THE NATURE AND ORIGIN OF MUTATIONS WHICH BLOCK A TEMPORAL SEQUENCE FOR GENIC EXPRESSION I N PARAMECIUM
R. W. SIEGELl A N D JUNE COLE2
Department of Zoolo,gy, University of California, Los Angeles, California 90024
Received November 28, 1966
HERE is a temporal sequence for the expression of two genetically independ- Tent loci in Paramecium bursaria. This sequence was brought to light by demonstrating differences in the competence for mating among cells derived at successive intervals during the vegetative growth of exconjugant clones ( SIEGEL and COHEN 1963). The junctional linkage of the two genes was suggested by the discovery that a mutation a t one of the two loci blocks the expression of the second locus so that the normal sequence is permanently interrupted (SIEGEL
1965). We present here a detailed account of the new mutation together with experiments which bear on the special circumstances under which it may arise. In the next paragraphs we briefly summarize the previous results basic to these problems.
Conjugation between cells belonging to complementary mating types is the only form of sexual reproduction in P. bursaria. All of the stocks of syngen 1
belong to one or another of four mating types which are determined by two independent loci. Genes at these loci have been shown to control the production of the mating-type substances appearing on the cell surfaces. The dominant and recessive alleles at the A locus are responsible for one pair of mutually comple- mentary ciliary proteins A and a, and the alleles at the B locus determine a second such pair of complementary ciliary proteins,
B
and b. Each of the four mating types synthesizes a unique combination of these substances(AB,
Ab, aB and ab) whose interactions-A:a or B:b or both-are basic to the specific cellular adhesions leading to mating and conjugation (METZ 1954; COHEN and SIEGEL 1963).Following the origin of a new clone, the two loci concerned with the mating- type substances come to expression in the course of a rather well defined period (normally about 50 cell doublings) of vegetative reproduction. Early in this period neither of the two mating-type loci is expressed; then one locus alone comes to expression and, finally, both loci are expressed. It is to be stressed that these changes in cell phenotype occur during the expansion of clones under con- stant environmental conditions. For this reason the pattern of sequential gene expression cannot be a reflection of a systematic series of environmental cues, and the regulation of cellular differentiations must be an entirely intracellular
1 Supported by grants from the National Science Foundabon and by Cancer Research Funds of the University of
a Supported by Pubhc Health Servit:e Training Grant 5-TI-AI-70. California
608 R. W. SIEGEL A N D J . COLE
process. In agreement, each exconjugant clone normally passes through one or the other of two alternative developmental sequences, each of which appears to be under genetic control. Some clones express the A locus first, then A and B together while others express the B locus first, then A and B. The experimental evidence for these statements has been presented in papers by SIEGEL and COHEN
(1963) and SIEGEL (1965). The picture of the genetic regulation of the temporal sequences is further clarified by a description of the exceptional clones central to the present studies. In these mutants, the normal intraclonal temporal se- quence, B+A+B, is blocked following the expression of the B-locus. Since the mutation appears at the B locus itself, we have been led to conclude that this region must serve to regulate the normal expression of the A locus and also par- ticipate in the production of its own mating-type substance.
While the vast majority of crosses always yield progeny which are in com- plete accord with the simple two-locus hypothesis for the determination of the four mating-type specificities, others characteristically provide unexpected results
(SIEGEL 1963). Data concerned with the nature and origin of exceptional clones are of interest at least in so f a r as they suggest that genic instability, possibly classified with either gene-conversion a r paramutation-type phenomena, may occur in yet another organism.
