Copyright 0 1995 by the Genetics Society of America
Mutations that Alter the
Timing
and
Pattern of
cubitus interruptus
Gene
Expression
in
Drosophila melanogush
Diane
C.
Sl~sarski,”~
Cynthia
Kelsey Motzny‘ and Robert Holmgren
Department of Biochemisty, Molecular Biology and Cell Biology, Northwestern University, Evanston, Illinois 60208-3500
Manuscript received June 8, 1994
Accepted for publication September 15, 1994
ABSTRACT
The cubitus intenuptus ( c i ) gene is a member of the Drosophila segment polarity gene family and
encodes a protein with a zinc finger domain homologous to the vertebrate Gli genes and the nematode
tra-l gene. Three classes of existing mutations in the d locus alter the regulation of ci expression and can
be used to examine ci function during development. The first class of cimutations causes interruptions in
wing veins four and five due to inappropriate expression of the ci product in the posterior compartment
of imaginal discs. The second class of mutations eliminates ci protein early in embryogenesis and causes
the deletion of structures that are derived from the region including and adjacent to the engrailed
expressing cells. The third class of mutations eliminates ci protein later in embryogenesis and blocks
the formation of the ventral naked cuticle. The loss of d expression at these two different stages
in embryonic development correlates with the subsequent elimination of wingless expression. Adults
heterozygous for the unique
ci“
mutation have deletions between wing veins three and four. A similarwing defect is present in animals mutant for the segment polarity gene fused that encodes a putative
serine/threonine kinase. In cia/+ and fused mutants, the deletions between wing veins three and four
correlate with increased ci protein levels in the anterior compartment. Thus, proper regulation of both
the ci mRNA and protein appears to be critical for normal Drosophila development.
D
URING Drosophila embryogenesis the antero-pos- terior body axis is divided into a series of seg- ments that are the basic units for establishing the pat- tern of structures within the developing embryo. The segment polarity genes play a critical role in specifymgpattern along the antero-posterior axis of each segment
(NUSSLEIN-VOLHARD and WIESCHAUS 1980). The initial
expression of several segment polarity genes is con-
trolled by pair rule genes, but continued expression
depends on interactions among the segment polarity
genes and their protein products (reviewed in INCHAM
1991; HOOPER and Scorn 1992).
Central to the pattern formation process is the main-
tenance of wingless (wg) gene expression in the cells
just anterior to the parasegmental boundary (MARTI-
NEZ-ARIAZ and LAWRENCE 1985; BAKER 1987, 1988; VAN
DEN HEUVEL et al. 1989). wgencodes a Drosophila mem-
ber of the Wnt family of secreted growth factors and
may act as a morphogen to establish pattern within
segments [VAN DEN HEUVEL et al. 1989; BEJSOVEC and
MARTINEZ-ARIAS 1991; GONZALEZ et al. 1991; BEJSOVEC
and WIESCHAUS 1993; SAMPEDRO et al. 1993 (who sug-
Corresponding author: Robert Holmgren, Department of Biochemis- try, Molecular Biology and Cell Biology, Northwestern University,
2153 Sheridan Rd., Evanston, IL 60208-3500,
E-mail: [email protected]
Baltimore, MD 21218-2685.
’
Present address: Department of Biology, Johns Hopkins University,Co-first authors.
Genetics 139: 229-240 (Januav, 1995)
gest that wg is not a morphogen)]. In animals lacking
the wg protein, most of the pattern elements within a
segment are eliminated and replaced with a lawn of
denticles. In animals deficient for the ci region of the
fourth chromosome, wg expression in the epidermis is
eliminated during germ band extension (HIDALGO and
INCHAM 1990; C. MOTZNY and R. HOLMGREN, u n p u b lished results) and the ventral cuticle is covered with
denticles (NUSSLEIN-VOLHARD and WIESCHAUS 1980;
ORENIC et al. 1987).
The cubitus i n t m p u s ( c i ) region of the fourth chro-
mosome contains three genetically distinct classes of
mutations with a complex pattern of complementation
(Table 1) (HOCHMAN 1973; ORENIC et al. 1987). In this
work it is shown that all of these mutations alter the expression of a single transcription unit, which is re- ferred to as ci. Conceptual translation of the ci transcript
predicts a protein with five zinc fingers (ORENIC et al.
1990). The zinc finger domain has a high degree of
amino acid sequence identity with the vertebrate Gli
genes (84-93%) (KINZLER et al. 1987; RUPPERT et al.
1990) and a lower degree of identity with the tra-1 gene
of Caenorhabditis elegans (63%) (ZARKOWER and HODG
KIN 1992). Because ci was the first mutation to be iso- lated in this locus, we have given all of the mutations ci allele designations.
The
ci’
class of mutations causes breaks in the fourthand fifth wing veins and includes the recessive mutation
230 D. C. Slusarski, C. Kelsey Motzny and R. Holmgren
TABLE 1
Classes of ci mutations
Complementation
Class Phenotype properties
ci'
Disruptions in wing Complements the veins 4 and 5 segmentation defectsof the ~ 2 " ~ " ~ and
d1(4)17
classes
#) 13
Deletion of ventral Complements the naked cuticle in segmentation defects embryos of the ciL(4j17 class
and the wing vein defects of the
ci'
class&4j 1 7 Deletion of the first row Complements the
of ventral denticles segmentation defects and variable deletions of the ~ i ' ( ~ j ' j class of ventral naked and the wing vein
cuticle defects of the ci'
class
mutation ci".
c
'
i
is an unusual allele because it is alsoa member of the c2zf41z3 class of mutations. This class
includes the
c
'
i
and l(4) 13 ( ci1f4113) mutations and causesrecessive segmentation defects in which the ventral na- ked cuticle of each segment is deleted and replaced with a mirror image duplication of the denticle belt. The ci"4'17cla~~contains the
Cz
(ci") and Z ( 4 ) 1 7 ( ~ i ' ( ~ ) . ' ~ )mutations. These mutants also have recessive segmenta-
tion defects. In ~ i ' ' ~ ) . ' ' mutants, the first row of denticles is removed from the abdominal segments, and there
are variable deletions of naked cuticle. The ci" mutants
are more extreme, and the naked cuticle is removed in
every segment. The ci" mutation also causes variable
dominant adult defects, including partial fusions be-
tween wing veins three and four and the deletion of
the ocelli.
The mutations were placed into these three groups
on the basis of their complementation properties.
~ i " ~ ' . ' ~
mutations fail to complement the ci" recessive segmen-
tation defect but complement the ci.' wing vein defect.
