Copyright © 2000, American Society for Microbiology. All Rights Reserved.
Randomly Amplified Polymorphic DNA Analysis of
Erysipelothrix
spp.
ALEXANDRE TOMOMITSU OKATANI,1HIDEKI HAYASHIDANI,1* TOSHIO TAKAHASHI,2
TAKAHIDE TANIGUCHI,1MASUO OGAWA,1ANDKEN-ICHI KANEKO1
Department of Veterinary Medicine, Faculty of Agriculture, Tokyo University of Agriculture and Technology, Fuchu,
Tokyo 183-8509,1and National Veterinary Assay Laboratory, Kokubunji, Tokyo 185-8511,2Japan
Received 10 April 2000/Returned for modification 28 July 2000/Accepted 29 September 2000
The usefulness of randomly amplified polymorphic DNA method (RAPD) to identify each species of genus Erysipelothrixand for epidemiological analysis of this genus was studied. Eighty-one strains and 18 random primers were tested. Among the tested primers, the primers NK51 (GGTGGTGGTATC) and NK6 (CCCGC GCCCC) produced noticeable results. The primer NK51 revealed four species-specific RAPD patterns. Of the 66 strains ofE. rhusiopathiae, 64 had the same unique band of 884 bp. Of the 12 strains ofE. tonsillarum, 11 produced a 1,265-bp band. In addition, two strains, previously thought to beE. rhusiopathiae, produced the 1,265-bp band, suggesting that they had been misclassified. One strain ofE. tonsillarumproduced the 884-bp band, suggesting that it too wasE. rhusiopathiae. TheE. rhusiopathiaestrain of serovar 13 produced a 650-bp band, and the strain of serovar 18 produced a clear 420-bp band as well as three weak bands of 1,265, 918, and 444 bp. The primer NK6 revealed 14 RAPD patterns that were not serovar specific. However, different patterns were produced among strains of the same serovar showing that the RAPD method is able to identify the genetic variations of strains of this genus and can rapidly and easily differentiate strains of the same serovar. Based on these results, we concluded that the RAPD method with primers NK51 and NK6 is a rapid and reliable method to identify the species of this genus; we also concluded that this method might be a useful tool for the epidemiological analysis of theErysipelothrixspecies.
Erysipelothrix rhusiopathiae, a gram-positive, slender,
straight or slightly curved rod, is known to be the causative agent of erysipelas in swine and erysipeloid in humans. This bacterium has been isolated from many species of wild and domestic animals in most parts of the world (31).
Until recently, the genus Erysipelothrix was thought to be comprised of only one species,E. rhusiopathiae. However, Ta-kahashi et al. (19, 20) showed that this genus comprises at least two distinct species by the DNA-DNA hybridization:E.
rhu-siopathiae—comprising serovars 1a, 1b, 2, 4, 5, 6, 8, 9, 11, 12,
15, 16, 17, 19, and 21, and type N—andE. tonsillarum —com-prising serovars 3, 7, 10, 14, 20, 22, and 23. Moreover, they inferred that serovars 13 and 18 might be members of a new and separate species (20). Current bacteriological culture methods require at least 3 days to isolate this bacterium and about 10 days to determine its serovars (21). To replace these time-consuming methods, primer pair MO101-MO102, primer pair ER1-ER2 and a four primer pair set (ER1F-ER1R, ER2F-ER2R, ER3F-ER3R, and ER4F-ER4R) were devel-oped by Makino et al. (9), Shimoji et al. (17), and Takeshi et al. (22), respectively, for rapid and direct detection using PCR method.
However, each primer has some drawbacks. The primers MO101-MO102 is genus specific and is unable to differentiate species. The primers ER1-ER2 is able to identifyE.
rhusio-pathiaestrains but is unable to identifyE. tonsillarumstrains
and theE. rhusiopathiaestrains of serovars 13 and 18 due to the lack of amplification products with DNAs extracted from these strains. The primers ER1F-ER1R, ER2F-ER2R,
ER3F-ER3R, and ER4F-ER4R are considered sufficient to identify each species of this genus. However, due to a difference of size as small as 3 bp among the amplification products, it is difficult to differentiate them. Moreover, four amplification tubes per sample are necessary for identification of the species.
