THE EFFECT OF RADIATION ON SMALL COMPETING POPULATIONS OF DROSOPHILA MELANOGASTER.
IV. CHANGES IN LONGEVITY.
K. F. DYER*
Medical Research Council, Radiobiology Unit, Harwell, Didcot, Berkshire, England.
and
MARION J. LAMB
Zoology Department, Birkbeck College, Malet Sireet, London, W.C.I., England.
Received January 27, 1971
PREVIOUS investigations of the cumulative effect of irradiation on the fitness of Drosophila populations have concentrated mainly on changes in fitness during the pre-adult stages. In this paper we describe the results of a study of changes in what may be considered as another aspect of fitness, adult lifespan. The irradiated populations which provided the material for this study were those in which DYER investigated the accumulation of genetic damage (DYER 1966, 1969a,b) and changes in competitive ability (DYER 1 9 6 9 ~ ) . W e can therefore attempt to relate changes in lifespan with the genetic changes, and also to com- pare changes in competitive ability, a component of fitness for which in our ex- periment there was strong positive selection, with changes in longevity for which there was no direct selection.
MATERIALS A N D METHODS
The flies used for this investigation were from the populations described in detail in the first paper in this series (DYER 1969a) and previously (DYER 1966). Two separate inbred lines, each of which was segregating for the two closely linked autosomal recessive markers light and straw, were maintained by close brother-sister mating for 146 generations. The foundation populations for the experiments consisted of homozygous light and homozygous straw flies obtained from a cross of these two inbred lines. Twenty-five males and 25 virgin females of both light and straw phenotypes were put into a population cage containing 200 ml of a standard maize meal-agar- sugar food medium seeded with live yeast. After 5 days these parents were removed. The off- spring from the cage were collected and counted, and 25 males and 25 virgin females of both light and straw phenotypes were put into a fresh cage which was continued as previously. This procedure was repeated for 25 generations. Since the heterozygotes were eliminated at each gen- eration, the light and straw lines remained genetically distinct throughout the experiment.
Two populations were maintained: in one, the Control line (C), neither light nor straw flies received any irradiation; in the other, the Irradiated line (R), the 25 male and 25 female straw flies which formed half of the parents at each generation were irradiated with 1500 R X rays (250 kVp, HVL 1.2 mm Cu) at. 546 R/min immediately before being put into the cage.
From the light, straw and heterozygotes which emerged at each generation up to 50 virgin females of each phenotype were used for longevity measurements. It must be stressed that none
* Present address: D e p a r h i e n t of Genetics, hlonash University, Clayton, Victoria 3168, Australia.
560 K. F. DYER A N D M. J. LAMB
of these flies had received any direct radiation so any effect on the lifespan of the straw and heterozygous flies from the R cage was due to the ancestral radiation. Females were used rather than males since experience has shown that the lifespans of females are less influenced by en- vironmental fluctuations than the lifespans of males. It was necessary to use virgin females since mating and the changes in egg-laying that mating produces are known to affect longevity
(MAYNARD SMITH 1958). Longevity comparisons were made between flies emerging on the same days. Wherever possible, flies emerging on one particular day were used, but where this was impossible, flies emerging on three consecutive days were used and the second day was taken as the day of emergence for all of them. The flies were kept at 25"C, 5 flies per 7.5 x 2.5 cm tube containing the standard food medium seeded with live yeast. They were transferred to fresh medium every two days. Deaths were recorded at the time of transfer and were assumed to have occurred mid-way i n the interval between transfers.
RESULTS
Most of the results of the studies of the accumulation of genetic damage and changes in competitive ability in the irradiated populations have been reported in the preceding papers in this series. However, since these results are important in the interpretation of the longevity data presented in this paper, they will be briefly summarised here before considering the results on longevity.
