HOMOLOGOUS CHROMOSOME PAIRING REMAINS A N
UNSOLVED PROBLEM: A TEST OF A POPULAR HYPOTHESIS
UTILIZING
MAIZE MEIOSIS
MARJORIE P. MAGUIRE
Zoology Department, University of Texas, Austin, Texas 78712
Manuscript received December 27, 1982 Revised copy accepted January 17,1983
ABSTRACT
A recently proposed and popular model for the mechanism of meiotic homologue pairing relies on prior association of nonhomologous chromosome arms of most similar length. According to this model, the diploid complement is organized into two genomic linear chains, each containing the various heter- ologues in the same sequence. At meiosis, then, appression of the two genomic chains could presumably readily accomplish homologue pairing. This model fails in its simplest form when observations of meiotic pairing of homologues in heterozygotes for arm length alterations are compared with computer-simulated predictions of the model. Contrary to predictions of the model, heterozygotes for arm length changes were found to exhibit only small frequencies of homo- logue-pairing failure, and this only for a single homologue pair in each case. It is difficult to conceive of a reasonable modification of this model that would be consistent with the observations.
HE question of how homologous chromosomes pair with each other at
T
meiosis remains a major unsolved problem of biology. This vital feat is one of several which must be accomplished in most diploid organisms in order to provide for normal chromosome segregation with the production of gametes which carry exactly one chromosome representative of each member of the genome. Even a minor failure, involving only a single chromosome pair, can result in inviable gamete production or abnormal offspring, such as Down’s syndrome.Mechanisms for homologue pairing which have been proposed fall into three general categories: (a) long-range specific force of attraction, (b) random meeting of homologues in motion, and (c) mediation of specialized, extrachromosomal structures. As pointed out long ago (FABERGE 1942), ordinary specific intermo- lecular forces of attraction are extremely weak at distances greater than contact, and the case for random motion of chromosomes relative to each other which might fortuitously produce contact of homologues at corresponding positions (as required) is conjectural (MAGUIRE 1974) or supported by very limited obser- vational evidence (PARVINEN and SODERSTROM 1976). Thus, cytologists have increasingly sought solution to the problem in terms of hypothesized interac- tions of specific chromosome parts, especially the telomeres or the centromeres, with specialized sites on the inner nuclear membrane (AVIVI and FELDMAN 1980;
174 M. P. MAGUIRE
COMINGS 1980). Telomeres and centromeres, in fact, often appear to be attached to the inner nuclear membrane, but precise juxtaposition of homologous telo- meres or centromeres before the synaptic stage is usually not impressive or is blatantly absent, and synaptic contact of homologues seems not to be initiated precisely at the telomeres, even though points of initial synapsis may be close to terminal (BURNHAM et al. 1972; MOENS 1969; ZICKLER 1977).
Interestingly, it has been reported that pairs of nonhomologous telomeres (or
approximately terminal centromeres) of the chromosomes within the haploid complement of Ornithogalum virens pollen tend strongly to be associated so as consistently to produce a specific linear chromosome arrangement (ASHLEY 1979). Since this specific linear order tends to persist (or be reestablished) in diploid cells, it was suggested that a relatively simple, homologous attachment
of the two ends of each of the two pairs of terminal members of the linear arrays in the two genomes to produce a circle would in turn yield a structure that could give side by side orientation of homologues upon appropriate appres- sion of the two genomic halves of the circle (ASHLEY 1979). The possibility of general applicability of this idea has gained considerable support from studies of the relative positions of identifiable chromosomes in barley and rye somatic cells (BENNETT 1982). Observations suggest that, although a variety of distribu- tions are found, statistically, homologues tend to be farther apart than random expectation predicts at diploid mitosis, and a specific kind of spatial distribution within each genome tends to prevail. In the kind of arrangement that seems favored nonhomologous chromosome arms of most similar length tend to be adjacently positioned to produce the same sequence within each genome, with the additional proviso that long arms will be adjacent to long arms and short arms adjacent to short arms. It seems reasonable to speculate that such order might follow from some kind of interaction of the spindle during nuclear division with the varying quantities of chromosome material present in their respective arms, from the tendency of chromosome positions at anaphase to persist into interphase and even prophase of the next division, and from association of telomeres to the inner nuclear membrane. Acknowledging that the problem of how chromosome arms recognize and attach to their correct partners (once this order is established) remains unknown, LEWIN (1981) was prompted to conclude that “If the description of chromosome position is correct, then the problem of homologous pairing is immediately solved.”