Mutations in the ciLf4"jr class and the ~i"".'~ class comple-
ment each other's segmentation defect and are viable in certain combinations. Deletions of the locus have a
phenotype similar to that of cia and fail to complement
both the and ci1'4'17 classes.
Here it is shown that mutations in the ci locus cause
a variety of alterations in the expression of the ci pro-
tein. The two classes of embryonic mutations eliminate
expression of the ci protein at different developmental
stages. The adult wing vein defects of the
ci'
class muta-tions are caused by misexpression of the ci protein in
the posterior compartment of imaginal discs. The
tic'
mutation produces a truncated ci protein with an al- tered distribution pattern in the anterior compartment.
The distribution of the ci protein in
ci""/+
mutantimaginal discs is similar to that of the wild-type ci pro- tein in imaginal discs mutant for the putative serine/
threonine kinase gene fused
vu)
(PREAT et al. 1990).MATERIALS AND METHODS
Stocks: The ci" and alleles were obtained from B. HOCHMAN, the ciu, Mhz<
ci'
and ciw alleles were provided by the Bowling Green Drosophila stock center, the tip'"" allele was provided by T. KORNBERG and the su(Hd) and su(Hw")alleles were from V. CORCES. The gamma-ray revertant
gRS0
is a partial revertant of c i D (ORENIC et al. 1987).fu" is a strong
EMS mutation of f u generated by R. HOLMGREN.
Generating c i P h revertants: y q tip'"' females were crossed to P[q+A2-3] Sb/TM3; spaFi males. y q P[q+A2-3] S b / + ; ciphc/spupoL males were crossed to yw, spaPo' females. Each indi- vidual excision event was crossed to yw; s p P l flies and the y q sPaP"'+/sPuP0' sibling progeny crossed to identify both embry- onic and adult mutant phenotypes. Each lethal line was char- acterized by examining cuticle pheno pes and by testing complementation with ci", ci1(4'17 and ci
2
.Mapping ci mutants: Mutations were mapped by per- forming a series of restriction digests with DNA isolated from appropriate mutants, blotting the gel fractionated fragments onto nitrocellulose and probing with various cloned frag- ments that spanned the ci gene region. Fragments from ci cDNA clones were particularly useful because they did not contain repetitive sequences that are present in flanking ge- nomic regions.
Cuticle preparations: The procedure of VAN DER MEER (1977) was used. The cuticles were mounted in a 1:l solution of Hoyers:lactic acid and photographed with phase-contrast microscopy.
Wing blade preparations: Wings were removed, dehy- drated with propanol and mounted in euparal.
Antibody stainings: Antibody staining of embryos was per- formed according to PATEL et al. (1989) using the anti-ci rat monoclonal antibody 2A1 (C. MOTZNY and R. HOLMGREN, unpublished data), the anti-engrailed (en) mouse monclonal antibody 4D9 (gift from N. PATEL) and an anti-wg rabbit anti- body (gift from R. NUSSE). With ci" it is possible to unambigu- ously identify mutant embryos because the ci" allele is viable and fertile over ciplnc. Progeny from c i D / c i p l " " parents were double labeled with anti-ci antibodies and anti-@galactosidase antibodies; embryos that fail to stain with the anti-p-galactosi- dase antibody are homozygous for the
c
'
i
mutation. The other ci embryonic lethal mutations are not fertile over tip"'. Mutant embryos were identified by the distinct staining patterns pres- ent in approximately one quarter of the embryos. For the anti-en stainings, the ci" mutant embryos were identified by double labeling with anti-ci antibodies. Embryos were staged according to CAMPOSORTECA and HARTENSTEIN (1985).Imaginal discs were stained in a similar manner. Wandering third instar larvae were collected, rinsed in BSS (CHm and GEHNNG 1971), cut in half and turned inside out. Animals were fixed for 20 min in phosphate-buffered saline containing
2% paraformaldehyde. Stained discs were removed from the larval carcass and dehydrated for mounting.
In situ hybridizations: In situ hybridization to ci mRNA was performed using DIGlabeled RNA probes (TAUTZ and PFEIFLE 1989).
Regulation of ci Expression 23 1
were fractionated on a 7.5% acrylamide gel and blotted onto nitrocellulose. Protein levels were examined by staining the
nitrocellulose with Ponceau S solution (Serva). ci protein was
visualized using the 2A1 monoclonal antibody, an HRPcou-
pled goat anti-rat IgG secondary (Sigma) and the electro-
chemiluminescence (ECL) detection system from Amersham.
RNA analysis: Late third instar larvae were homogenized
in an SDSurea solution, phenol extracted and ethanol precip
itated (MCKENZIE et al. 1975). PolyA+ RNAwas selected using
oligo dT cellulose
.
Equal amounts of RNA were loaded ontoa formaldehyde gel. After blotting, the RNAs were probed
with the 5' end of a n' cDNA. The hybridization intensities
were quantified relative to an rp49 standard using a Molecular
Dynamics phosphorimager.
RESULTS
Molecular analysis of the
ci',
ciw, c i D ,dLC
and c i C emutations: Genetic analysis of the
ci',
c i D andcite
muta-tions did not distinguish whether they represented dis- tinct genes or partially complementing alleles of a sin- gle locus. Previous work showed that each of these
mutations is associated with alterations in a 6-kb BglII
fragment that contains the 5' end of the ci transcript
(ORENIC et al. 1990) (Figure 1). These alterations could be due to polymorphisms, but the lack of recombina- tion on the fourth chromosome makes that possibility
unlikely (BERRY et al. 1991). Therefore, we mapped
these mutations in more detail and assessed changes in
the expression pattern of the ci protein.
Sequences
5'
to the ci transcript were convenientlydivided into four regions using the restriction enzymes
Nsd and BglII (Figure 1). The
ci'
mutation is caused bythe insertion of a gypsy element into region 3. The ci"
mutation is associated with the insertion of foreign DNA
into region
2.
The insertion appears to be an I element(FINNEGAN 1989), because an I element probe hybrid-
izes in situ to the ci region (101F) in
ci"
mutant polytenechromosomes and region 2 DNA hybridizes to a band
that comigrates with an I element containing fragment
from
tic" mutants (data not shown). The
c i D mutationresults from a small inversion (ORENIC et al. 1990). We
have mapped the breakpoints to two regions: region 1
and 10 kb 3' to the ci transcription unit (not shown in
Figure 1). The breakpoint is represented by a dashed
line because the exact position has not been deter-
mined. The spontaneous mutation c i w also has alter-
ations in these same two regions, but the nature of the
alterations has not been characterized. The ciphc line
contains an insertion of a whitei (w') enhancer-trap
(EATON and KORNBERC 1990) into region 3.