MATERIALS A N D M E T H O D S
The general relationship between the four mating types, their genetic control, and the mating-type substances found on the cell surfaces is given below:
Mating type Genotypes Mating-type substances
I AABB, AABb, AaBB, AaBb AB
I1 aaBB, aaBb aB
I11 aabb ab
IV AAbb, Aabb Ab
The designations, origins, and mating-type determining genes of the stocks used are as follows:
Stock and mating type 8-1 45-1 25-11 32-111 3-IV 137-111 29-11 31-11 46-11 56-11 8-11 23-11 Mating-type
Origin genes
Malibu Lake, Calif. AdBb
Cottonwood Stream, Calif. AaBB
Malibu Lake, Calif. aaBb
Malibu Lake, Calif. aabb-a
Malibu Lake, Calif. Aabb
Segregant from 25-11 x 3-IV aabb Segregant from 25-11 x 3-IV backcrosses aaBB
Same as 29-11 aaBB
Segregant from 8-1 x 3-IV backcrosses aaBB
Same as 4 - 1 1 aaBB
Segregant from 8-1 x 25-11
Same as 8-11
aaBB
aaBB
Comments
Homozygous for allele B
Homozygous for allele B
Homozygous for allele B
Homozygous for allele B
Possesses isoalleles ( 1 )
Possesses isoalleles ( 1 )
from stock 25
from stock 25
from stock 8
from stock 8
B from stocks 8 and 25
G E N E E X P R E S S I O N I N P A R A M E C I U M 609
It should be pointed out that when two stocks are derived as segregants from a given cross and selected only on the basis of l;he alleles at the B locus, they must not be considered as otherwise genetically identical. M e t h d : j for the culturing, crossing, and scoring of stocks and their sexual progeny follow those of SONNEBORN (1950) with minor modifications as noted previously (SIEGEL
1963).
The mating-type designation 111-IV indicates that the clone will conjugate with tester cells of types I and I1 but never with those of types 111 and IV. Many entirely normal clones express the 111-IV phenotype for a brief period before becoming type I11 or type IV and these must not be confused with “permanently adolescent” clones. Since normal adolescents remain type 111-IV for less than 20 fissions, clones which were 111-IV for 50 or more fissions were scored as perma- nent adolescents. Several such clones have been maintained for over three years, and at least 600 fissions, and have given no indication of further differentiation.
In the majority of crosses to he reported, large numbers of firmly united conjugating pairs were singly isolated from mass matings of appropriate cultures; then a single line of descent was started from each pair, allowed to reproduce at maximal fission rate by serial reisolations, and the left-overs were tested a t various intervals for mating type. But i n other crosses, systematic variations within the progeny of a given pair were sought; thus the parental cells were unmis- takably “marked’ by the presence or abscence of symbionts (SIEGEL 1960) to permit the reisola- tion and identification of the cytoplasmic origins of the two exconjugant cells from each pair. The two exconjugants were allowed to undergo one more fission and the resulting four cells, each the progenitor of a new caryonide, were again isolated. The four caryonides, called a syn- clone, were then expanded and tested as noted above.
RESULTS
The
production of exceptional clones: According to the genetic determination of mating types, all of ithe progeny of the cross of typeI1
(aaBB) by type I11 (aabb) should be type I1 (aaBb). But in particular crosses, two kinds of excep- tional clones were produced in low frequency; the first kind of exceptional mani- fests mating-type 111; the second sort of exceptional mates with typesI and
TABLE 1
The distribution of exceptio.wl clones among independent crosses of stock 29-11 to stocks 32-111
and 137-111 (Ezcepiionals appear in boldface)
Progeny
Cross Date I I1 I11 IV 111-IV
29 x 32 June 1963 0 114 2 0 2
Nov. 1963 0 83 2 0 0
Mar. 1964 0 22 0 0 1
Apr. 1964 0 67 2 0 0
July 1964 0 83 0 0 0
Mar. 1966 0 316 1 0 2
0 685 7 0 5
Totals
29 x 137 Nov. 1963 0 106 0 0 0
Feb. 1964 0 158 0 0 0
Mar. 1964 0 32 0 0 0
Apr. 1964. 0 82 0 0 0
July 1964 0 97 0 0 0
Mar. 1966 0 376 0 0 0
610 R. W. SIEGEL A N D J. COLE
TABLE 2
The appearance of exceptional clones in crosses of diverse stocks to stock 32-111
Stock and genotype*
56-11 aaB8B8
46-11 aaB8B8
29-11 aaBe5B25
31-11 MB95B25
8-11 MB25B8
23-11 aaBS5B8
I I1 I11 IV 111-IV
0 113 0 0 1
0 457 0 0 2
0 685 7 0 5
0 128 1 0 0
0 91 0 0 0
0 116 2 0 0
* Superscripts denote the stocks from which the B alleles were de?ived.-See MATERIALS AND METHODS.
