. Cancer is defined as uncontrolled cell prolifera− tion; therefore genes controlling cell differentia− tion and proliferation were selected as candidate genes for PC in this study.
The role of vitamin D receptor (VDR) in cell proliferation and differentiation was established over 20 years ago [6]. There is strong evidence for correlation of variations in VDRgene with many types of cancer, including breast, renal, colon, and PC [6]. Some variations, such as single−nucleotide polymorphisms (SNPs) and STR polymorphisms, have been related to increased PC risk in numer− ous studies in various populations [7–11]; howev− er, the results provided discrete data. The E−cad− herin gene (CDH1), encoding a transmembrane glycoprotein which mediates cell−cell adhesion and signaling [12], has been recently associated with increased risk of PC. It has been shown that impaired expression of CDH1resulted in prostate cancer metastasis and progression and poor prog− nosis [13]. According to Hoogerdoorn et al. [14], SNPs in the promoter region might influence the level of expression. Changes in the promoter region of CDH1 can lead to a disruption of cell adhesion and signaling and finally to cancerogen− esis. Therefore we investigated SNP in the pro− moter region of CDH1gene to evaluate its associ− ation with prostate cancer.
The aim of this study was to search for genet− ic correlation between SNPs in the VDR and
CDH1 genes and the risk of prostate cancer in a Lower Silesian population. The results were also compared with those obtained by others authors to confirm or reject an association with PC.
Material and Methods
Study Population
SNP genotyping was performed on 100 patients diagnosed with PC treated at the Regional Specialist Hospital, Research and Development Center, Wroclaw. The mean age of the PC patients was 75 ± 7.6 years and the mean prostate−specific antigen (PSA) level was 40 ± 55 ng/ml. Genetic material of 100 control subjects came from the DNA bank of the Molecular Technique Unit, Wroclaw Medical University, representing a broad male population from the Lower Silesian region undergoing paternity tests. The cancer patients and control subjects were unrelated. The study was approved by the Ethics Committee and conformed to the ethical standards of the Declaration of Helsinki.
Genotyping
Genetic material of the PC patients was isolat− ed from whole blood samples using an E.Z.N.A Blood DNA Kit (Omega Bio Tek). The DNA of the control subjects was isolated using the Chelex method from blood spots. DNA was ampli− fied by multiplex PCR (Qiagen Multiplex MasterMix, Qiagen) using the GeneAmp PCR System 9700 (Applied Biosystems) under the fol− lowing conditions: 95°C for 15 min, followed by 32 cycles at 94°C for 30 s, 57°C for 90 s, and 72°C for 90 s, then 72°C for 10 min and 4°C there-after. The primers used in PCR are presented in Tab. 1.
The PCR products were evaluated by agarose gel electrophoresis. Six µl of product was run in 2% agarose gel in 0.5 TBE buffer. The remaining PCR product was purified of incorporated primers and dNTPs by digestion with a mixture of Exol (0.1 µl, 20 U/µl) and SAP (1 µl, 1 U/µl) (Fermen− tas) and a quantity of 0.5 µl was subjected to minisequencing. The minisequencing reaction (ABI Prism SNaPshot Multiplex Kit, Applied Bio− systems) was performed as a multiplexing reaction using the GeneAmp PCR System 9700 (Applied Biosystems) under the following conditions: 25 cycles at 96°C for 10 s, 50°C for 5 s, and 60°C for 30 s, then 4°C thereafter. The primer set used in the minisequencing reaction is shown in Tab. 2.
rs2238135) in the VDRgene located on intron 4 and intron 1, respectively, and one (rs16260) in the
CDH1 gene located in the promoter region were genotyped.
Statistical Analysis
The chi−squared test was used to evaluate the Hardy−Weinberg equilibrium (HWE) as well as the differences in genotype frequencies between the two groups. Non−parametric tests (Wald− Wolfowitz and Mann−Whitney U) were used to assess correlation between genotype and PSA level and Gleason Score of the PC patients. Statistical significance was defined as p < 0.05. The statistical analyses were performed with STA− TISTICA 8 (StatSoft). The classical linkage dise− quilibrium value (D) was computed for a two− locus haplotype using the formula:
D = hf – p x q
(hf – haplotype frequency, p and q – allele fre− quency).