Analysis
of ci protein expression in ci mutant em- bryos: Expression of the cigene was assessed in mutantsusing an anti-ci rat monoclonal antibody, 2A1 (C.
MOTZNY and R. HOLMGREN, unpublished results). The
antibody is specific to the ci protein product because
there is no labeling of A f Z f mutant embryos in which
the ci region is deleted (data not shown).
-
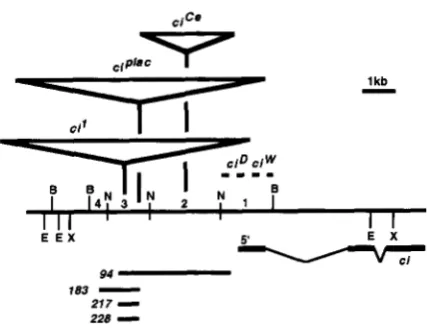
1 kbE E X 5' E X
v
E l94
183
-
217
-
228
-
FIGURE 1.-Map of the 5' end of the ci gene. The long
horizontal line represents the DNA from the 5' region of the
ci gene. The 6kb BgnI (B) fragment has been divided into
four segments using the restriction enzyme NstI (N). The
locations of insertions (V) and DNA alterations (-
-
-) associ- ated with each mutation are indicated. The positions of these aberrations have been determined by Southern blot analysisand are mapped to the resolution of the restriction map (with
the exception of the c i P l n C insertion, which has been precisely
mapped). The exon pattern of the ci transcript is shown below
the map. The 5' end corresponds to the end of the longest
ci cDNA clone. The extent of deletion mutations generated by imprecise excision of the ciPlac element is also shown below
the map of the region. The endpoints of the excision events
were determined by probing Southern blots of DNA from the
appropriate mutants and are mapped relative to the indicated
restriction sites. The distal break in mutation 183 is in frag- ment 4, whereas the distal breaks for deletions 94, 21 7 and
228are all within fragment 3. Deletions 183, 21 7and 228end
within the element, whereas deletion 94 eliminates the entire
P element and has a proximal breakpoint in fragment 1. The
extent of the deletions within the P element have not been
indicated on this map. The NsiI sites are only shown within
the 6kb BgZII fragment. B, BgZII; E, EcoRI; N, NsiI; X, XhoI.
In wild-type animals, expression of the ci protein initi-
ates during stage
5
and rapidly becomes expressedthroughout the embryo. During stage 10 ci mRNA is
eliminated from the posterior compartment of each
segment (EATON and KORNBERG 1990; ORENIC et al.
1990) and there is a corresponding decay of the protein
(Figure 2B). During stage 11 the wide stripe of ci pro-
tein expression begins to split, with higher levels of
antibody staining on the margins of the stripe and lower
level staining in the middle (Figure 2C).
In
ci"
mutants, low level uniform expression of ciprotein is observed at stage 5 and this expression peaks
at or shortly after gastrulation (Figure 2D). With the initiation of germ band extension, the ci protein level begins to decrease relative to wild type (Figure 2E; com- pare the mutant embryo in the center with the adjacent
wild type siblings) and is barely visible by the end of
stage 11 (Figure 2F).
mutant embryos fail to express the
ci protein until stage 7, but as development proceeds,
232 D. C. Slusarski, C. Kelsey Motzny and R. Holmgren
C
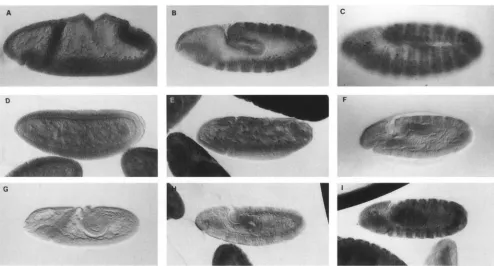
FIGURE 2.-Pattern of ci protein expression in embryonic development. Anterior is to the left and dorsal is up. Embryos were
stained with the 2A1 rat antici monoclonal. (A-C) Stages 8, 10 and 11 wild-type embryos. Initial expression of ci protein is
uniform throughout the segment; during stage 10 ci expression is eliminated in the en expressing cells and during stage 11 the
cells bracketing the m expressing cells show a higher level of antici antibodv labeling. (D-F) Stages 7, 8 and early stage 11 n'"
mutant embryos. The stage 7 embryo shows uniform expression of ci protein though the levels are somewhat lower than wild
type. With the initiation of germ band extension, protein levels begin to decrease, and bv stage 11 staining for the ci protein
has decreased to nearly background levels. (The embryo collections were specifically overstained to allow visualization of the
low ci protein levels in the mutants.) ( G I ) Stages 8, 10 and early stage 11 c;''~''' mutant embryos. At stage 7 expression of the
ci protein is not detected, but during stage 8 cells begin expressing the ci protein, and by stage 11 the pattern of expression,
though not wild type, is quite robust.
clusters of cells initiate ci protein expression. During
stage 10 (Figure 2H), the clusters begin to coalesce
into stripes, a n d by stage 11 the pattern of expression
approximates that of wild type, but at lower protein
levels (Figure 21). A distinction between the pattern of
ci expression in
cif'
and~i"~"'
embryos is observed instage 11 embryos. In ci'(4)f7embryos the wide stripe splits
as it does in wild type. In ci" mutants the stripe remains
uniform (data not shown). In ci"/cirM embryos the pat-
tern of ci protein expression is a combination of the
two mutant patterns. Athough the level of protein ex-
pression is lower than that of wild-type, ci protein is
expressed throughout embryonic development and the
pattern resembles that of wild-type embryos.
In
n"
and c i " ' mutants, which generally have normalembryonic development, the ci protein continues to
be expressed throughout the segment and fails to be
repressed in the posterior compartment (Figure 3, A
and
R).
Expression of wg
and
en in ci mutant embryos: T h econsequences of eliminating ci protein expression a t
different times in development were assayed by follow- ing the patterns of wg and en protein expression. In
ci"
mutants, expression of the wg protein decays duringstage 12 (Figure
a),
whereas expression of t h e e npro-
tein appears normal (data not shown). By stage 10,
wg
protein expression is eliminated in regions of the ven-
tral epidermis of ciN4)" mutant embryos (Figure 4B).
In these mutants, ci protein levels rise during stages 10
a n d 11 and epidermal expression of the wg protein
returns in late stage 12 (Figure 4C). In c i N 4 ) ' i mutants,
en protein expression is normal through stage 10 but
during stages 11 a n d 12 breaks in the stripes of e n
expression become evident (Figure 4D).