On the other hand, recently the randomly amplified poly-morphic DNA (RAPD) method with one random primer has shown the ability to differentiate bacteria and strains of one genus at species level, and even some effectiveness as a tool for epidemiological and taxonomic studies (5–7, 10, 11, 14–16, 24–28, 30). In the study described in this paper we examined the possibility of identifying species and differentiating strains using the RAPD method and the usefulness of this method in epidemiological analysis of genusErysipelothrix.
MATERIALS AND METHODS
Bacterial strains and biochemical tests.The details of the 81Erysipelothrix
strains used in this study are shown in Table 1. These comprise 54 field isolates (8, 12, 13, 18), reference strains for 23 serovars (including subserovars 1a and 1b) and type N, and type strains ofE. rhusiopathiaeandE. tonsillarum(20). Bio-chemical characterization of the strains were made on the basis of the carbohy-drate fermentation patterns, test-tube growth in gelatin medium, production of H2S in triple sugar iron agar (Difco Laboratories) slants, and catalase and
oxidase production, as described previously (18, 20). The carbohydrate fermen-tation test was carried out using nutrient broth supplemented with 1% Andrade’s indicator and 10% horse serum (20, 29). The serovars were identified by the agar gel double-diffusion precipitation technique using the heat-stable antigen ex-tracted from the cell wall of each strain and rabbit antisera representing serovars 1 through 23 of theErysipelothrixspecies (20).
DNA preparation.Total DNAs from the strains listed in Table 1 were
pre-pared using the method described by Makino et al. (9). Briefly, bacterial cells of a 24-h culture were suspended in 200l of TES buffer (50 mM Tris-HCl, 5 mM EDTA, 50 mM NaCl [pH 8.0]), containing 10l of lysozyme (10 mg/ml) and 10
l ofN-acetylmuramidase SG (1 mg/ml). They were then incubated for 30 min at 37°C before the addition of 10l of 10% sodium dodecyl sulfate and 10l of proteinase K (20 mg/ml). After further incubation at 55°C for 60 min, the crude DNA preparation was treated with RNase, extracted three times with phenol-chloroform, precipitated with ethanol, and dissolved in TE buffer (10 mM Tris-HCl, 1 mM EDTA [pH 8.0]). The DNA concentration was determined using the * Corresponding author. Mailing address: Department of
Veteri-nary Medicine, Faculty of Agriculture, Tokyo University of Agriculture and Technology, Saiwai-cho 3-5-8, Fuchu, Tokyo 183-8509, Japan. Phone: 81-42-367-5775. Fax: 81-42-360-8830. E-mail: [email protected] .jp.
4332
on May 15, 2020 by guest
http://jcm.asm.org/
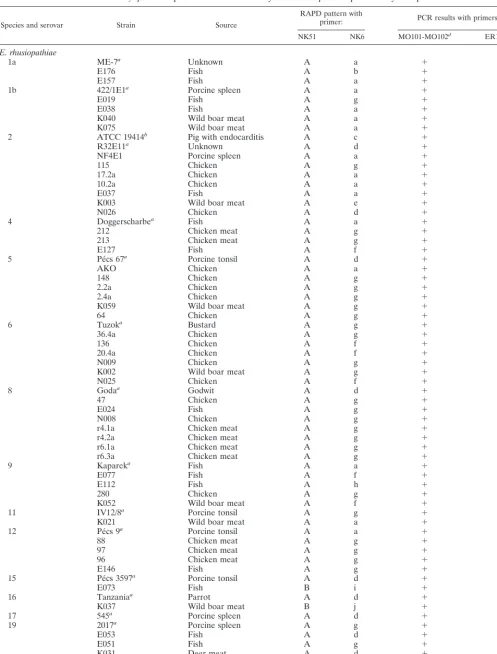
TABLE 1. Erysipelothrixsp. strains used in this study and RAPD patterns produced by each primer
Species and serovar Strain Source
RAPD pattern with
primer: PCR results with primers:
NK51 NK6 MO101-MO102d ER1-ER2e
E. rhusiopathiae
1a ME-7a Unknown A a ⫹ ⫹
E176 Fish A b ⫹ ⫹
E157 Fish A a ⫹ ⫹