11, never with I11 or IV, and is referred to as type 111-IV. The previous conclusion
(
SIEGEL
1965) that such exceptionalF,
lines appear when stock 32 is used as the type I11 parent but never when stock 137 is crossed is fully confirmed by the more extensive results shown in Table 1. The data in Table 2 establish that stock 29 as well as other stocks can participate with stock 32 in the formation of exceptionals.The four caryonides descended from a single pair of conjugants-a “synclone” -are normally genetically identical and express the same mating type ( SIEGEL
and LARISON 1960; SIEGEL 1963). The question arises as to whether the rule of synclonal uniformity applies to those rare pairs which yield exceptional type-I11 and 111-IV progeny. For if it does, then a premeiotic or meiotic event is probably responsible for exceptionals; if not, then the event probably occurs during the postzygotic development of clones descended from stock 32. A total of 317 syn- clones from the cross 32-111 x 29-11 were tested in this connection. As shown in Table 3, five gave at least one exceptional caryonide. The two cases listed at the bottom of the table are clear exceptions to the rule of synclonal uniformity strongly suggesting that exceptionals arise after fertilization. The remaining three cannot be interpreted from this standpoint because of inviability. The pa- rental clones for this study were “marked” by the presence or absence of sym- bionts so that the cytoplasmic origins of exceptional caryonides could be deter- mined. It is significant that in the two cases in ‘which sister caryonides of type
TABLE 3
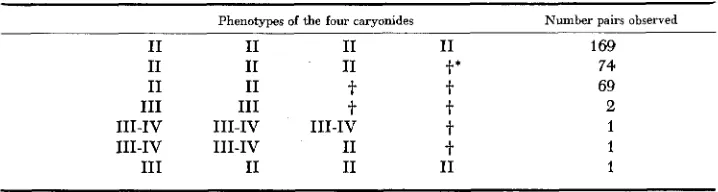
Stock 29-11 x Stock 32-111; the phenotypes of the four caryonides from each of 317 pairs
Phenotypes of the four caryonides Number pairs observed
I1 I1 I1
I1 I1 I1
I1 I1
t
I11 I11
t
111-IV 111-IV 111-IV
111-IV 111-IV I1
111 I1 I1
I1 169
t*
74t
69t
2t
1t
1I1 1
GENE EXPRESSION I N PARAMECIUM 61 1
I11 were found, the exceptional lines were the cytoplasmic descendents of a 29-11
parental cell. Hence these exceptionals, and presumably others, do not arise as nonconjugants, nor can they be readily explained away as cases of such forms of uniparental nuclear reorganization as cytogamy o r macronuclear regeneration (SONNEBORN 1947). Experiments to be described later lead to the same conclu- sions. But before considering further the various possible explanations for the origin of exceptional clones, the genotype of stock 32 and the genotypes of the ex- ceptional clones themselves need to be set forth.
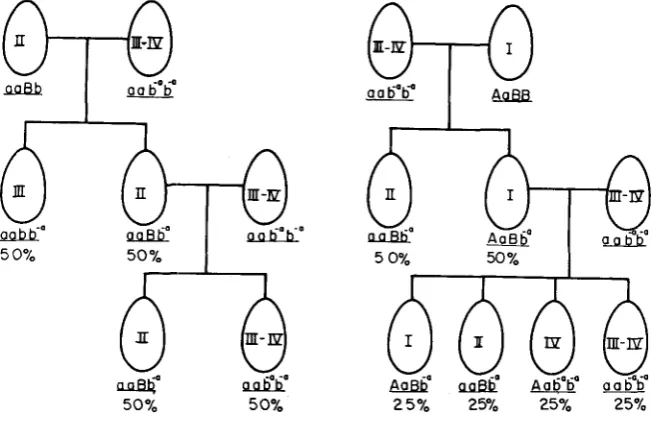
The genotypes of exceptional clones: The exceptional clones shown in Tables 1 and 2 have been repealtedly tested for phenotype during vegetative reproduction and no instance of instability has been found. Hence the macronuclear genes ap- pear to be stable in expression. Breeding analyses of the exceptional 111-IV clones, depicted in Figure 1, have revealed their micronuclear genes. In crosses to stock
25-11 (aaBb), the progeny consisted of types I1 and I11 in a 1 : 1 ratio (12 to 16) ;
backcrosses of the F, type
I1
clones to the parental 111-IV yielded only types I1and 111-IV, again in a 1 : 1 ratio (54 to 46). Crosses of the 111-IV clones to stock