Generation of deletion mutations withii the ci
pro-
moter region: To define the promoter elements re-
quired for proper n'expression, we generated deletions
of a marked P element insertion. This approach seemed
particularly appropriate given the complex nature of the existing ci alleles. T h e
n'/''"'
line carries a zu" en-hancer trap and has a normal pattern of ci protein
expression during embryogenesis. Excision events were
induced by crossing this line to P[ly'A2-3](99R), a P
element transposase expressing strain (ROBERTSON d
Regulation of ci Expression 233
patterns of these mutations placed them in the ci"""
group (Figure 5). On the ventral surface the first row
of denticles is missing from most o f the abdominal seg-
ments, and there are variable deletions of naked cuticle
including the Keilin's organs of the thoracic segments
(Figure 5, C and D). Dorsally the isolated row of large
triangular hairs (visible in abdominal segments 2-8,
see arrowhead in Figure 5E) and flanking naked cuticle
are eliminated from most segments, and there is a cor- responding expansion of the region of socketed hairs
and fine hairs (Figure 5, G and H ) . T h e molecular
nature of the excision events was examined by Southern blot analysis and the approximate extent of the dele-
tions mapped (Figure 1). In all cases sequences around
the distal inverted-repeat of the transposable element
were altered.
ci protein expression in imaginal discs: Two types of
wing vein defects are observed in a' mutants. Mutations
of the
n"
class all cause disruptions in veins 4 and 5. Inn'"/+,
n", cri"'andci'"''"/+
mutants (h""5''is a revertantof b"), we find defects ranging from minor interrup-
tions of vein 5 to fusions of veins 4 and 5 (Figure 6,
R-
FIGLIRE S.-ci expression in n" and ci'"mr1tants. Anterior E).
ci';./+
mutants have deletions between veins 3 andis to the left and dorsal is up. (A) Stage 10
n"
mutant embryo. 4 (Figure 6F). The ci protein distrihutions in these mu- (€3) Stage 1 1 ci"' mutant embryo. In both mutants ci proteinexpression persists in the posterior compartment. tants were examined in the developing imaginal discs.
In normal imaginal development, ci transcript and pro-
tein are restricted to the anterior compartment (Figure
Six independent embryonic lethal mutations were iso-
7,
A and B). T h e ci mRNA is uniformly expressed in thelated. The nature of the segmentation defects, the pat- anterior compartment (Figure 7A), whereas antibody
terns of ci protein expression and the complementation labeling of the ci protein is elevated along the compart-
FICCRE 4.-Expression of en and wg proteins in ci mutants. Anterior is to the left. In a stage 12 Yj" mutant embryo, expression
of wg is fading (A). In n"'.""animals, wg expression begins to be eliminated in the epidermis (*) bv stage I O (B), but expression
234 D. C . Slusarski, C. Kelsey M o t m y a n d R. Holmgren
Regulation of n' Expression 2.73
x.
. ..-
'8"-
. " .
. _._- ~. - . . -
em.+,/
.
- _."_ . -
F1c.t IU;. (i.--M'ing defects in ci mrmnts. Anterior is up. (A) A wild-type wing. From antcrior t o posterior t h r veins are n a m h c w d 1 through 3 , o f ~ v h i c h 3 t o .5 arc indic;ltctl. The I X ) U I I ~ ; I I ~ separating t h c anterior ;mcl posterior coI111);1rtl?lrnts runs.just anterior
t o \ving vein 4. 111 r i " / + , ci', c.i"md d ' " 5 "
/+
mutants there is a range o f vein t I c f c ~ t s in the posterior conl MI tmcnt. I n c.i"/+ mutants. the f o u r t h and filih veins do not reach t h c margin o f t h c wing black ( B ) . n" at 18' ( C ) anti r i h ' ( h ) have :I 111019' estrclne phrnotye i n which w i n s 4 and .5 fuse t o make a singlr w i n t h a t angles toward t h c posterior margin o f ' the wing. vi""5"/+
(E) is a g ; ~ m n ~ : ~ - ~ x y rcvcrtant o f ri" ;and h a s t h c we;tkcst m u t a n t phenotype in which thcrc is a slight tliswption in vein 3. <if"/+
n l ~ ~ t ; ~ n t s h:~vc a diffcwmt alteration that consists of deletions hecwccn w i n s 3 and 4 (F).ment boundary (Figure 7B). I n c i ' / c i ' , e?'/+, r i L ' / +
and r i " / r i " ' mutants, inappropriate expression o f the
ci pt-otein is found i n the posterior compartmrnt (Fig-
~11-e
7,
C and D ) . T h e level o f inappropriate expressioncorrelates well with the extent of the wing vein disrup-
tions. r?"'5"/+ has the lowest level of ectopic expression (data not shown) and the least extreme defect. n " ' j c i " '
mutants have the highest level of ectopic expression
and the most extreme mutant phenotype. The c i ' / r i '
phenotype is temperature sensitive and more extreme
at lower temperatures; a correspondingly higher lrvel
of ectopic ci expression is obsened a t lower tempera-
tures. A s mentioned above, the ci' mutation is caused
by the insertion of ;I p p s y element. A number of muta-
tions caused by the insertion of gypsy elements can be
suppressed by mutations in the S I ~ / I / I ~ P . S . S ~ ~
or
Huily roing( n l (H70)) gene (reviewed in COK(:I<S and G I W X 1991 ).
In .su(HuJ);
n"
animals the wing vein defect is sup-pressed, and the ectopic expression ofci protein is rlirn-
inated (data not shown).
mRNA was isolated from ri' mrltants and . s ? r ( I f 7 ~ ) ; r i '
mlltants at third instar, pupal and adult stages. The
mRNA was fractionated by gel electrophoresis, blotted
onto nitrocellulose and probed for the ci transcript.