(i) The accumulation of genetic damage.-The second chromosome recessive lethal mutation rate in the irradiated flies was 6% per generation (DYER 1966). The mean rate of accumulation in the early generations was 4.4% but after gen- eration 14 the net rate of increase was very low; an equilibrium of between 60
and 70% lethal chromosomes was reached in these later generations (DYER
1969a). The frequency of second chromosome recessive lethal mutations in the heterozygotes was between 25 and 30% which was significantly less than the mean lethal frequency of the light and straw populations from which they were derived (DYER 1969a, Table 4 ) .
DYER (1969b) estimated that, assuming that the third chromosome was similar
to the second, after generation 14 more than 95% of the straw flies from the ir- radiated population carried at least one autosomal recessive lethal and that be- tween 25 and 30% carried 3,4,5 or more lethals. In addition there was evidence of increasing sterility and an increase in the frequency of sex-linked lethals. The levels of genetic damage in the irradiated populations were clearly very high.
ANCESTRAL RADIATION A N D LONGEVITY 561
P
I I I I f I I f f I I I I 1 ' 1 1 ' 1 1 1 1 1 1 1 1
0 5 10 15 20 25
G E NE R AT1 ON
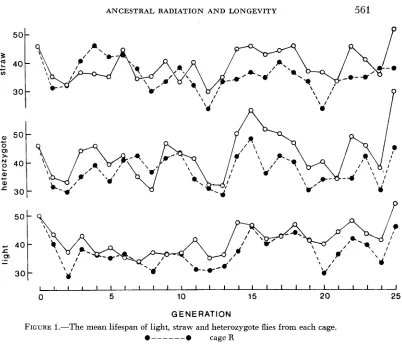
FIGURE 1.-The mean lifespan of light, straw and heterozygote flies from each cage.
.---e cage R
0- 0 cageC
dominant mutations. During the course of the experiment the competitive ability of the irradiated flies improved overall by between 12 and 15
% despite the severe
radiation regime (DYER 1969c), this increase in competitive ability being most clearly demonstrated after the suspension of irradiation. Generations 2-7 showed a marked improvement although there was later a fall from this maximum level (DYER 1966, Fig. 6 ) . The mean disadvantage of flies heterozygous for the irradi- ated chromosomes was, however, 40% and they showed no sign of an overall in- crease in competitive ability during the course of the experiment.(iii) Longeuity-Both the mean lifespans and the variances fluctuated con- siderably from generation to generation. This was not a n unexpected finding since the lifespan of Drosophila is extremely sensitive to minor environmental changes. The fluctuations in lifespan and relatively small numbers of flies in each sample make a rigorous quantitative analysis of the data difficult, but since for each generation the lifespan of the irradiated straw flies can be compared with that of the light flies from the same cage and that of the straw flies €rom the C
562 K. F. DYER AND M. J. LAMB
(a) The lifespan of flies from the C and R cages: For each genotype the lifespan of the flies in the R cage tended to be less than that of those in the C cage. The
variance also showed consistent differences between the C and R cages. For the light genotype both the variance and coefficient of variation were less in the C
cage than in the R and the same tendency was shown with the heterozygotes al- though it was less well marked. With the straw flies, however, the situation was reversed: the variance and coefficient of variation tended to be less in the R cage than in the C cage.
(b) T h e lifespan of the light pies: The differences between the means and variances for the light flies from the two cages was surprising since the flies which formed the two parental generations were identical; in neither cage did the light flies receive any radiation and there was no opportunity for gene flow from the
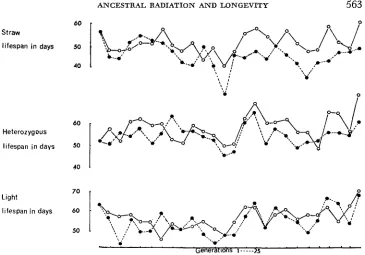
straw to the light populations. However, examination of the distribution of time of death of flies from the two cages suggests a possible explanation. For all geno- types there is a tendency for the distribution of time of death to be skewed to the left and the differences between the means and variances of the light flies from the two cages are due largely to differences in the proportions of flies dying early. Figures 2 and 3 show the means of the first and last quarters of the flies dying. For the light flies there is no difference between the means of the last quarters but for the first quarters the mean of the R light flies is consistently less than that
Generations 1 - - - 2 5
FIGURE 2.-The mean lifespan of the first quarter of the light, straw and heterozygote flies to die.