45-1 (AaBB) produced types I and I1 in the anticipated 1: 1 ratio (22 to 23), and in the backcross, types I, 11, IV, and 111-IV occurred in approximately equal numbers (46:21:36:30; P
>
0.02).If an allele at the A locus is postulated as the basis for the 111-IV phenotype, then the backcross shown on the left of Figure 1 would be expected to yield mat- ing types I1 and I11 in equal numbers together with new permanent adolescent clones of the type 1-11; in fact neither type I11 nor type 1-11 clones were found. Moreover, this postulate predicts that the backcross shown on the right in Figure 1 will not yield mating-type I1 clones but, on the contrary, this phenotype did appear. Hence, the genic basis for type 111-IV cannot reasonably be located at
612 R. W. SIEGEL A N D J. COLE
TABLE 4
The properties of the three B-locus alleles
Mating type Effect on
Allele Dominant to: substance formed A locus
-
Aa Expressed
aa Expressed
B b, b* B
Aa Expressed aa Expressed
Aa Expr-ssed
aa Inhibited
b 6” b
b-a b
the A locus. Let us suppose next that the gene responsible for 111-IV clones is at neither the A nor the B locus. Then types I11 and 1-11 would be expected from the first backcross and types I11 and 1-11 would also be expected among the clones produced by the second of the backcrosses. Since these phenotypic classes were absent in the two crosses, there is not support for this assumption. Finally, if the gene for type 111-IV behaves as a recessive allele at the B locus then all of the data are accounted for in a satisfactory *way.
The results shown on the last line of Table 5 include two new exceptional lines, and the interpretation of these will be presented in the DISCUSSION. With this
reservation, clones of type 111-IV will henceforth be considered homozygous for the new allele b” (“b m i n u s a”). The properties of alleles B, b and b“ are com- pared in Table 4.
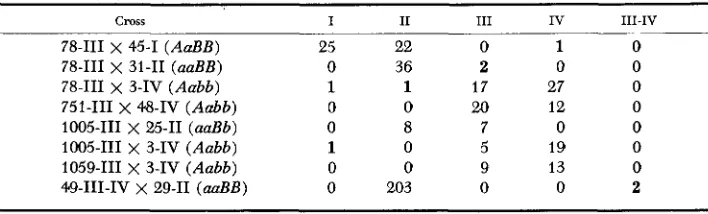
In order to determine the micronuclear genotypes of the exceptional-I11 clones, the progeny of crosses to standard stocks 45-1 (AaBB), 25-11 (aaBb) and 3-IV (Aabb) were examined for mating type. In general, the results shown in Table 5 suggest the genotype aabb for the exceptionals, consistent with the phenotypes
of the clones. On the other hand, a small fraction of the progeny are themselves exceptional-these appear in boldface-and will be discussed in more detail later. Although the foregoing results are consistent with the genotype aabb for the
TABLE 5
The progeny of five exceptional clones ( N e w exceptional clones appear in boldface)
Cross I I1 I11 IV 111-IV
78-111 X 45-1 (AaBB) 25 22 0 1 0
78-111 x 31-11 (aaBB) 0 36 2 0 0
78-111 X 3-IV (Aabb) 1 1 17 27 0
751-111 X 48-IV (Aabb) 0 0 20 12 0
1005-111 x 25-11 (aaBb) 0 8 7 0 0
1005-111 x 3-IV (Aabb) 1 0 5 19 0
1059-111 x 3-IV (Aabb) 0 0 9 13 0
G E N E E X P R E S S I O N I N P A R A M E C I U M 61 3 exceptional type-I11 clones, they do not test the adequacy of two possible alterna- tive genotypes; the exceptional-111’s could be heterozygotes aabb“, or they might be hemizygotes aab-, in which the B locus (or chromosome) is deleted. To ex- plore these possibilities, an exceptional-I11 was crossed to stock 3-IV (Aabb) and eight of the resulting FPl clones were backcrossed to the parental exceptional-111.