1b 422/1E1a Porcine spleen A a ⫹ ⫹
E019 Fish A g ⫹ ⫹
E038 Fish A a ⫹ ⫹
K040 Wild boar meat A a ⫹ ⫹
K075 Wild boar meat A a ⫹ ⫹
2 ATCC 19414b Pig with endocarditis A c ⫹ ⫹
R32E11a Unknown A d ⫹ ⫹
NF4E1 Porcine spleen A a ⫹ ⫹
115 Chicken A g ⫹ ⫹
17.2a Chicken A a ⫹ ⫹
10.2a Chicken A a ⫹ ⫹
E037 Fish A a ⫹ ⫹
K003 Wild boar meat A e ⫹ ⫹
N026 Chicken A d ⫹ ⫹
4 Doggerscharbea Fish A a ⫹ ⫹
212 Chicken meat A g ⫹ ⫹
213 Chicken meat A g ⫹ ⫹
E127 Fish A f ⫹ ⫹
5 Pe´cs 67a Porcine tonsil A d ⫹ ⫹
AKO Chicken A a ⫹ ⫹
148 Chicken A g ⫹ ⫹
2.2a Chicken A g ⫹ ⫹
2.4a Chicken A g ⫹ ⫹
K059 Wild boar meat A g ⫹ ⫹
64 Chicken A g ⫹ ⫹
6 Tuzoka Bustard A g ⫹ ⫹
36.4a Chicken A g ⫹ ⫹
136 Chicken A f ⫹ ⫹
20.4a Chicken A f ⫹ ⫹
N009 Chicken A g ⫹ ⫹
K002 Wild boar meat A g ⫹ ⫹
N025 Chicken A f ⫹ ⫹
8 Godaa Godwit A d ⫹ ⫹
47 Chicken A g ⫹ ⫹
E024 Fish A g ⫹ ⫹
N008 Chicken A g ⫹ ⫹
r4.1a Chicken meat A g ⫹ ⫹
r4.2a Chicken meat A g ⫹ ⫹
r6.1a Chicken meat A g ⫹ ⫹
r6.3a Chicken meat A g ⫹ ⫹
9 Kapareka Fish A a ⫹ ⫹
E077 Fish A f ⫹ ⫹
E112 Fish A h ⫹ ⫹
280 Chicken A g ⫹ ⫹
K052 Wild boar meat A f ⫹ ⫹
11 IV12/8a Porcine tonsil A g ⫹ ⫹
K021 Wild boar meat A a ⫹ ⫹
12 Pe´cs 9a Porcine tonsil A a ⫹ ⫹
88 Chicken meat A g ⫹ ⫹
97 Chicken meat A g ⫹ ⫹
96 Chicken meat A g ⫹ ⫹
E146 Fish A g ⫹ ⫹
15 Pe´cs 3597a Porcine tonsil A d ⫹ ⫹
E073 Fish B i ⫹ ⫺
16 Tanzaniaa Parrot A d ⫹ ⫹
K037 Wild boar meat B j ⫹ ⫺
17 545a Porcine spleen A d ⫹ ⫹
19 2017a Porcine spleen A g ⫹ ⫹
E053 Fish A d ⫹ ⫹
E051 Fish A g ⫹ ⫹
K031 Deer meat A d ⫹ ⫹
21 Ba˜no 36a Sheep dip A a ⫹ ⫹
N MEW22a Unknown A a ⫹ ⫹
Continued on following page
on May 15, 2020 by guest
http://jcm.asm.org/
GeneQuanto pro RNA/DNA Calculator (Pharmacia Biotech ltd, Cambridge, England) and adjusted to the working concentration.
RAPD PCR and gel electrophoresis.Random primers from the collection of
primers available in our laboratory were randomly chosen, and a total of 18 primers were tested to amplify the DNAs. These were 7 10-mer primers (gua-nine-plus-citosine contents of 60 to 100%) and 11 12-mer primers (guanine-plus-citosine contents of 41.7 to 58.3%). The PCR amplification was performed with the Program Temperature Control System PC-700 (Astec Co., Ltd., Tokyo, Japan). The amplification was carried out in a 50-l reaction mixture containing 5l of 10⫻reaction buffer (Sawady Technology Co., Ltd., Tokyo, Japan); 200
M (each) dATP, dTTP, dCTP, and dGTP; 200 nM random primer; 2.5 U of
TaqDNA polymerase (Sawady Technology Co.), and 500 ng of template DNA under a drop of mineral oil. The cycling program was 4 cycles at 94°C for 5 min, 34°C for 5 min, and 72°C for 5 min; 30 cycles at 94°C for 1 min, 34°C for 1 min, and 72°C for 2 min; and a final incubation at 72°C for 10 min. An aliquot of 10
l of the amplified products was subjected to electrophoresis in 2% agarose gel (Iwai Chemicals Co., Tokyo, Japan), stained with ethidium bromide, and pho-tographed under UV light. All the DNA samples were amplified by using the primers MO101-MO102 (9) and ER1-ER2 (17).