.---e cage R
0
ANCESTRAL RADIATION A N D LONGEVITY 563
Straw lifespan in days
Heterozygous
lifespan in days
Light
lifespan in days
60
50
40
4 0 I
FIGURE 3.-The mean lifespan of the last quarter of the light, straw and heterozygote flies to die.
.---e cage R
0- 0 cageC
for the C light. W e suggest that this is correlated with the difference between the cages in the numbers of light flies emerging: because irradiation reduced the number of straw flies emerging from the R sages, the number of light flies was greater than from the C cages. If this increase in the number of light flies emerging means that more of the ‘less fit’ flies reach the imaginal stage, and if
these less fit flies have a shorter lifespan than the others, then the reduction of the mean and increase in the variance of the lifespan of light flies from the
R
cage can be explained. The essential point is that flies which will survive from the egg to the adult stage only if the competitive conditions are reduced are presumed also to have inferior ability to survive as adults.5
64 K. F. DYER A N D M. J. LAMB +10 0, U C 0 O m !! + 5
Z W
C O O
c
m .5
Q U) 0 ) - 5 .C .- -I
1
-10 +10 Q) CU + 5
E
I n
..- C A 0
0 :
5 =
/*-•
P
/"- 5
Q .- v) Q) -I .C .- -10 d
G E N E RAT IONS 1 . *
. .
25FIGURE 4.-Three generation moving average of the difference between the lifespans of light and straw flies and the lifespans of the light and heterozygote flies from each cage.
0
-
- - - 0 0 - - - - - -0.----@
0- 0
straw minus light from cage R heterozygotes minus light from cage R straw minus light from cage C heterozygotes minus light from cage C
is less than that of the light, significantly so at generations 12, 15, 18. 19 and 25. The variance of the straw flies from the R cage is less than that for light from the same cage or for straw flies from the C cage at most generations.
Figures 2 and 3 show that the situation regarding the mean lifespan of the first and last quarters of the straw flies to die is very different from that seen with the light flies where the difference between the two cages was due to differences in the mean lifespans of the first quarter of the flies to die. For the straw flies be- tween generations 2 and 8 the mean of the first quarter is greater for flies from the R cage than for those from the C cage, but there is little difference between the means of the last quarters of the flies to die. These are the generations when the longevity of the irradiated straw flies is superior to that of the unirradiated light flies from the same cage. They are also the generations in which the ir- radiated straw flies showed an increase in competitive ability in the pre-adult stages (DYER 1966, 1 9 6 9 ~ ) . In this case therefore we conclude that the enhanced competitive ability and increase in lifespan can be attributed to changes in flies which for genetic reasons normally have poor pre-adult competitive ability and a high probability of early adult death. For this specific category of flies the effect
ANCESTRAL RADIATION A N D LONGEVITY 565
L
.-.
a .
I \ I .
, \
50
45
C
P
YI
,
Generations I-. .. .25
FIGURE 5.-Comparison of the weighted mean lifespan of the light and straw flies with that of the heterozygotes from the same cage.
0- 0
0
- -
-
- -
-
-
-0 0
W --- W
mean lifespan of the light and straw flies from cage R
mean lifespan of the heterozygotes from cage R
mean lifespan of the light and straw flies from cage C
mean lifespan of the heterozygotes from cage C
In later generations, with increasing genetic damage, DYER ( 1 9 6 9 ~ ) found a small decrease in competitive ability and, as Figure 4 shows, there was a marked decline in the relative lifespan of the irradiated straw flies. The means of both the last and first quarters of the irradiated straw flies to die tend to be less than those of the unirradiated straw. It seems therefore that there has been a genuine relative decrease in the overall mean lifespan of the irradiated flies in later gen- erations. When the lifespan of the irradiated straw flies is compared with that of the competing light population, o r with that of the unirradiated straw flies, or with that of flies from earlier generations, it appears to be reduced by a n average of 10-15%, and at its maximum the reduction reaches 25%. Since we have already seen that there is a reduction of at least 10% in the lifespan of each of the genotypes from the R cage compared with cage C, we may suppose that the net effect of accumulated ancestral radiation on lifespan is a reduction of no1 more than about
5 % .