If the exceptional-I11 is aabb”, then type III-IV (aab”b-“) should segregate among the backcross progeny; if it is aab-, then types 11-111 (aa--) and I-IV
(Aa- -) should appear following the backcross.
In fact, no such permanently adolescent clones were found among a total of 209 backcross clones. Since a similar analysis of another exceptional type-I11 yielded no permanent adolescents among 166 backcross clones, it can be assumed that these exceptional-lWs are genotypically aabb.
The genetics
of stock 32-111: Exceptional clones are uniquely associated with crosses involving stock 32; for this reason any attempts to explain the origin of these exceptionals requires an adequate description of the genetic constitution of this stock. The majority of the Fl’s from crosses of stock 32 to aaBB are type I1(see Tables 1 and 2) amd, presumably, aaBb. However, when a random sample of 30 such F, clones was testcrossed to a clone of type III-IV, previously estab- lished as aab-“b+, 20 vvere found to be aaBb and 10 were aaBb”. These results make it perfectly clear that stock 32 can contribute either of the alleles b or b-“
to a cross, and hence its micronuclear genotype is probably aabb”.
614 R. W. SIEGEL A N D J. COLE
progeny, eight were III-IV, but no cross gave two III-IV clones. In sum then, these observations-together with the fact that stock 32 is the product of a single cell iso- lated from a natural source-strongly suggest that stock 32 is a homogeneous population with the genotype aabb”.
Do normal conjugants produce exceptional clones? Although illegitimate mat- ing-intraclonal conjugations-is known to be extremely rare in P . bursaria
(LARISON
and SIEGEL 1961),
occasional conjugations among cells of stock 32-occurring, say, in a mixture of stock 32
x
stock 2 9 4 o u l d produce the excep- tional clones. To determine the incidence of 32 x 32 conjugations in a mass mat- ing between stocks 32 and 29, an asymbiotic sample of stock 29 was mixed with normally symbiotic cells from stock 32, then all of the pairs to form were isolated, and the parental origins of the two mates for each pair were determined on the basis of cell color. Since every one of the 1057 pairs was found to consist of a white cell (stock 29) and a green cell (stock 32), illegitimate conjugation can hardly account for the origin of the exceptional lines. This conclusion is strengthened by the analysis of exceptional synclones presented earlier.Normal conjugation involves the formation of isogenic haploid male and fe- male pronuclei in each mating cell followed by the exchange of male pronuclei and their subsequent union with the “stationary” female nuclei. One could ac- count for the exceptional clones by supposing that cells of stock 32 may induce the effective loss of pronuclei (or the genetic material) contributed by cells of other stocks, so that new clones arise either by an autogamous nuclear reorgani- zation or by the diploidization of gametic nuclei derived from stock 32. Such ab- normal forms of uniparental inheritance would be expected to yield the observed aabb and aab”b-” exceptional genotypes. This explanation leads to the prediction that if genetically damaged, and hence virtually sterile, cells from stock 29 are mated to stock 32, the new clones will include the exceptional types I11 and III-IV in just the same numbers as found in matings of normal cultures because in both crosses, the origin of exceptionals is independent of the genetic function of the stock 29 pronuclei. In practice, a cell isolated from a subculture of stock 29 which had been exposed to 100,000r from a cobalt-60 source was expanded into a mass culture and then mated to stock 32. Of 1140 mating pairs isolated, only 69 yielded viable progeny. Since the normal viability of the 29 x 32 cross is about 50%, the fact that 12 exceptional clones were found among 685 progeny (see Table 1) means that this cross can be expected to produce seven to 19 exceptional lines among a total of about 1400 conjugations (P = 0.05). But no exceptional clones were found among the 69 survivors of the experimental cross. Since at least six would be expected if nuclei from stock 32 are alone responsible for the origin of exceptionals, it must be concluded that both mates in a conjugating pair con- tribute to and participate in the formation of an exceptional clone.
(GENE EXPRESSION I N P A R A M E C I U M 615
arisen by illegitimate mating (45 X 45, AaBB x AaBB; 78 X 78, aabb X aabb) or
by some other form of uniparental reorganization.