Results
Study Population Analysis
No correlations were found between PSA level and GS (r= 0.1941), PC patient age and PSA level (r= –0.0282), or age and GS (r= 0.1576).
Genotyping Analysis
The main aim of this study was to assess dif− ferences in the frequencies of SNPs in the VDR
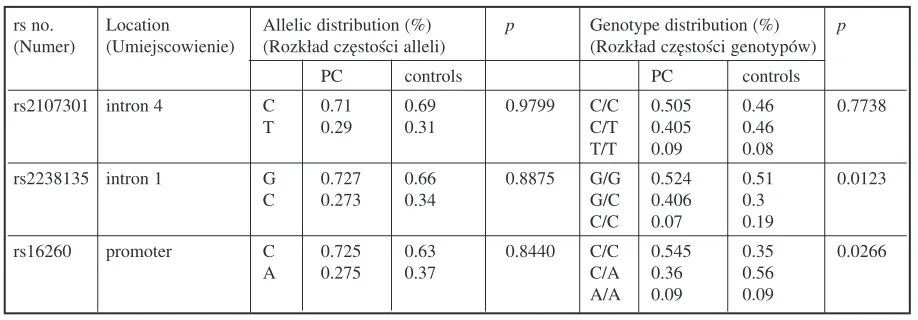
and CDH1genes. The genotyped SNPs showed no divergence from Hardy−Weinberg equilibrium (p= 0.9992, p= 0.9924, and p= 0.9988 for rs2107301, rs2238135, and rs16260, respectively). The rs2107301 SNP in VDRshowed no statistical dif− ference in genotype frequency (p= 0.7738); how− ever, there was a slight increase in the C/C geno− type in the PC patients, while in the control group the numbers of C/C homozygotes and C/T het− erozygotes were equal. The rs2238135 SNP fre− quency in VDRshowed statistical difference (p= 0.0266), with the C/C genotype dominating in the control subjects and the number of G/C heterozy− gotes considerably higher in the PC patients. The rs16260 SNP in CDH1also showed a difference in genotype frequency between the two groups (p= 0.0038), with the A/C genotype occurring more frequently in the controls and the C/C homozy− gotes prevalent in the PC patients. The genotype and allele frequencies are presented in Table 3.
SNP Association Analysis
The associations of particular genotypes with the clinical parameters of the PC patients were assessed using non−parametric tests (Wald− Wolfowitz and Mann−Whitney U). No statistical Table 1.Sequences of primers used in the PCR multiplex reaction
Tabela 1. Sekwencje primerów użytych w multipleksowej reakcji PCR
Gene SNP Primers sequences Product size (bp)
(Gen) (Sekwencje primerów) (Wielkość)
VDR rs2107301 (F) 5’ GTC TTG CAT GGG AAT AAC TTG 3’ 252
(R) 5’ GAT TGA ACC TAA GAT GTC ATT AC 3’
VDR rs2238135 (F) 5’(GACT)4CCA CCA CTT GCC TAG CTG T 3’ 75
(R) 5’(GACT)4AGT GAC TTA CCC AGG GTC C 3’
CDH1 rs16260 (F) 5’ CAA AAG AAC TCA GCC AAG TGT A 3’ 174
(R) 5’ (GACT)2CGG CCT CGC ATA GAC GCG 3’
Table 2.Sequences of primers used in the minisequencing reaction Tabela 2. Sekwencje primerów użytych w reakcji minisekwencjonowania
Gene SNP Primers sequences Product size (bp)
(Gen) (Sekwencje primerów) (Wielkość)
VDR rs2107301 (F) 5’ GTC TTG CAT GGG AAT AAC TTG 3’ C – 28, T – 29
(R) 5’ TTG CTG AGT GTG AAA TAA TTT TGC 3’ G – 29, A – 27 VDR rs2238135 (F) 5’(GACT)4CCA CCA CTT GCC TAG CTG T3’ C – 39.5, G – 38.5
(R) 5’(GACT)4 AGT GAC TTA CCC AGG GTC C3’ G – 38.6, C – 38.7
CDH1 rs16260 (F) 5’ GTC TTG CAT GGG AAT AAC TTG 3’ C – 31.5, A – 33
correlations between the SNPs and the age at diag− nosis were found. The C/C genotype of rs16260 in
CDH1gene positively correlated with the Gleason score (p = 0.0003); carriers of the C/C homozy− gote were characterized by increased GS. Statistically significant association was also found for the C/C genotype of rs2107301 in VDR and increased PSA level (p= 0.0073). Rs2238135 in the VDR gene did not display statistical associa− tion with PSA level or GS despite the fact this SNP showed a difference in frequency between the test− ed groups.