RESULTS
Of the 18 primers tested, 15 produced one or more RAPD patterns; among these the primers designated NK51 (GGTG GTGGTATC) and NK6 (CCCGCGCCCC) produced notice-able results.
The primer NK51 produced four species-specific RAPD pat-terns. Three of these patterns were composed of a single band of 884, 1,265, 650 bp and were designated RAPD patterns A, B, and C, respectively. One was composed of a clear band of 420 bp and three weak bands of 1,265, 918, and 444 bp, and it was designated RAPD pattern D. The RAPD pattern A was produced with DNAs extracted from the type strain of E.
rhusiopathiaeand from the reference strains of serovars 1a, 1b,
2, 4, 5, 6, 8, 9, 11, 12, 15, 16, 17, 19, and 21 and type N, believed to be E. rhusiopathiae. The RAPD pattern B was produced with DNAs extracted from the type strain ofE. tonsillarumand from the reference strains of serovars 3, 7, 10, 14, 20, 22, and 23, considered to beE. tonsillarum. The RAPD patterns C and D were produced, respectively, with DNAs extracted fromE.
rhusiopathiaestrains of serovars 13 and 18 (Fig. 1). The same
RAPD patterns, A, B, C, or D, were produced with DNAs extracted from the 54 field isolates; among them, RAPD pat-terns and taxonomic classification of 51 strains were in agree-ment with the classification based on the serovars, as the se-rovar reference strains mentioned above (Table 1). However, three strains showed RAPD patterns that were not in agree-ment with the classification based on the serovars. The strains E073 and K037 of serovars 15 and 16, classified asE.
rhusio-pathiae, showed the RAPD pattern B produced from the type
strain ofE. tonsillarum. The strain K024 of serovar 10, classi-fied asE. tonsillarum, showed the RAPD pattern A produced from the type strain ofE. rhusiopathiae. All 81 strains showed positive results with the primer MO101-MO102. Among the 66
E. rhusiopathiaestrains, 64 showed positive results; 11 of 12E.
tonsillarumstrains and theE. rhusiopathiaestrains of serovars
13 and 18 showed negative results with the primers ER1-ER2. However, in a manner similar to the results obtained with the primer NK51, the three strains described above showed differ-ent results. The strains E073 and K037 had negative results, and the strain K024 had a positive result (Table 1). Of the biochemical tests, these three strains showed the ability to ferment saccharose (data not shown).
The primer NK6 produced 14 RAPD patterns designated as RAPD patterns a through n (Fig. 2). No serovar-specific RAPD pattern was produced. However, different RAPD pat-terns were produced from strains of the same serovars (Table 1). Sixty-fourE. rhusiopathiaestrains showed RAPD patterns varying from a through h, among which the number of bands differed from one to eight, and 11E. tonsillarumstrains showed RAPD patterns j, k, or l, which differed by one band. The strain K024 of serovar 10 showed the RAPD pattern g. TheE.
rhu-siopathiae strains of serovars 13 and 18 showed the RAPD
[image:3.612.56.552.84.290.2]patterns m and n. The strain K037 of serovar 16 showed the RAPD pattern j, and the strain E073 of serovar 15 showed a unique RAPD pattern, i (Table 1).