DISCUSSION
The crosses which have been presented describe the properties of a new allele at the B mating-type lo~cus in P. bursaria. This gene, b”, controls the production of the mating-type substance b and, according to present methodology, in this respect its activity is indistinguishable from that of its allele b. Since heterozy- gotes Bb and Bb” form mating-type substance B but not b, the gene B is dominant to both its alleles. But while genes B and b permit the full and normal expression of the A-locus, b” does not. Most importantly, the fact that b” is recessive to alleles B and b in this respect probably means that the mutant does not produce a repressor for the A locus and that for the developmental sequence B+ASB genes B and b could serve as activators or derepressors.
Perhaps the most remarkable property of the mutant is revealed by the fact that only one of the alternative alleles known for the A locus is suppressed by
b-”b* genotypes; the function of the genotype aa, leading to the formation of mating-type substance EL, is lost whereas that of the genotype Aa, which normally
produces substance A, is not.
The basic question n i t h which these studies began, “What are the genotypes of clones which are penmanently adolescent?”, has been answered at the level of the classical gene; they are aab”b-“. Thus we are led to conclude that the B locus is in some way concerned with the two mating-type substances and that the two loci which show genetic: independence (separate chromosomes) are functionally related. But a deeper understanding of the intergenic regulation which has been brought to light demands additional study and, perhaps, new methods. Most importantly, we need to learn whether the two functions attributable to the B
locus-synthesis of B or b mating-type substance and regulation of the A locus- depend upon the activities of two closely linked and independently mutable cis- trons o r upon a single cistron which serves (in some way) two functions.
A discussion of the e vent responsible for the occurrence of exceptional clones should begin by focusing attention on two important facts. The first of these is that the unexpected phenotypes I11 and III-IV have a genic basis. The B locus, or an extremely closely linked locus, is involved in all exceptions so far studied; while most of the descendants from the cross aabb” X aaBB belong to the expected classes aaBb and aaBb-”, a few are phenotypically exceptional and are geno- typically aabb or aab-b-“. Second, these exceptionals appear only among the progeny of crosses invcllving stock 32 or among the progeny of clones derived from stock 32. The foregoing points serve to rule out certain plausible explana- tions for the phenomenon. For example, genetic segregation would account for the appeareance of exceptional clones in the cross 32 x 29 if it could be shown that stock 29 carries-but only rarely contributes for some unknown reason-the
alleles b and b”. Or it might be imagined that stock 29 can form alleles b and b-“
616 R. W. SIEGEL A N D J. COLE
(prezygotic). However, the cross 137 X 29 never yields either type I11 or type III-IV; all of the progeny are aaBb. This establishes the genotype of stock 29 as aaBB and hence eliminates these possibilities. Nor is any hypothesis which requires segregation consistent with the observation that only some of the de- cendants of a given pair of conjugants may be exceptional. Parenthetically, that observation virtually excludes nondisjunction or attached chromosomes in stock 32 as the basis for the exceptionals. The reasonable assumption that stock 29 can- not form alleles b and b” by ordinary crossing over or mutation together with the fact that exceptional lines may originate from the stock 29 parental cell in matings to stock 32 leads to the conclusion that these exceptions cannot be a consequence of uniparental inheritance (i.e., cytogamy, macronuclear regeneration, noncon- jugation, or illegitimate mating) ; this strongly infers that both parental cells participate in the event which leads to the production of exceptionals. This point is considerably strengthened by other kinds of results all of which indicate that exceptionals arise in the course of normal conjugations. In sum, it is difficult to escape the conclusion that exceptional lines are the products of biparental inheri- tance followed by some unorthodox event (s) leading to the apparent loss of allele B from stock 29, and the concommitant doubling of either b or b”, from stock 32. The kinds of exceptionals produced upon crossing exceptionals to standard stocks (see Table
5 )