T h e level of the ei transcript was quantified o n a phos-
phorimager relative t o an r/)49 svantlard ( O ' < : O S X ~ . I . I .
and ROSBASII 1984). At the third instar l a ~ ~ a l stage, but
not in pupae or adults, the level of the ci transcript i n
ri' mutants is increased IO-fold relative t o that in
SN(H~O);
ri' mutants (data n o t shown). The size of theci transcript in these mutant backgrounds is i t l c ~ ~ t i ~ ; ~ l
to that ofwild-type, arguing that the ei' mutation causrs
higher levels o f expression from the normal c.i pro-
236 D. C. Slusarski, C . Kelsev M o t m y and R. Holmgren
- B
C'
-
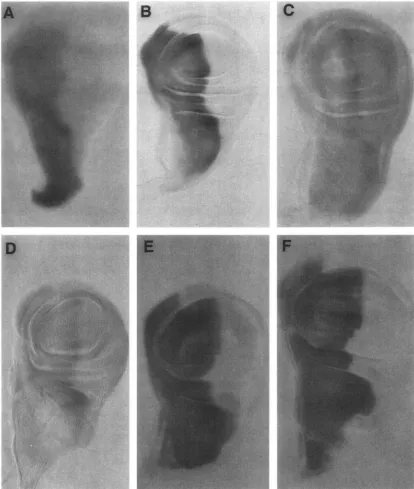
FIGURE 7.-ci expression in wing imaginal discs. Ventral is up and anterior is to thc Icft. I n wandering third instar larvae ci
mRNA is restricted to the anterior compartment (A). ci protein is also restricted to the anterior compartment of imaginal discs, but a4jacent to the compartment borlndary the level of antibody staining is greater ( R ) . In homozygous n"" mutants (C) and
n"'/+
mutants (D) expression of the ci protein is observed in the posterior compartment. The amount of ectopic ci protein correlates with the severity of the mutant phenotype. The highest level of ectopic expression is seen in homozygous n'"'mutants, which have the most severe defects. In d'/+ mutants (E), the level of antibody staining in the anterior Compartment is much higher than normal, and a distinction in levels between the region along the compartment boundary and the rest of the compartment is not seen. In Jc mutants (F) there is a similar change in the pattern of antibody staining to the ci protein; higher levels are seen across the entire segment.In
CY''/+
imaginal discs, the ci protein is properly body staining is elevated throughout the anterior com-restricted to the anterior compartment, but its level of partment (Figure
7E).
Regulation of n' Expression 237
A
B
C
D
E
-
b-
-
-
-
FIGCRE 8.-ci protein isolated from wing imaginal discs. Wild-type imaginal discs have a low level of ci protein (lane
A). In j l l mutants, ci protein levels are higher (lane R). In
n'"/+ the wild-type protein band 180 X 10'' M, can be seen
aswell as a mutant form at -120 X IO3 M, (lane C). The level of the mutant form is higher than that of the wild type.
In n'"/+ mutants the wild-typr form can be seen as well a s a
hint of a higher molecular weight form (lane D). A longer exposure shows that the ci protcin generated by the n " ) allcle
is -20 X 10'' M, larger than wild-type (lane E). The staining levels with ECL. detection give a general indication of relative protein concentrations hut the response is not linear and accentuates dimerences. Positions of molecular weight mark- ers are indicatrd a1 the left; sizcs correspond to 200, 116, 97,
66 and 45 (xIO" M , ) from top to bottom.
wing veins three and four. To determine whether there is a corresponding increase of ci protein in Jit mutant discs, we stained discs with antibodies for ci protein. A.
shown in Figure 7F, the level and distribution of the ci protein in J I L discs is nearly identical to
ci"/+
discs.c i D and cirr mutations are associated with the genera-
tion of altered protein products: Western blotting was
used to examine the protein products of the ci locus from wild-type, )L,
d'/+
andd'/+
imaginal discs.Equal amount5 of protein were loaded onto an SDS polyacrylamide gel, blotted onto nitrocellulose and probed with the 'LA1 antibody. Figure 8. A and B, shows that the level of ci protein is greatly increased in J ~ L
mutants. In kc'/+ discs the wild-type protein band of
180 X 10'
M,
is present along with a second band at-120 X IO3
M,
(Figure 8C). T h e lower band is presentat much higher levels than the wild-type band and pre- sumably corresponds to a truncated ci protein product from the n'" gene. In
ci"/+
discs, a longer exposure (Figure 8E) shows a protein band -20 X 10"M,
larger than the wild-type protein. This product presumably corresponds to a fusion protein generated by the ci"inversion.
DISCUSSION
&I,
C;", C;",
me and CIJV)I7 are allelic and alter theexpression pattern of the ci protein: We have shown
that the h1, n'\!: n./>, ,.ti and c j / l q l 1 7 mutations all alter
the expression of the same transcription unit and are associated with DNA alterations that map at or 5' to the start of transcription. T h e observed alterations are most likely the cause of the corresponding mutations because polymorphisms are very rare on the fourth chromosome. This is a consequence of the lack of re- combination that causes the fourth chromosome to be selected as a single genetic unit (BERRY Pt nl. 1991).
Natural Drosophiln mplnnofinstprpoprllations appear to be isogenic for the fourth chromosome. Therefore, anv polymorphism present on mutant fourth chromosomes would have to have been generated since the initial isolation of that parental laboratory stock.
ci function is required to maintain wg protein ex-
pression: Each mutant phenotype can be explained bv
an alteration in the expression of the ci transcription unit. T h e embryonic defects are recessive and result from a loss of ci protein expression. T h e n5(4'1i class of mutations fail to express the ci protein through early germ band extension. Loss of early ci expression results in the elimination of wg protein expression in the ven- tral epidermis during stage 10, and presumably as a
consequence, causes the variable loss of en protein ex- pression during stage 11 (BF.ISO\'EC and MARTINEZ- ARIAS 1991; HEKMSKKRK ~l nl. 1991). The consequences of these alterations include the elimination of the struc- tures derived from the m expressing cells (HAMA ~t nl.
1990) and of structures derived from either side of the m expressing domain. Deletions anterior to the m ex- pressing domain include the ventral black dots of the thoracic segments and deletions posterior to the pn ex-
pressing domain include the region of dorsal naked cuticle that follows the isolated row of triangular hairs.
late stage 12 and takes part in the formation of ventral naked cuticle (DFJSOVKC and MARTINEEARIA. 1991).
Elimination of r q f ~ ~ n c t i o n t h r o u g h stage 11 by temper- ature shifting a 71!g"allele from the nonpermissive to the permissive temperature 6 hr after egg laying generates a
phenotype similar to that of ci""li. T h e first row of
ventral denticles is eliminated and breaks in the stripes
of en protein expression are observed by stage 1 1 (R?l-
sow.<:
and MARTINEZ-ARIAS 1991). The phenotype of on the dorsal surface resembles that of tempera- ture sensitive h ~ d g ~ h o g (hit) mutants that have been shifted to the nonpermissive temperature during stage1 1 ( HEEMSKERK and DINARDO 1994). This suggests that just as m expression is disrupted in stage I 1 n'""" mu- tants, there may be a corresponding change in hh ex- pression.