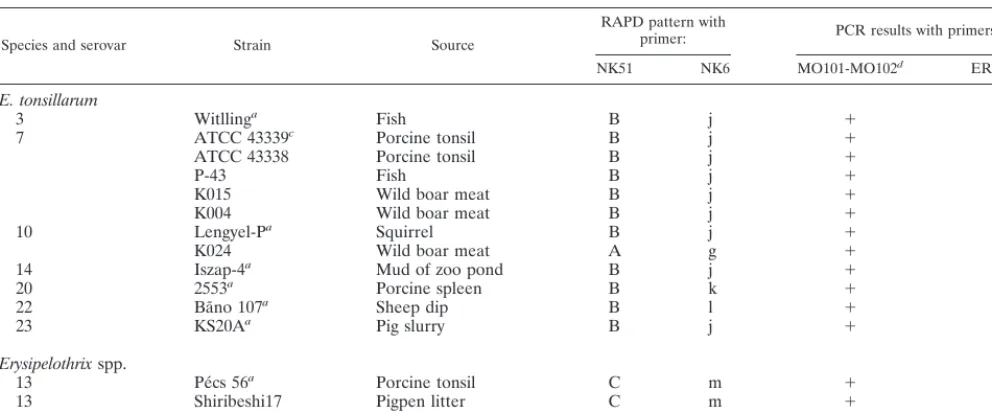
TABLE 1—Continued
Species and serovar Strain Source
RAPD pattern with
primer: PCR results with primers:
NK51 NK6 MO101-MO102d ER1-ER2e
E. tonsillarum
3 Witllinga Fish B j ⫹ ⫺
7 ATCC 43339c Porcine tonsil B j ⫹ ⫺
ATCC 43338 Porcine tonsil B j ⫹ ⫺
P-43 Fish B j ⫹ ⫺
K015 Wild boar meat B j ⫹ ⫺
K004 Wild boar meat B j ⫹ ⫺
10 Lengyel-Pa Squirrel B j ⫹ ⫺
K024 Wild boar meat A g ⫹ ⫹
14 Iszap-4a Mud of zoo pond B j ⫹ ⫺
20 2553a Porcine spleen B k ⫹ ⫺
22 Ba˜no 107a Sheep dip B l ⫹ ⫺
23 KS20Aa Pig slurry B j ⫹ ⫺
Erysipelothrixspp.
13 Pe´cs 56a Porcine tonsil C m ⫹ ⫺
13 Shiribeshi17 Pigpen litter C m ⫹ ⫺
18 715a Porcine spleen D n ⫹ ⫺
aSerovar reference strain. bE. rhusiopathiaetype strain. cE. tonsillarumtype strain.
dPrimers developed by Makino et al. (9). ePrimers developed by Shimoji et al. (17).
on May 15, 2020 by guest
http://jcm.asm.org/
DISCUSSION
In this study, we identified a random primer, NK51, able to produce species-specific RAPD patterns forE. rhusiopathiae
(A), E. tonsillarum (B), and even for the E. rhusiopathiae
strains of serovars 13 (C) and 18 (D), which shows that the RAPD method can be used to identify the species of genus
Erysipelothrix. The results obtained with serovar reference
strains and the 51 (98%) field isolates were in agreement with the classification based on the DNA-DNA hybridization, sero-typing, and biochemical characteristics reported by Takahashi et al. (20). It should be noted that among the 81 strains used, the strains E073 and K037 showed atypical biochemical and molecular biological characteristics, and the strain K024 showed an atypical molecular biological characteristic. If the taxonomic classification of these strains is based only on the serovars, the strains E073 and K037 must be classified asE.
rhusiopathiae and the strain K024 must be classified as E.