(d) The lifespan of the heterozygote flies: The heterozygote flies are a discon- tinuous population. Since they take 50% of their chromosomes from each of the
light and straw populations, their lifespan can be compared with the average life- span of these two populations or, alternatively, they can be compared with the completely unirradiated light flies.
566 K . F. DYER A N D M. J. L A M B
for the heterozygotes from the C cage to have a longer lifespan from generation 14 onwards. Since the light and straw flies became more inbred with each gen- eration. this relative increase in lifespan of the heterozygotes may be due to hy- brid vigour (CLARKE and MAYNARD SMITH 1955) but none of the differences is significant.
I n the
R
cage the heterozygotes show superiority over the mean for light and straw between generations 6 and 12 although the individual differences are them- selves not significant. However, as Figure 4 shows, for these generations the heterozygotes also have a greater lifespan than the unirradiated light flies from the same cage and two of these differences are significant so the trend is thought to be meaningful. Figures 2 and 3 show that, apart from these 6 generations, there is normally a slightly greater difference between the mean of the first quarter of theR
and C heterozygotes dying than between the last quarters although these differences are not at all significant. We therefore believe that the explanation of the reduction in the relative lifespan of the heterozygotes from the R cage in later generations and the increase in the coefficient of variability which accom- panies it is similar to that which we have suggested in the case of the light popu- lations where this difference was more pronounced, namely that rather more of the poorer genotypes are managing to emerge but that they die early and there- fore reduce the mean lifespan and increase the overall variance.Between generations 6 and 12 the number of heterozygotes emerging from the
R
cage underwent a marked decline. If it was the poorer genotypes which were failing to emerge, then the relative increase in the overall mean lifespan can be explained. The fact that the variance of lifespan did not decrease is not, on the face of it, compatible with such a suggestion. DYER ( 1 9 6 9 ~ ) however, has pre- sented evidence that many of the radiation induced mutations have rather dif- ferent effects on the straw and heterozygote populations and that in general only lethal genes very favourable in the straw line and therefore fairly frequent are not at a disadvantage in the heterozygotes. There is in fact quite a difference in the array of lethal genes recovered from the straw population compared to that recovered from the heterozygotes. and this is evidence of the different gene pools of the two populations. DYER (1968) has also presented evidence to show that the disruption caused by forced heterozygosis through interpopulation hybridisa- tion has drastic effects on competitive ability. Severe biological upsets of this type make predictions of possible lifespan changes very difficult and the anomalies present in our data for lifespan of heterozygotes could only be resolved by a more detailed analysis and different experimental design.DISCUSSION
The discussion will be restricted to the results of the single group of experi- ments which have been described. W e did, in fact, repeat these experiments in essential details up to 12 generations of radiation and obtained similar results.