In view of the appeal of the idea that the stage could be so simply set for meiotic homologue pairing, it occurred to me that this hypothesis should be tested with other approaches. For example, if homologue pairing generally depends upon the telomere pairing of nonhomologous chromosome arms of most similar length, established, for example, at the end of mitotic anaphase (or
upset the pairing of a number of chromosomes. Although generalized meiotic disturbance with heterozygosity for a single rearrangement has not been re- ported (to my knowledge), it seemed possible that appropriate heterozygotes had not been duly scrutinized in this respect. Accordingly, the behavior of chromosomes at meiotic prophase in maize was studied in heterozygotes for pericentric (between arm) inversions characterized by original breakpoint po- sitions at differing distances from the centromere in the two arms (so as to produce changes in arm length). Maize may be uniquely suitable for such a study because a variety of known inversions is available, and meiotic cytology is probably unsurpassed.
MATERIALS AND METHODS
Inversions studied were In 50 and In 7-3717. In In 50 the ratio of long arm to short arm of chromosome 5 is changed from 1.1 to 2.9. In In 7-3727 the ratio of long arm to short arm of chromosome 7 is changed from 2.6 to 1.5. Also, in the case of the chromosome 5 inversion, the distal portion of the normal short arm (including the telomere) is now a part of the physically longer arm, and the distal portion of the normal long arm (including the telomere) is now a part of the physically shorter arm. Thus, the change in the genome which results from the chromosome 5 inversion is the more drastic of the two with respect to alteration of relative arm length, and it is very much more drastic in this respect if the telomeres carry a persistent memory, as to which arm is the longer, after chromosome rearrangement (as suggested by BENNETT (1982) for a barley rearrangement stock).
Plants heterozygous for pericentric inversion In 7-3727 or In 5a were grown in a growth chamber. Microsporocyte samples were collected at meiotic stage, fixed in ethanol acetic 3:l mixture and stored in a freezer until examined in acetocarmine squash preparations with high resolution (1.4 N.A.) light microscopy.
RESULTS AND DISCUSSION
In each case virtually all cells were classifiable at diakinesis, metaphase I,
anaphase I and anaphase I1 stages for meiotic chromosome behavior, and all were normal except as follows. In In 5a samples, nine bivalents and two univalents were found in 19 of 298 (6%) of cells at diakinesis, as well as in 19 of
352 (5%) of cells at metaphase I, and at anaphase I one or two laggards (a common form of behavior of maize univalents) were found in 23 of 317 (7%) of cells. In In 7-3737 samples nine bivalents and two univalents were found in 18
of 125 (8%) of diakinesis cells and in 26 of 268 (10%) of metaphase I cells, and one or two laggards were found in 7 of 91 (8%) of anaphase I cells. In both cases impressive numbers of cells at anaphase I1 (15 and 22% respectively) contained two laggards, apparently usually a result of premature separation of an other- wise normally behaving dyad during the second meiotic division (MAGUIRE
1982).
176 M. MAGUIRE
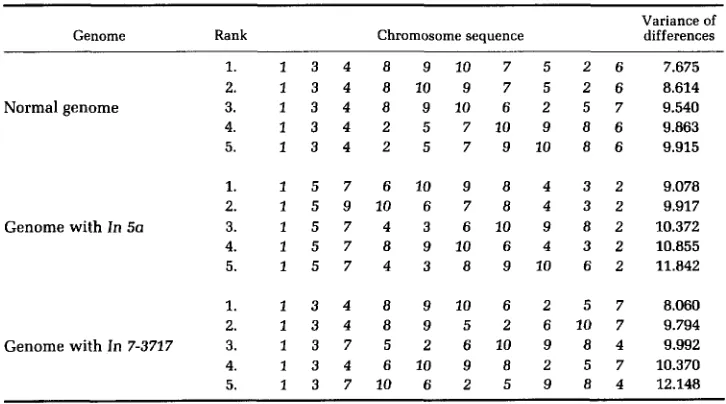
follows, Standard arm lengths of the ten chromosomes of the normal maize genome, expressed as percentage of the total length of chromosome 10 (NEUFFER, JONES and ZUBER 1968), and appropriately revised arm lengths for the two inverted chromosomes were utilized to generate with computer the five best linear chromosome sequences (with respect to each of two criteria) for the normal genome, the genome with In 5a and all other chromosomes normal, and the genome with In 7-3727 and all other chromosomes normal. One criterion was minimum sum of differences between adjacent chromosome arm lengths as in BENNETT’S model (Table 1); the other criterion was minimum variance of differences between adjacent chromosome arm lengths (Table 2). In both cases the additional proviso used by BENNETT was also imposed, that long arms could only be positioned adjacent to long arms and short arms adjacent to short arms. In the case of the chromosome 5 inversion it was assumed that physical length and not original telomere identity determined which arm was long, because to do otherwise would have imposed new, very large arm length difference on any linear sequence generated. (In the case of the chromosome 7 inversion, the original long arm and short arm telomeres are still located on the physically long and short arms.) All possible sequences, with the proviso listed before and with the understanding that the short arm of chromosome 2 must be terminally located (because of its great difference in length from all other short arms), were examined by the computer. It can be seen from Tables 1 and 2 that the best sequences for the genome containing the In 5a inversion are very different from the sequences for the normal genome, and that the best sequences for the genome containing the In 7-3727 inversion are also, for the most part, different from the sequences for the normal genome, although the third best sequence for variance of differences for the normal genome is the same as the best sequence for variance of differences in the In 7-3717 inversion genome. Thus, these computer-generated sequences suggest that, if homologous chromosome pairing depended upon the formation of a linear array of chromosomes, asso- ciated by their telomeres in each genome such that arms of most closely matching length tended to occupy adjacent positions, the pairing of much more than one pair of homologues should be consistently disrupted in the case of heterozygosity for In 5a, and probably also in the case of In 7-3717, since appression of the two genomic chains would fail to produce juxtaposition of homologues.