tonsillarum. However, studies carried out by Chooromoney et
al. (4) and Ahrne´ et al. (1) using the multilocus enzyme elec-trophoresis and restriction fragment length polymorphisms, respectively, reported that some strains of same serovars can be classified into different clusters, casting doubt on the clas-sification based on serotyping. On the other hand, the RAPD performed with the primer NK51 was able to differentiate and classify these atypical strains, and the results were in agree-ment with the PCR using those previously developed primers, suggesting that they had been misclassified. Thus, strains E073 and K037 might be consideredE. tonsillarum, and the strain K024 might be consideredE. rhusiopathiae. In previous studies, because only strains of serovars 3, 7, 10, 14, 20, 22, and 23, classified asE. tonsillarumbased on the DNA-DNA hybridiza-tion, were able to ferment saccharose, this fermentation ability has been considered a unique characteristic of these serovars and of E. tonsillarum species (20). However, we found the strains E073 and K037 of serovars 15 and 16 were also able to ferment saccharose and had the molecular biological charac-teristic ofE. tonsillarum, and the strain K024 of serovar 10 was able to ferment saccharose but had the molecular biological characteristic of the E. rhusiopathiae species. These results demonstrated that the RAPD method can be used to classify
Erysipelothrixstrains into the genetically correlated species and
[image:4.612.111.495.72.220.2]also suggest that, similar to serotyping, the ability to ferment saccharose is somewhat questionable as a means of identifying the species of this genus. Moreover, as the primer NK51 pro-duced bands from both of the two species, from the strains of serovars 13 and 18, and the characteristic bands of each species differ from each other by more than 200 bp, the identification of these species can be carried out using only one primer and one amplification tube per sample. Thus, this method might be useful in identifying the species of genusErysipelothrix, and it might be easier than the method developed earlier (9, 17, 22). Furthermore, we used the primer NK6 and found that RAPD patterns that differed by one to eight bands were pro-duced with DNAs ofE. rhusiopathiaestrains of the same se-rovar, and RAPD patterns that differed by one band were produced with DNAs ofE. tonsillarumstrains. Moreover, when RAPD patterns of strains E073 and K037 (serovars 15 and 16) FIG. 1. RAPD patterns of theErysipelothrixsp. serovar reference strains produced with the primer NK51. Lanes 1 to 16 show RAPD pattern A (884 bp) produced from theE. rhusiopathiaestrains: lane 1, ME-7 (serovar 1a); lane 2, 422/1E1 (serovar 1b); lane 3, ATCC 19414 (type strain, serovar 2); lane 4, Doggerscharbe (serovar 4); lane 5, Pe´cs67 (serovar 5); lane 6, Tuzok (serovar 6); lane 7, Goda (serovar 8); lane 8, Kaparek (serovar 9); lane 9, IV 12/8 (serovar 11); lane 10, Pe´cs9 (serovar 12); lane 11, Pe´cs3597 (serovar 15); lane 12, Tanzania (serovar 16); lane 13, 545 (serovar 17); lane 14, 2017 (serovar 19); lane 15, Ba˜no36 (serovar 21); lane 16, MEW22 (type N). Lanes 17 to 23 show RAPD pattern B (1,265 bp) produced from theE. tonsillarumstrains: lane 17, Witlling (serovar 3); lane 18, ATCC 43339 (type strain, serovar 7); lane 19, Lengyel-P (serovar 10); lane 20, Iszap-4 (serovar 14); lane 21, 2553 (serovar 20); lane 22, Ba˜no107 (serovar 22); lane 23, KS20A (serovar 23). Lanes 24 and 25 show RAPD patterns C (650 bp) and D (420, 444, 918, and 1,265 bp, sizes from bottom to top), respectively, produced fromErysipelothrixspp. Pe´cs56 (serovar 13) and 715 (serovar 18). Lane M, 1-kb ladder (GIBCO-BRL).
FIG. 2. RAPD patterns obtained from the 81Erysipelothrixstrains with the primer NK6. The strains of lanes a to i areE. rhusiopathiae: lane a, ME-7 (serovar 1a); lane b, E176 (serovar 1a); lane c, ATCC 19414 (type strain, serovar 2); lane d, R32E11 (serovar 2); lane e, K003 (serovar 2); lane f, E127 (serovar 4); lane g, Tuzok (serovar 6); lane h, E112 (serovar 9); lane i, E073 (serovar 15). The strains of lanes j to l areE. tonsillarum: lane j, ATCC 43339 (type strain, serovar 7); lane k, 2553 (serovar 20); lane l, Ba˜no107 (serovar 22). The strains of lanes m and n are, respectively,Erysipelothrixspp. Pe´cs56 (serovar 13) and 715 (serovar 18). Lane M, 1-kb ladder (GIBCO-BRL).
on May 15, 2020 by guest
http://jcm.asm.org/
[image:4.612.63.285.499.652.2]and strain K024 (serovar 10) were compared with those of strains of the same serovar, no similar band was found. These results proved that RAPD is not only able to differentiate the strains of the same serovar but is also able to identify the genetic diversity among strains of these species. Similarities and differences among strains of the same serovar have been described by comparing the cell protein composition using sodium dodecyl sulfate-polyacrylamide gel electrophoresis (2, 3, 23); genetic diversities have been described by multilocus enzyme electrophoresis (4). Our study differs from earlier stud-ies in identifying the strains of these specstud-ies and determining the genetic diversity based directly on the DNA amplification. Based on these results, we conclude that RAPD carried out using the primers NK51 and NK6 might be a rapid and reliable method of identifying the species and strains of this genus; it also might be a useful tool for epidemiological studies of the genus Erysipelothrix. We are now identifying the nucleotide sequences of the amplified products and attempting to design a specific primer able to identify bacteria of this genus and differentiate each species. Furthermore, in view of the genetic diversity, and the similarities of biochemical characteristics among theErysipelothrixstrains, further molecular biological studies with a large collection of isolates of each serovar would be needed to elucidate the taxonomic relationship of the se-rovars with species of the genusErysipelothrix.