In
n"'
mutants low levels of ci protein are expressed through the beginning of germ band extension but then decay. T h e expression of the en protein appears normal, but wg protein expression decays during stage12. In n'" mutants, at least some of the structures de-
In n.l14)!7 mutant5 wg protein expression returns during
238 D. C. Slusarski, C. Kelsey Motzny and R. Holmgren
rived from the en expressing cells are still generated. On
the ventral surface the first row of denticles is generally retained, and on the dorsal surface the isolated row of triangular hairs is present. The ventral naked cuticle that would be generated by the anterior en expressing cells is eliminated along with the rest of the ventral
naked cuticle. The phenotype of ci" mutants is very
similar to that of wg" mutants that have been shifted to
the nonpermissive temperature during stages 11 and/
or 12 (BAKER 1988;
BE~sowc
and MARTINEZ-ARIAS1991), suggesting that the ci" mutant phenotype can
be explained by the elimination of wg function. The
actual situation may be more complicated. SAMPEDRO
et 01. (1993) have shown that heat-shock expressed wg
is not able to cause naked cuticle formation in a cz"
mutant background. This result suggests that ci is epi-
static to wg in naked cuticle formation. The protein
product of the ci" mutation is actually a fusion protein.
The fusion protein may be missing up to 16 amino acids
from the N-terminus of ci and contains 2 2 0
x
103 M,.from a different protein. The ci" fusion-protein should
contain the entire zinc finger domain and the re-
maining C-terminal sequences. Clearly, the protein
product of the ci" allele is in part functional, and it is
possible that the fusion protein may have some novel gain of function properties.
The complementation observed between and
ci1(4"7 class mutations is probably due to their different effects on ci gene expression. In the ~ i ' " ' ' ~ class muta-
tions a promoter element required for early ci expres-
sion is disrupted, and the inversion responsible for the
ci" mutation fortuitously causes the expression of a functional ci fusion protein early in embryogenesis. In
trans-heterozygotes between ci" and the
~i"~"'
class mu-tants, a near wild-type pattern of expression is gener- ated, and embryonic development proceeds relatively normally.
Deletion analysis of the promoter defines a region
required for early ci expression: A series of deletions
were generated that eliminate the early embryonic ex- pression of ci protein. All such deletions affected se-
quences distal to the d'"" insertion site, suggesting that
an element required for early expression is located in this region. The position of this element is adjacent to
the gypsy insertion in ci' and distal to the putative I
element insertion in
ci'".
With gypsy insertions in theyellow locus it has been shown that regulatory elements between a gypsy element and the promoter are func- tional, whereas elements beyond a gypsy element inser-
tion are inactivated (GEYER and GORGES 1992). In a
modi& of mdg4 (mod) mutant background, gypsy can
inhibit all the elements that activate yellow expression
independent of their position (GEORGIEV and GERASI-
MOVA 1989). This is not the case with the gypsy insertion
in
ci.
mod; ci' animals have a phenotype that appearsto be identical to that of ci alone (data not shown) ;
therefore, the promoter elements required for embry-
onic ci expression do not appear to respond to su(Hw)/
mod regulation. The elimination of early ci expression
by the insertion in ci" can probably be explained by its
location between the promoter and the element re-
quired for early expression.
All of the c2"nr deletions complement the ci" muta-
tion. Thus, the element required for late embryonic
expression of the ci gene was not removed by any of
the deletions generated by excision of the ci'""' element.
It is tempting to suggest that the late embryonic ele- ment may be proximal to the insertion in citr because late embryonic expression is not altered in these mu- tants.
ci
wing
vein defects are due to inappropriate proteinexpression: In wild-type animals, ci mRNA and protein
expression are restricted to the anterior compartment of imaginal discs. The compartment boundary runs be-
tween wing veins three and four, so it was somewhat
surprising that a set of ci alleles cause defects in wing
veins four and five, which are derived from the posterior
compartment. This issue was resolved by finding that these mutations result in inappropriate expression of
the ci protein in the posterior compartment of the wing
imaginal disc. Ectopic expression in the posterior com- partment is also observed in the developing embryo.
In particular, ci" mutant embryos have high protein
expression levels throughout the embryo that do not appear to compromise embryonic development. This
suggests that the elimination of ci protein from the
posterior compartment is more critical for subsequent adult development.
In the ci' mutation, a gypsy element is responsible
for the ectopic expression in the posterior compart-
ment. The gypsy element contains binding sites for the
su(Hw) protein (SPANA et al. 1988). It appears that bind-
ing of the su(Hw) protein to the gypsy element medi- ates the dramatic increase in the level of ci mRNA be-
cause this increase is absent in a su(Hw) mutant
background.
Position effects at the ci locus: The ci locus is particu-
larly sensitive to chromosomal position effects. Many
translocations involving the fourth chromosome give
rise to ci mutant phenotypes (both recessive and domi-
nant) that have disruptions in wing veins four and five
(STERN and KODANI 1955). As we have shown, this type
of phenotype is due to inappropriate expression of the ci protein in the posterior compartment. Although posi- tion effects could cause inappropriate expression in sev-
eral different manners, two seem particularly attractive.
The translocation of sequences onto the fourth chro-
mosome might disrupt the normal en mediated repres-
sion of ci expression in the posterior compartment, or
the position effect could change chromosomal struc-
Regulation of ci Expression 239
promoter and nearby enhancer elements which are not subject to en regulation.
n' function is regulated posttranscriptionally The ci
mRNA is uniformly distributed in the anterior compart- ment as is Pgalactosidase expression from an enhancer
trap in the ci locus (EATON and KORNBERG 1990). The
pattern of ci protein expression in imaginal discs is
distinct from that of the mRNA. The ci protein levels, as visualized by antibody staining, are higher in regions adjacent to the compartment boundary. This is true for both the monoclonal antibody 2A1 and a rat polyclonal serum (data not shown). Therefore, it is unlikely that this distinction is due to the masking of a specific epi- tope. These results suggest that the rate of ci protein synthesis and/or turnover are regulated.
The product of the fu gene appears to play a role in
the posttranscriptional regulation of
ci.
In wing discselimination of the fu kinase elevates ci protein levels
throughout the anterior compartment. This is particu-
larly striking away from the compartment boundary
where the quantity of ci protein is low. The elevation
of ci protein levels is also seen in the cicx/+ wing discs.
Interestingly, alterations away from the compartment boundary in these mutants may not be relevant to the
wing vein defect. Mosaic analysis with fu has shown that
i t is only the region just anterior to the compartment
boundary that requires fu activity (FAUSTO-STERLING
1978).