ANCESTRAL RADIATION A N D LONGEVITY 567 and second generation offspring of mice irradiated with neutron or 6oCo y-rays; LINDOP and ROTBLAT (1963) reported no life-shortening effects among the prog- eny of male mice irradiated for three generations with 350 R X rays; FROL~N
(1965) found no effect on the offspring of male mice which had received 500 R of X rays. The cumulative effects of six generations of ancestral irradiation with 6oCo y-rays on five strains of mice led GOWEN and STADLER (1964) to the con- clusion that ‘the ancestral irradiation over the six generations had little effect on the lifespan of the mice’ and SCHLAGER, RODERICK and STORER (1966) reported that ‘no consistent effect of ancestral spermatogonial X irradiation on longevity was found’ when mice were exposed to 50 and 100
R
per generation o r 900R
for six generations. SPALDING, BROOKS and MCWILLIAMS (1966) concluded that 25 generations of ancestral X irradiation had no effect on longevity although other components of fitness were affected adversely. An exception to these broadly negative findings are the results of MORIWAKI and TOBARI (1960) that the off- spring of male and female Drosophila melanogaster irradiated with up to 3000 R X rays had a reduced lifespan, the effect being more marked among the male offspring than the female offspring.The results from the present experiments show that the lifespan of female Drosophila, kept under the conditions that we have described, is reduced by no more than 5% compared with an unirradiated population after twelve genera- tions of 1500 R X rays per generation, and that there is no further significant re- duction during a further 13 generations of irradiation. Since prior to generation 12 there had been an apparent increase in lifespan of irradiated flies, the actual progressive reduction in lifespan from the maximum value reached amounted to 10-15%. Our results suggest that three distinct processes are each in part re- sponsible for these changes in mean lifespan during the course of the experiment. (i) The induction of dominant lethals and the increasing occurrence of homo- zygous recessive lethals reduced the number of individuals competing f o r a lim- ited amount of food during the larval stages. As a consequence of this reduction in competition, more of the relatively less fit flies reached the adult stage and these flies had a relatively short lifespan. This effect of ancestral irradiation was entirely indirect; in our data it was in fact shown most clearly among the un- irradiated light flies which were competing with the irradiated straw population in each generation. These unirradiated light flies from cage
R
had a lower mean lifespan and the coefficient of variation was greater than that of the light flies from cage C , but there was no change in the maximum lifespan. The effects of larval density on longevity have been inveztigated by MILLER and THOMAS(1958) and by LINTS and LINTS (1969a). In both of these studies adult longevity increased when larval density was increased. These authors have also shown that the size of the adult diminishes with increasing larval density and LINTS and
568 K. F. DYER A N D M. J. LAMB
(ii) Between generations 2 and 7 there was an increase in the mean lifespan of the flies in the irradiated population due largely to a reduction in the propor- tion of flies dying early. This increase in mean lifespan was found in the same generations as DYER (1966) found an increase in the competitive ability of the irradiated population. DYER suggested that this increase in competitive ability could be explained in terms of the optimum heterozygosity hypothesis developed by
WALLACE
(1958) and MUKAI, CHIGUSA and YOSHIKAWA (1965). The increase in longevity found in the early generations of the irradiated straw population is also compatible with this hypothesis.(iii) In later generations the lifespan of the irradiated population decreased. This decrease appeared to be a general decrease in the lifespan of all the flies since the means of both the last and first quarters of the flies to die were less than those of the control populations. In these generations DYER found that the competitive ability of the flies was also tending to decrease.
Our main conclusion from these experiments is that the direct effects of an- cestral radiation on lifespan were small. In general, the effects on lifespan paral- leled the effects cm pre-adult competitive ability although there was no selection for increased longevity and very intense selection f o r increased competitive ability. The similarity of the effect of ancestral radiation on lifespan and com- petitive ability is. perhaps, not surprising since it must be assumed that many of the genes and gene combinations which are important during pre-adult stages are also important in adult life.
S U M M A R Y
Two populations, one of which was homozygous for the marker gene light and the other for the marker gene straw but which were otherwise genetically almost identical, were allowed to compete in the same population cages for 25 successive generations. 25 males and 25 virgin females of each phenotype were used as par- ents for each generation and at the beginning of each generation the straw par- ents were irradiated with 1500 R
X
rays. The longevities of virgin females from the light and straw populations and the heterozygotes between them were measured and compared. Comparisons were also made with the lifespans of fe- males from control cages in which neither population had received any irradia- tion. The lifespans of all three genotypes from the cages in which the strawA N C E S T R A L R A D I A T I O N A N D L O N G E V I T Y 569
small changes in longevity paralleled the changes found in pre-adult competitive ability.