Linkage Disequilibrium:
Two−Locus Analysis
In two−locus analysis it was noted that the risk allele C/C of rs16260 was in linkage disequilibri− um with the C/C genotype of rs2238135 (D = 0.728). Moreover, the number of C/C genotype in carriers of both SNPs was significantly greater in the PC patients than in the healthy controls (p= 0.0141). All PC patients with the C/C genotype of rs2238135 in VDRwere carriers of the C/C geno− type of rs16260 in CDH1. The Wald−Wolfowitz (WW) and Mann−Whitney U(MW) tests showed no significant association of the tested two−locus haplotype with PSA level and GS (WW: p = 0.5896 and MW: p= 0.2167 for PSA and WW: p= 0.6183 and MW: p= 0.6859 for GS).
Discussion
The results presented here indicate that the genotyped single−nucleotide polymorphisms in the
VDRand CDH1 genes may increase the risk for PC, especially for G/G carriers of rs2238135 in the
VDR and the C/C genotype of rs16260 in the
CDH1 gene. Furthermore, the latter displayed
associations with increased Gleason score. The rs2107301 SNP in VDRdid not show significant differences in frequency; there was only a slight increase in the frequency of C/C carriers in the patient group. However, the carriers of this geno− type were characterized by higher PSA levels. Moreover, two−locus analysis showed linkage dis− equilibrium between the C/C genotype of rs2238135 and the C/C genotype of rs16260 in the PC patients. The number of C/C−C/C carriers of both SNPs was considerably higher in the patients than in the controls.
VDR Gene Polymorphism
SNPs in the VDRgene have been widely test− ed by numerous research groups [7–10, 15]. In the present study, two SNPs located on intron 1 (rs2238135) and intron 4 (rs2107301) were ana− lyzed. The results of rs2238135 obtained in this study are unique and contradict those of others. A study performed by Holick et al. [7] found sta− tistically significant differences in genotype fre− quency; however, the risk genotype was a rare homozygote. Unexpectedly, the present study showed that the G/G homozygote was the preva− lent genotype in the PC patients and the rare allele occurred more frequently among the healthy sub− jects. The results observed for rs2107301 also con− tradict those obtained by Holick et al. In genotyp− ing analysis, the present study found only a slight increase in homozygotes for the rare C/C genotype in the patient group as well as for the homozygote of the common G/G genotype, and the frequencies did not approach significance. Moon et al. obtained similar results, i.e. a lack of association between SNP rs2107301 and PC risk [8].