The position of the ci gene in the segmental pat-
terning pathway: In the patterning of embryonic seg-
ments both ci and fu are epistatic to patched (ptc)
(FORBES et al. 1993) and ptc is epistatic to hh (INGHAM
et al. 1991). If a simple linear pathway exists from the
hh signal through the generation of naked cuticle, the
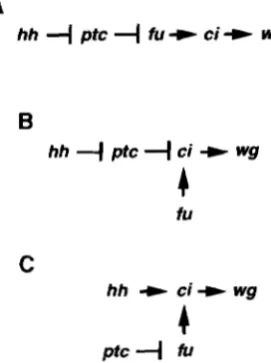
gene order would be hh, ptc, fu, ci, wg (Figure 9A)
(FORBES et al. 1993; PERRIMON 1994). Imaginal discs
develop at the compartmental boundaries of the em-
bryo and the patterns of hh, ptc, fu and ci expression in
imaginal discs closely reflect those established by stage
1 1 of embryonic development. hh is expressed in the
posterior compartment (LEE et al. 1992; MOHLER and
VANI 1992; TABATA et al. 1992), ptc (HOOPER and SCOTT
1989; NAKANO et al. 1989) and ci (EATON and KORNBERG
1990; ORENIC et al. 1990) are restricted to the anterior
compartment and fu (PREAT et al. 1990) is expressed in
all cells. The hh protein is secreted from cells in the posterior compartment and is thought to initiate a sig- nal transduction cascade in the cells adjacent to the compartment boundary. Therefore, it is of interest to
consider whether the linear pathway proposed above
can explain the pattern of ci protein expression in fu
mutant imaginal discs, If fu kinase activity is negatively regulated by ptc and ptc repression is relieved in re- sponse to the hh signal, then fu activity should be re- pressed in cells that are beyond the range of the hh
signal. In imaginal discs the fu kinase has a demon-
B
hh
4
ptc4
ci -t wg4
iU
C
hh
+
ci+ wgPtC
"I
fuFIGURE 9.-Models for the regulation of wgexpression and
naked cuticle formation within segments. (A) A linear model based on the epistasis relationships of hh, ptc, fu and ci. Cells receiving the hh signal will repress ptc protein function and activate the expression of wg. Cells at a distance from the hh signal will have active ptc protein, inactive fu kinase and no wgexpression. (B) A branched model in which the activity of the fu kinase is not regulated by the hh signal. In this model the activity of the ptc protein is regulated by hh. (C) An
alternative branched model in which the ptc protein is not regdated by the hh signal and instead ptc regulates the activ- ity of the fu kinase.
strated effect on ci protein levels far away from any potential hh signal, suggesting that branched or parallel pathways may be present.
A linear pathway is also inconsistent with the results
of SAMPEDRO et ai. (1993). As described above, the ex-
pression of wg from a heat-shock promoter is able to
cause naked cuticle formation in an hh mutant back-
ground but not in a c i D mutant background. Therefore,
there is a distinction between the roles of ci and hh.
This suggests that ci functions downstream of wg in
naked cuticle formation or that more complicated in-
teractions between wgand ci exist. Figures 9, B and C,
presents two branched models that are consistent with the observations. Additional models are possible, and a combination of genetic and biochemical experiments will be necessary to determine which best reflects the actual signal transduction pathway.
We thank DAVID WRIGHT for many stimulating discussions and for
his invaluable help in working out the relationship between ci" and
ci". We thank VICTOR CORCES, JOAN HOOPER, YU ZHANG, DARLENE
BUENZOW,JENNIFER KENNEDY, ANNA MARTI SUBIRANA and AMY BElso-
VK for critically reading the manuscript. R.H. thanks ANGUS LAMOND
and a Fogarty fellowship for giving him a desk, a bench and no telephone for his sabbatical at the EMBL in Heidelberg. This work was supported by National Science Foundation grant DCB 89-01394 and National Institutes of Health grant NS28472 to R.H.
LITERATURE CITED
240 D. C. Slusarski, C. Kelsey Motzny and R. Holmgren
BAKER, N., 1988 Localization of transcripts from the wingless gene in whole Drosophila embryos. Development 103: 289-298. BE~SOVE~, A., and A. MARTINEZ-ARIAS, 1991 Roles of wingless in pat-
terning the larval epidermis of Drosophila. Development 113: 471-485.
BEISO~C, A., and E. WIESCHAUS, 1993 Segment polarity gene inter-
actions modulate epidermal patterning in Drosophila embryos. Development 119: 501-517.
BERRY, A. J., J. W. AJIOKA and M. KREITMAN, 1991 Lack of polymor-
phism on the Drosophila fourth chromosome resulting from
selection. Genetics 129: 1111-1117.
CAMPOS-ORTEGA, J. A., and V. HARTENSTEIN, 1985 TheEmbryonic De-
velopmrnt oflkosophila melanoguster. Springer-Verlag, New York.
C M , I,.-N. and W. GEHRING, 1971 Determination of blastoderm cells in Drosophila melanogaster, Proc. Natl. Acad. Sci. USA 68:
CORCES, V. G., and P. K. GEYER, 1991 Interactions of retrotranspo- sons with the host genome: the case of the gypsy element of Drosophila. Trends Genet. 7: 86-90.
EATON, S., and T. KORNBERG, 1990 Repression of n'D in posterior
compartments of Drosophila by engrailed. Genes Dev. 4 1068-
1077.
FAUSTO-SPERLING, A,, 1978 Pattern formation in the wing veins of the /used mutant (Drosophila melanoguster). Dev. Biol. 63: 358- 369.
FINNEGAN, D. J., 1989 The I factor and I-R hybrid dysgenesis in Drosophila melanogaster, pp. 503-517 in Mobile DNA, edited by D. E. BERG and M. M. HOW. American Society for Microbiology, Washington, DC.
FORBES, A. J., Y. NAKANO, A. M. TAYLOR and P. W. INGHAM, 1993 Genetic analysis of hedgehogsignalling in the Drosophila embryo. Dev. Suppl. 119: 115-124.
GEORGIEV, P. G., and T. I. GERASIMOVA, 1989 Novel genes influenc- ing the expression of the yellow locus and mdg4 (gypsy) in Dro- sophila melanogaster. Mol. Gen. Genet. 220: 121-126.
GEW,R, P. K., and V. G. CORCES, 1992 DNA position-specific repres- sion of transcription by a Drosophila zinc finger protein. Genes Dev. 6: 1865-1873.