L I T E R A T U R E C I T E D
CLARKE, J. M. and J. MAYNARD SMITH, 1955 The genetics and cytology of Drosophila subobscura XI. Hybrid vigour and longevity. J. Genet. 53: 172-180.
Fitness and competitive ability in irradiated populations of Drosophila melanogaster. Mutation Res. 3: 327-339. -, 1968 The effects of accumulated an- cestral irradiation on some populations of Drosophila. Heredity 23: 630-631 1969a The effect of radiation on small competing populations of Drosophila melanogaster. I. The accumulation of genetic damage. Genetics 61 : 227-244. -, 1969b The effect of radia- tion on small competing populations of Drosophila melanogaster. 11. The changing fre- quencies of autosomal recessive lethals. Genetics 61: 245-274 -, 1969c The effect of radiation on small competing populations of Drosophila meLanogaster. 111. Changes in competitive ability. Genetics 61 : 275-291.
The effect on the length of life in the offspring of X-irradiated male mice.
Lifespans of mice as affected by continuing irradiation from Cobalt-60 accumulated ancestrally and under direct irradiation. Genetics 50: 11 15-1 142.
Study of the offspring of irradiated mice. Summarised by E. C. AMOROSO. pp. 408429. In: Radiation Effects in Physics, Chemistry and Biology. Edited by M. EBERT and A. HOWARD. North Holland Publishing Co., Amsterdam.
Respiration in Drosophila. 111. Influence of pre-imaginal environment on respiration and ageing in Drosophila melanogaster hybrids. Exp. Gerontol. 4: 81-94. -, 196913 Influence o l pre-imaginal environment on fecundity and ageing in Drosophila melanogaster hybrids. I. Pre-imaginal population density. Exp. Gerontol. 4 :
The effects of temperature and of egg-laying on the longevity of Drosophila subobscura. J. Exptl. Biol. 35: 832-842.
The effects of larval crowding and body size on the longevity of adult Drosophila melanogaster. Ecology 39: 118-125.
The genetic effects of radiation on the longevity of progeny in Drosophila melanogaster. J. Radiation Res. 1 : 14-22.
The genetic structure of natural populations of Drosophila melanogaster. 111. Dominance effect of spontaneous mutant polygenes con- trolling viability in heterozygous genetic backgrounds. Genetics 52 : G3-501.
Shortening of life in the offspring of male mice exposed to neutron radiation from an atomic bomb. Proc. Natl. Acad. Sci. U S . 43: 324-329.
Longevity and body weights of mice with ancestral spermatogonial X irradiation. Mutation Res. 3 : 230-236.
Longevity of first and second generation offspring from male mice ex- posed to fission neutrons and gamma-rays. pp. 147-152. In: Effects of Ionizing Radiation on the Reproductiue System. Edited by W. D. CARLSON and F. X. GASSNER. Pergamon Press, London.
Reproductivity and lifespan of mouse
The average effect of radiation-induced mutations on viability in Drosophila DYER, K. F., 1966
-
,FROL~N, H., 1965
Mutation Res. 2: 287-292. GOWEN, J. W. and J. STADLER, 1964
LINDOP, P. J. and J. RGTBLAT, 1962
LINTS, F. A. and C. V. LINTS, 1969a
231-244.
MAYNARD SMITH, J., 1958
MILLER, R. S. and J. L. THOMAS, 1958
MORIWAKI. D. and I. TOBARI, 1960
MUKAI, T., S. CHIGUSA and I. YOSHIKAWA, 1965
RUSSELL. W. L., 1957
SCHLAGER, G., T. H. RODERICK and J. B. STORER, 1966
SPALDING. J. F., 1964
SPALDING, J. F., M. R. BROOKS and P. MCWILLIAMS, 1966
WALLACE. B., 1958
populations from 25 generations of irradiated sires. Genetics 54: 755-761.