177
TABLE 1
Best sequences for sum of differences
Sum of dif-
Genome Rank Chromosome sequence ferences
~~ ~
1. 1 3 4 2 5 7 9 1 0 8 6 54.5 2. 1 3 4 8 1 0 9 7 5 2 6 58.7
Normal genome 3. 1 3 4 2 5 7 9 1 0 6 8 60.0
4. 1 3 4 2 5 7 8 6 1 0 9 61.0 5. 1 3 4 2 5 7 1 0 9 8 6 62.1
1 5 9 1 0 6 7 8 4 3 2 53.3 Tie
{::
1 5 9 10 6 8 7 4 3 2 53.3.
1 5 7 8 9 1 0 6 4 3 2 54.3.
1 5 8 7 9 1 0 6 4 3 2 54.3 Tie{z
3. 1 5 8 6 1 0 9 7 4 3 2 57.1 Genome with In 5a
1 1 3 7 9 8 6 1 0 5 2 4 60.3 Tie [1'
.
1 3 7 1 0 6 8 9 5 2 4 60.3 1 3 7 5 2 6 1 0 9 8 4 60.5
Tie
{z:
1 3 7 5 2 8 9 10 6 4 60.5 3. 1 3 4 2 5 1 0 6 8 9 7 64.1 Genome with In 7-3717TABLE 2
Best sequences for variance of differences
Genome Rank Chromosome sequence Variance of differences
1. 1 3 4 8 9 10 7 5 2 6 7.675 2. 1 3 4 8 10 9 7 5 2 6 8.614
Normal genome 3. 1 3 4 8 9 10 6 2 5 7 9.540
4. 1 3 4 2 5 7 10 9 8 6 9.863 5. 1 3 4 2 5 7 9 10 8 6 9.915
1. 1 5 7 6 10 9 8 4 3 2 9.078 2. 1 5 9 10 6 7 8 4 3 2 9.917
Genome with In 5a 3. 1 5 7 4 3 6 10 9 8 2 10.372
4. 1 5 7 8 9 10 6 4 3 2 10.855 5. 1 5 7 4 3 8 9 10 6 2 11.842
1. 1 3 4 8 9 1 0 6 2 5 7 8.060 2. 1 3 4 8 9 5 2 6 10 7 9.794
Genome with In 7-3717 3. 1 3 7 5 2 6 10 9 8 4 9.992
4. 1 3 4 6 10 9 8 2 5 7 10.370 5. 1 3 7 10 6 2 5 9 8 4 12.148 specificity of telomere association to ensure that the same sequence will prevail in both genomes? It seems to me that the variability in BENNETT'S observations do not encourage great confidence in this approach either.
178
specialized capabilities of telomeres or centromeres (regardless of arm length considerations) fails to account for the high frequency of observed or inferred pairing of rearranged segments with their normal sequence counterparts in such cases as ring chromosomes (SCHWARTZ 1953) and insertional translocations (MAGUIRE 1965) and for intercalary pairing partner switches in the multivalents of polyploids and polysomics (HENDERSON 1969) (where more than two copies of homologues are present). Also, normal recombination frequency has been reported in the vicinity of a homozygous terminal deficiency (RHOADES 1978), where the normal telomeres are missing in both members of a pair of homo- logues. Additional evidence for a major role of intercalary chromosome parts in the initiation of homologous pairing has been provided by studies of recombi- nation frequencies in Drosophila melanogaster reciprocal translocation hetero- zygotes (HAWLEY 1980) and by cytogenetic studies of maize trisomes and tetraploids heterozygous for a paracentric (within arm) inversion, where pref- erential pairing of internally sequentially matched chromosome arms has been noted (DOYLE 1963). The latter observation suggests that remnant anaphase orientation may contribute to the selection of pairing partner where choice exists (with the presence of three or four homologues per cell), and that pairing effective for crossing over was inspired internally within the inverted arm.