Furthermore, the present study found statisti− cally significant correlation between the C/C genotype of rs2107301 and increased PSA level. On the other hand, no correlation between Gleason Table 3. Allelic and genotype distributions of the analyzed SNPs in the two groups
Tabela 3. Rozkład częstości alleli i genotypów analizowanych SNP−ów wśród badanych grup
rs no. Location Allelic distribution (%) p Genotype distribution (%) p (Numer) (Umiejscowienie) (Rozkład częstości alleli) (Rozkład częstości genotypów)
PC controls PC controls
rs2107301 intron 4 C 0.71 0.69 0.9799 C/C 0.505 0.46 0.7738
T 0.29 0.31 C/T 0.405 0.46
T/T 0.09 0.08
rs2238135 intron 1 G 0.727 0.66 0.8875 G/G 0.524 0.51 0.0123
C 0.273 0.34 G/C 0.406 0.3
C/C 0.07 0.19
rs16260 promoter C 0.725 0.63 0.8440 C/C 0.545 0.35 0.0266
A 0.275 0.37 C/A 0.36 0.56
score and the age at diagnosis was found. Holick et al. [7] and Moor et al. [8] also showed a lack of association between SNP variants of the VDRgene and clinical parameters; however, the correlation with PSA level found in the present study repre− sents new data, not revealed by others.
CDH1 Gene Polymorphism
Since the CDH1gene has been associated with increased risk of PC, numerous SNPs in this gene have been analyzed [16–19]. In the present study the genotype frequencies as well as the association between the progression of prostate cancer and an SNP located in the promoter region of CDH1gene at the –160 locus upstream from the gene were analyzed. The results of this study contradict data obtained by other research groups. The at−risk genotype in the Lower Silesian population was the C/C variant of –160A>C (rs16260). The C/A het− erozygote dominated in healthy subjects, suggest− ing a protective role rather than an association with an increased risk of PC. As the numbers of A/A carriers were equal in the two groups, there was no correlation with PC, which also varies from results obtained in previous studies [16–18] indicating that the A allele represented a risk allele for prostate cancer. Bonilla et al. [16] screened the whole promoter region and detected 21 SNPs, of which the –160 C/A variant (rs16260) showed
higher association of the A allele with PC in European Americans. Jonsson et al. [17] obtained similar results, as did Lindström et al. [18], in which the A/A or heterozygous C/A variants cor− related with increased risk of PC, which diametri− cally differs from the results obtained in the pre− sented Lower Silesian population. The association between the A allele of –160C/A (rs16260) poly− morphism with increased risk of PC was con− firmed in a meta−analysis [19], especially in European and Asian populations. The present results decidedly demonstrate a protective role of the A/C genotype rather than correlation with increased risk. It should also be mentioned that the present study found an association between the C/C genotype and increased Gleason score, sug− gesting an influence on the progression of prostate cancer.
In conclusion, two of the three tested SNPs displayed statistically significant differences in frequency in patients with prostate cancer and in healthy controls. The results presented here diverge from those obtained by others, indicating disagreement about the association between these SNPs and prostate cancer. These differences might be due to the specific pattern of the Lower Silesian population or the coincidence of control and patient group selection. Further analyses must be performed to evaluate the role of the analyzed SNPs in the risk of prostate cancer.
References
[1] Jemal A, Siegel R, Ward E, Hao Y, Xu J, Murray T, Thun MJ:Cancer Statistic 2008. CA Cancer J Clin, 2008, 58, 71–96.
[2] Steinberg GD, Carter BS, Beaty TH, Childs B, Walsh PC:Family history and the risk of prostate cancer. Prostate 1990, 17, 337–347.
[3] Forrest MS, Edwards MS, Houlston R, Kote−Jarai Z, Key T, Allen N, Knowles MA, Turner F, Ardern−Jones A, Murkin A, Williams S, Oram R, Bishop DT, Eeles RA: Association between hormonal genetic polymor− phisms and early−onset prostate cancer. Prostate Cancer Prostatic Dis 2005, 8, 95–102.
[4] Schaid DJ:The complex genetic epidemiology of prostate cancer. Hum Mol Genet 2004, 13(1), 103–121.
[5] Gos M, Sadowska M, Wiechno P, Demkow T, Janik P:Tło genetyczne a ryzyko rozwoju raka prostaty oraz obraz kliniczny choroby. Urol Pol 2006, 59, Supl. 1.