place some restrictions on the characterization of the un- orthodox event noted above. First, in sharp contrast to the standard type I11(stock 137 aabb) exceptional-111’s have been found to yield mating types I and I1
in crosses to stock 3-IV ( A a b b ) ; occasionally then they contribute the allele B to progeny clones. This probably means that the original gene B is not simply deleted but rather that its function has been metastably altered. Granting the interpretation of a change from B to b followed by rare reversions of the orgi- nated allele to B, one should like to test for reversions of new b” alleles to B. Unfortunately, type III-IV exceptionals can only be crossed to clones which already carry the allele B, and thus the direct test is impossible. Second, excep- tional-111’s differ from the standard type I11 in yet another way; when excep- tionals are crossed to stocks which are AaBB or aaBB new exceptionals of types
I11 and IV appear and for this reason new changes of B to b must be presumed. Similarly, a cross of type III-IV to aaBB yielded two new type III-IV clones and, apparently, new substitutions of b” for B. A reasonable interpretation of these observations is that (1) changes in the allele B occur and (2) the changes are specified by the allele brought to the cross by either stock 32 or an exceptional clone descended from stock 32. More precisely, the change of B to b is conditioned by the presence of b and the change B to b” is sponsored by b”. In agreement, it will be recalled that the exceptionals formed in the interstock cross 32 x 29 are either aabb or aab”b-”, never aabb-”. Moreover, the data on the distribution of exceptionals within synclones (Table 3) support this interpretation in so far as it was found that either type I11 or type III-IV, never both, appear among the progeny of a given pair.
GENE EXPRESSION I N PARAMECIUM 61 7 with special emphasis on corn. While the preliminary results on P. bursaria indi- cate a relationship to the kinds of phenomena noted by COE, it seems premature to speculate on alternative mechanisms which could account for our present observations.
SUMMARY
Certain clones of P. bursaria are exceptional in that they mate with two rather than three of the four standard mating types. Exceptionals of mating-type 111-IV
(these mate with types I and 11) were subjected to a breeding analysis and it was concluded that they are homozygous for a new allele at the B locus, b”.-During the life history of wild-type clones, the A and B loci which are concerned with mating-type determination come to phenotypic expression in a temporal sequence WB+B+A. But i n b” homozygotes this sequence is permanently blocked after the B locus has been expressed. A small but consistent number of phenotypically and genetically exceptional progeny are formed when stock 32 (aabb”) is crossed to other stocks which #are aaBB. Some exceptionals are type 111-IV and geno- typically aab-b-”; others are type 111, aabb. Experiments designed to uncover the basis for these clones suggest that gene B can undergo specific and metastable modifications; it appeal-s to change to b in the presence of allele b from stock 32 and to b” in the presence of b” from stock 32. On the other hand, B is stable when it is present with the alllele b derived from stock 137.
LITERATURE CITED
COE, E. H., JR., 1966
COHEN, L. W., and R. W. !$IFGEL, 1963
LARISON, L. L., and R. W. SIEGEL, 1961
METZ, C. B., 1954
SIEGEL, R. W., 1960
The properties, origin, and mechanism of conversion-type inheritance at
The mating-type substances of Paramecium bursaria.
Illegitimate mating in Parameicum bursaria and the
Mating substances and the physiology of fertilization to ciliates. pp. 284-334.
Sex in Microorganisms. Edited by D. H. WENRICH. Am. Assoc. Adv. Sci., Washington, D.C. Hereditary endosymbiosis in Paramecium bursaria. Exptl. Cell Res. 19: 239-252. - 1963 New results on the genetics of mating types in Paramecium bursaria. Genet. Res. 4: 132-142. __ 1965 Hereditary factors controlling development in Para- mecium. Brookhaven Symp. Biol. 18: 55-65.
A temporal sequence for genic expression: cell differ- entiation in Paramecium. Am. Zool. 3: 127-134.
The genic control of mating types in Paramecium
Recent advances in the genetics of Paramecium and Euplotes. Advan. Genet. 1: 263-358. -- 1950 Methods in the general biology and genetics of Para-
mecium aurelia. J. Exptl. Zool. 113: 87-147. the B locus in maize. Genetics 53: 1035-1063.
Genet. Res. 4: 143-150
basis for cell union. J. Gen. Microbiol. 26: 499-508.
SIEGEL, R.
w.,
and L.w.
<:OHEN, 1963SIEGEL, R. W., and L. L. IARISON, 1960
SONNEBORN, T. M., 1947