REFERENCES
1.Ahrne´, S., I. Stenstro¨n, N. E. Jensen, B. Petterson, M. Uhle´n, and G. Molin.
1995. Classification ofErysipelothrixstrains on the basis of restriction frag-ment length polymorphisms. Int. J. Syst. Bacteriol.45:382–385.
2.Berna´th, S., G. Kucsera, I. Ka´da´r, G. Horva´th, and G. Y. Morovja´n.1997.
Comparison of the protein patterns ofErysipelothrix rhusiopathiaestrains by SDS-PAGE and autoradiography. Acta Vet. Hung.45:417–425.
3.Berna´th, S., G. Y. Morovja´n, V. Sztojkov, and G. Szita.1998. Comparison of
the protein composition ofErysipelothrix rhusiopathiaestrains of subtype 1a isolated from ducks and pigs. Acta Vet. Hung.46:211–217.
4.Chooromoney, K. N., D. J. Hampson, G. J. Eamens, and M. J. Turner.1994.
Analysis ofErysipelothrix rhusiopathiaeandErysipelothrix tonsillarumby mul-tilocus enzyme electrophoresis. J. Clin. Microbiol.32:371–376.
5.Fitzsimons, N. A., T. M. Cogan, S. Condon, and T. Beresford.1999.
Pheno-typic and genoPheno-typic characterization of non-starter lactic acid bacteria in mature Cheddar cheese. Appl. Environ. Microbiol.65:3418–3426.
6.Grayson, T. H., L. F. Cooper, F. A. Atienzar, M. R. Knowles, and M. L.
Gilpin.1999. Molecular differentiation ofRenibacterium salmoninarum
iso-lates from worldwide locations. Appl. Environ. Microbiol.65:961–968.
7.Hayford, A. E., A. Petersen, F. K. Vogensen, and M. Jakobsen.1999. Use of
conserved randomly amplified polymorphic DNA (RAPD) fragments and RAPD pattern for characterization ofLactobacillus fermentumin Ghanaian fermented maize dough. Appl. Environ. Microbiol.65:3213–3221.
8.Kanai, Y., H. Hayashidani, K. Kaneko, M. Ogawa, T. Takahashi, and M.
Nakamura.1997. Occurrence of zoonotic bacteria in retail game meat in
Japan with special reference toErysipelothrix. J. Food Prot.60:328–331.
9.Makino, S., Y. Okada, T. Maruyama, K. Ishikawa, T. Takahashi, M.
Naka-mura, T. Ezaki, and H. Morita.1994. Direct and rapid detection of
Erysip-elothrix rhusiopathiaeDNA in animals by PCR. J. Clin. Microbiol.32:1526– 1531.
10. Makino, S., Y. Okada, T. Maruyama, S. Kaneko, and C. Sasakawa.1994.
PCR-based random amplified polymorphic DNA fingerprinting ofYersinia pseudotuberculosisand its practical applications. J. Clin. Microbiol.32:65–69.
11. Mitsuda, T., H. Kuroki, N. Ishikawa, T. Imagawa, S. Ito, T. Miyamae, M.
Mori, S. Uehara, and S. Yokota.1999. Molecular epidemiological study of
Haemophilus influenzaeserotype b strains obtained from children with men-ingitis in Japan. J. Clin. Microbiol.37:2548–2552.
12. Nakazawa, H., H. Hayashidani, J. Higashi, K. Kaneko, T. Takahashi, and M.
Ogawa.1998. Occurrence ofErysipelothrixspp. in broiler chickens at an
abattoir. J. Food Prot.61:907–909.
13. Nakazawa, H., H. Hayashidani, J. Higashi, K. Kaneko, T. Takahashi, and M.
Ogawa.1998. Occurrence ofErysipelothrixspp. in chicken meat parts from a
processing plant. J. Food Prot.61:1207–1209.