GONZALEZ, F., L. SWALES, A. DEJSOVEC, H. SUER and A. MARTINEZ- ARIAS, 1991 Secretion and movement of wingless protein in the epidermis of the Drosophila embryo. Mech. Dev. 35: 43-54. HAMA, C., Z. A L I and T. KORNBERC;, 1990 Region-specific recombina-
tion and expression are directed by portions of the Drosophila
engraikd promoter. Genes Dev. 4 1079-1093.
HEEMSKERK, J., and S. DINARDO, 1994 Drosophila hedgehog acts as a morphogen in cellular patterning. Cell 7 6 449-460.
HEEMSKKRK, J., S. DINARDO, R. KOSTRIKEN and P. H. O'FARRELI., 1991 Multiple modes of engrailedregulation in the progression towards cell fate determination. Nature 352: 404-410.
HIDALGO, A,, and P. INGHAM, 1990 Cell patterning in the Drosophila segment: spatial regulation of the segment polarity gene patched. Development 110: 291 -301.
HOCHMAN, B., 1973 Analysis of a whole chromosome in Drosophila.
Cold Spring Harbor Symp. Quant. Biol. 38: 581-589.
HOOPER, J. E., and M. P. SCOTT, 1989 The Drosophila patched gene encodes a putative membrane protein required for segmental patterning. Cell 59: 751-765.
HOOPER, J. E., and M. P. S~OTT, 199'2 The molecular genetic basis of positional information in insect segments. Results Probl. Cell Differ. 18: 1-48.
INGHAM, P. W., 1991 Segment polarity genes and cell patterning within the Drosophila body segment. Curr. Opin. Genet. Dev.
INGHAM, P. W., A. M. TAYLOR and Y. NAKANO, 1991 Role of the Drosophila patched gene in positional signalling. Nature 353: 184-187.
MNZI.ER, K. W., S. H. BIGNER, D. D. BIGNER, K. M. TRENT, M. L.
LAW et al., 1987 Identification of an amplified, highly expressed gene in a human glioma. Science 2 3 6 70-73.
LEE, J. J., D. P. VONKF.SSI.ER, S. PARKS and P. A. BEACHY, 1992 Secre- 2217-2221.
1: 261-267.
tion and localized transcript suggest a role in positional signaling for products of the segmentation gene hedgehog. Cell 71: 33-50.
MARTINEZ-ARIAS, A,, and P. A. LAWRENCE, 1985 Parasegments and
compartments in the Drosophila embryo. Nature 313: 639-642. MCKENZIE, S. L., S. HENIKOFF and M. MESELSON, 1975 Localization of RNA from heat-induced polysomes at puff sites in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 72: 1117-1121.
MOHLER, J., and K VANI, 1992 Molecular organization and embry-
onic expression of the hedgehoggene involved in cellcell commu- nication in segmental patterning of Drosophila. Development
115: 2635-2645.
N W O , Y., I. GUERRERO, A. HILDACO, A. TAYLOR, J. WHITTLE et
al., 1989 A protein with several possible membrane-spanning
domains encoded by the Drosophila segment polarity gene
patched. Nature 341: 508-513.
NUSSI.EIN-VOI.HARD, C., and E. WIESCHAUS, 1980 Mutations affect- ing segment number and polarity in Drosophila. Nature 287: 795-801.
O'CONNELI., P., and M. ROSBASH, 1984 Sequence, structure, and codon preference of the Drosophila ribosomal protein 49 gene. Nucleic Acids Res. 12: 5495-5513.
ORENIC, T., J. CHIDSEY and R. HOLMGREN, 1987 Cell and cubitus interruptus Dominant two segment polarity genes on the fourth chromosome. Dev. Biol. 1 2 4 50-56.
ORENIC, T., D. C. SI.USARSKI, K L. KROLL and R. HOLMGREN, 1990 Cloning and characterization of the segment polarity gene cubi- tus i n t m p t u s Dominant of Drosophila. Genes Dev. 4 1053- 1067. PATEI., N. H., B. SCHAEER, C. S. GOODMAN and R. HOLMGREN, 1989 The role of segment polarity genes during Drosophila neurogen- esis. Genes Dev. 3: 890-904.
PERRIMON, N., 1994 The genetic basis of patterned baldness in Dro- sophila. Cell 7 6 781-784.
PREAT, T., P. TI-IEROND, C. LAMOUR-ISNARD, B. LIMBOUG-BOUC:HON,
H. TRICOIRE et al., 1990 A putative serine/threonine protein kinase encoded by the segment-polarityyfusedgene of Drosophila. Nature 347: 87-89.
ROBERTSON, H. M., C. R. PRESTON, R. W. PHII.I.IS, D. M. JOHNSON-
SC€II.ITZ, W. K. BENZ et al., 1988 A stable genomic source of P element transposase in Drosophila melanogaster. Genetics 118: 461-470.
RUPPERT, J. M., B. VOGEISTEIN, K. ARHEDEN and K. W. KINZLER, 1990 GLI? encodes a 190-kilodalton protein with multiple regions of GLI similarity. Mol. Cell. Biol. 10: 5408-5415.
SAMPEDRO, J., P. JOHNSTON and P. A. LAWRENCE, 1993 A role for wingless in the segmental gradient of Drosophila? Development
117: 677-687.
SPANA, C., D. A. HARRISON and V. G. CORCES, 1988 The Drosophila melanogaster suppressor of Hairy-wing protein hinds to specific sequences of the gypsy retrotransposon. Genes Dev. 2: 1414-
1423.
STERN, C., and M. KOUANI, 1955 Studies on the position effect at the rubitus interruptus locus of Drosophila melanogaster. Genetics 40: 343-373.
TABATA, T., S. EATON and T. B. KORNBERG, 1992 The Drosophila hedgehog gene is expressed specifically in posterior compartment cells and is a target of engruiled regulation. Genes Dev. 6: 2635- 2645.
TAUTZ, D., and C. PFEIFLE, 1989 A non-radioactive in situ hybridiza- tion method for the localization of specific RNAs in Drosophila embryos reveals translational control of the segmentation gene
hunchback. Chromosoma 98: 81-85.
VAN n m HEUVFI., M., R. Nusse, P. JOHNSTON and P. A. LAW,NC:E,
1989 Distribution of the wingless gene product in Drosophila embryos: a protein involved in cell-cell communication. Cell 59: 739-749.
VAN DER MEER, J. M., 1977 Optical clean and permanent whole mount preparations for phase contrast microscopy of cuticular structures of insect larvae. Drosophila Inform. Serv. 52: 160.
ZARKOUTER, D., and J. HODGKIN, 1992 Molecular analysis of the C .
eleguns sex-determining gene tru-1: a gene encoding two zinc finger proteins. Cell 70: 237-249.