Another aspect deserves consideration. It seems likely that, if special affinities of telomeres for each other or for the inner nuclear envelope serve valuable metabolic roles, their function probably does not depend importantly on arm length. Organisms carrying chromosome rearrangements that drastically alter arm length are frequently phenotypically indistinguishable from their normal sequence siblings, as was the case with the plants studied here. It is even possible that the tendency, which has been observed for some consistent spatial chromosome arrangement, is a by-product consequence of remnant anaphase orientation and even perhaps physical properties of chromosomes (such as arm length or mass interactions with the spindle) and does not necessarily serve a function per se (COMINGS 1980). It is also conceivable that telomere association to the inner nuclear envelope somehow serves the special needs of this portion of chromosomes at chromosome replication. But even if specific telomere affinities with other telomeres are commonplace, and they indeed contribute somehow to the establishment of a necessary order within the nucleus, they, nevertheless, appear to fall far short of providing a basis for the general mechanism of homologue pairing at meiosis. It is hoped that BENNETT’S remark- able observations will inspire the development of even more ingenious and sophisticated approaches to the study of the general mechanism of homologue pairing. A profitable direction might involve a concerted search for interaction of intercalary chromosome regions with intranuclear or intraspindle compo- nents.
Special thanks are due J. R. RESPESS for the computer modeling. I am also very grateful to D. A.
LITERATURE CITED
ASHLEY, T., 1979 Specific end-to-end attachment of chromosomes in Ornithogalum virens. J. Cell Sci. 38: 357-367.
AVIVI, L. and M. FELDMAN, 1980 Arrangement of chromosomes in the interphase nucleus of plants. Hum. Genet. 5 5 281-295.
BENNETT, M. D., 1982 Nucleotypic basis of the spatial ordering of chromosomes in eukaryotes and the implications of the order for genomic evolution and phenotypic variation. pp. 239-261. In: Systematics Association Special Volume No. 20: Genome Evolution, Edited by G. A. DOVER and R. B. FALVELL. Academic Press, New York.
BURNHAM, C. R., J. T. STOUT, W. H. WEINHEIMER, R. V. KOWLES and R. L. PHILLIPS,
COMINGS, D. E., 1980 Arrangement of chromatin in the nucleus. Hum. Genet. 5 3 131-143. DOYLE, G. G., 1963 Preferential pairing in structural heterozygotes of Zea mays. Genetics 48:
FABERG&, A. C., 1942 Homologous chromosome pairing: the physical problem. J. Genet. 43 121-
HAWLEY, R. S., 1980 Chromosomal sites necessary for normal levels of meiotic recombination in Drosophila melanogaster. I. Evidence for and mapping of the sites. Genetics 94: 625-646. HENDERSON, S. A., 1969 Chromosome pairing, chiasmata and crossing over. pp. 326-357. In:
Handbook of Molecular Cytology, Edited by A. LIMA-DE-FARIA. American Elsevier, New York.
LEWIN, R., 1981
MAGUIRE, M. P. 1965 The relationship of crossover frequency to synaptic extent at pachytene in
MAGUIRE, M. P., 1974 A new model for homologous chromosome pairing. Caryologia 27: 349-357. MAGUIRE, M. P., 1982 Evidence for a role of the synaptonemal complex in provision for normal
MOENS, P. B., 1969 The fine structure of meiotic chromosome polarization and pairing in Locusta
NEUFPER, M. G., L. JONES, and M. S. ZUBER, 1968 The Mutants of Maize. p. 4. Crop Science Society
PARVINEN, L. M. and K. 0. SODERSTROM, 1976 Chromosome rotation and the formation of synapsis.
RHOADES, M. M., 1978 Genetic effects of heterochromatin in maize. pp. 641-671. In: Maize
SCHWARTZ, D., 1953 The behavior of an x-ray-induced ring chromosome in maize. Am. Nat. 8 7
ZICKLER, D., 1977 Development of the synaptonemal complex and the “recombination nodules” during meiotic prophase in the seven bivalents of the fungus Sordaria macrospora. Auersw. Chromosoma 61 289-316.
Corresponding editor: S. WOLFF 1972 Chromosome pairing in maize. Genetics 7 1 111-126.
1011-1027.
144.
Do chromosomes cross talk? Science 214 1334-1335.
maize. Genetics 51: 23-40.
chromosome disjunction at meiosis I1 in maize. Chromosoma 84: 675-686.
migratoria spermatocytes. Chromosoma 28: 1-25.
of America, Madison.
Nature 260: 534-535.
Breeding and Genetics, Edited by D. B. WALDEN. John Wiley and Sons, New York.