[6] Thorne J, Campbell MJ:The vitamin D receptor in cancer. Proc Mutr Soc 2008, 67(2), 115–127.
[7] Holick CN, Stanfork JL, Kwon EM, Ostrander EA, Nejentsev S, Ulrike P:Comprehensive association analy− sis of the Vitamin D pathway genes, VDR, CYP27B1, and CYP24A1 in Prostate Cancer. Cancer Epidemiol Biomarkers Pre 2007, 16(10), 1990–1999.
[8] Moon S, Holle S, Bodiwala D, Luscombe CJ, French ME, Liu S, Saxby MF, Jones PW, Fryer AA, Strange R:Associations between G/A1229, A/G30875, C/T48200 and C/T65013 genotypes and haplotypes in the vitamin D receptor gene, ultraviolet radiation and susceptibility to prostate cancer. Ann Hum Gen 2006, 70, 226–236.
[9] Mikhak B, Humter DJ, Spiegelman D, Platz EA, Hollis BW, Giovannucci E:Vitamin D receptor (VDR) gene polymorphisms and haplotypes, interaction with plasma 25−hydroxvitamin D and 1,25−dihydroxyvitamin D and prostate cancer risk. Prostate 2007, 67(9), 911–923.
[10] Li H, Stampfer MJ, Hollis JB, Mucci LA, Gaziano JM. Hunter D, Giovannucci EL, Ma J:A prospective study of plasma vitamin D metabolites, vitamin D receptor polymorphisms and prostate cancer. Plos Med 2007, 3, e103.
[12] Grunwald GB:The structural and functional analysis of cadherin calcium−dependent cell adhesion molecules. Curr Opin Cell Biol 1993, 5, 797–805.
[13] Dunsmuer WD, Gillett SE, Meyer SC, Young NP, Corbishley C, Eeles RA Kirby RS:Molecular markers for predicting prostate cancer stage and survival. BJU Int 2000, 86, 869–878.
[14] Hoogendoorn B, Coleman SL, Guy CA, Smith K, Bowen T, Buckland PR:Functional analysis of human pro− moter polymorphisms. Hum Mol Genet 2003, 12, 2249–2254.
[15] Onen IH, Ekmekci A, Eroglu M, Konac E, Yesil S, Biri H:Association of genetic polymorphisms in vitamin d receptor gene and susceptibility to sporadic prostate cancer. Exp Biol Med 2008, 233, 1608–1614.
[16] Bonilla C, Mason T, Long L, Ahaghotu C, Chen W, Zhao A, Coulibaly A, Bennett F, Aiken W, Tullock T, Coard K, Freeman V, Kittles RA:E−cadherin polymorphisms and haplotypes influence risk for prostate cancer. Prostate 2006, 66(5), 545–556.
[17] Jonsson BA, Adami HO, Hähhlund M, Bergh A, Göransson I, Statin P, Wiklund F, Grönberg H: − 160C/A polymorphism in the E−cadherin gene promoter and risk of hereditary, familial and sporadic prostate can− cer. Int J Cancer 2004, 109(3), 348–352.
[18] Lindström S, Wiklund F, Jonsson BA, Adami HO, Bälter K, Brookes AJ, Xu J, Zheng SL, Isaacs WB, Adolfsson J, Grönberg H:Comprehensive genetic evaluation of common E−cadherin sequence variants and prostate cancer risk: strong confirmation of functional promoter SNP. Hum Genet 2005, 118, 339–347.
[19] Qiu LX, Li RT, Zhang JB, Zhong WZ, Bai JL, Liu BR, Zheng MH, Qian, XP:The E−cadherin (CDH1) −160 C/A polymorphism and prostate cancer risk: a meta−analysis. Eur J Hum Genet 2008 [EPub ahead of print].
Address for correspondence:
Małgorzata MałodobraWroclaw Medical University Molecular Technique Unit Marii Sklodowskiej−Curie 52 50−369 Wroclaw
Poland
Tel.: +48 71−7841139
E−mail: [email protected]
Conflict of interest: None declared