14. Okazaki, M., T. Watanabe, K. Morita, Y. Higurashi, K. Araki, N. Shukuya,
S. Baba, N. Watanabe, T. Egami, N. Furuya, M. Kanamori, S. Shimazaki,
and H. Uchimura.1999. Molecular epidemiological investigation using a
randomly amplified polymorphic DNA assay ofBurkholderia cepaciaisolates from nosocomial outbreaks. J. Clin. Microbiol.37:3809–3814.
15. Popovic, T., C. Kim, J. Reiss, M. Reeves, H. Nakao, and A. Golaz.1999. Use
of molecular subtyping to document long-term persistence of Corynebacte-rium diphtheriaein South Dakota. J. Clin. Microbiol.37:1092–1099.
16. Rasmussen, H. N., J. E. Olsen, and O. F. Rasmussen.1994. RAPD analysis
ofYersinia enterocolitica. Lett. Appl. Microbiol.19:359–362.
17. Shimoji, Y., Y. Mori, K. Hyakutake, T. Sekizaki, and Y. Yokomizo.1998. Use
of an enrichment broth cultivation-PCR combination assay for rapid diag-nosis of swine erysipelas. J. Clin. Microbiol.36:86–89.
18. Shiono, H., H. Hayashidani, K. Kaneko, M. Ogawa, and M. Muramatsu.
1990. Occurrence ofErysipelothrix rhusiopathiaein retail raw pork. J. Food Prot.53:856–858.
19. Takahashi, T., T. Fujisawa, Y. Benno, Y. Tamura, T. Sawada, S. Suzuki, M.
Muramatsu, and T. Mitsuoka.1987.Erysipelothrix tonsillarumsp. nov.
iso-lated from tonsils of apparently healthy pigs. Int. J. Syst. Bacteriol.37:166– 168.
20. Takahashi, T., T. Fujisawa, Y. Tamura, S. Suzuki, M. Muramatsu, T.
Sawada, Y. Benno, and T. Mitsuoka.1992. DNA relatedness among
Erysip-elothrix rhusiopathiaestrains representing all twenty-three serovars and Ery-sipelothrix tonsillarum. Int. J. Syst. Bacteriol.42:469–473.
21. Takahashi, T., K. Zarkasie, S. Mariana, Sumadi, and M. Ogata.1989.
Serological and pathogenic characterization ofErysipelothrix rhusiopathiae
isolates from tonsils of slaughter pigs in Indonesia. Vet. Microbiol.21:165– 175.
22. Takeshi, K., S. Makino, T. Ikeda, N. Takada, A. Nakashiro, K. Nakanishi, K.
Oguma, Y. Katoh, H. Sunagawa, and T. Ohyama.1999. Direct and rapid
detection by PCR ofErysipelothrixsp. DNAs prepared from bacterial strains and animal tissues. J. Clin. Microbiol.37:4093–4098.
23. Tamura, Y., T. Takahashi, K. Zarkasie, M. Nakamura, and H. Yoshimura.
1993. Differentiation ofErysipelothrix rhusiopathiaeandErysipelothrix tonsil-larumby sodium dodecyl sulfate-polyacrylamide gel electrophoresis of cell proteins. Int. J. Syst. Bacteriol.43:111–114.
24. Tilsala-Timisja¨rvi, A., and T. Alatossava.1998. Strain-specific identification
of probioticLactobacillus rhamnosuswith randomly amplified polymorphic DNA-derived PCR primers. Appl. Environ. Microbiol.64:4816–4819.
25. Tynkkynen, S., R. Satokari, M. Saarela, T. Mattila-Sandholm, and M.
Sax-elin.1999. Comparison of ribotyping, randomly amplified polymorphic DNA
analysis, and pulsed-field gel electrophoresis in typing ofLactobacillus rh-amnosusandL. caseistrains. Appl. Environ. Microbiol.65:3908–3914.
26. Warner, J. M., and J. D. Oliver.1998. Randomly amplified polymorphic
DNA analysis of starved and viable but nonculturableVibrio vulnificuscells. Appl. Environ. Microbiol.64:3025–3028.
27. Warner, J. M., and J. D. Oliver.1999. Randomly amplified polymorphic
DNA analysis of clinical and environmental isolates ofVibrio vulnificusand otherVibriospecies. Appl. Environ. Microbiol.65:1141–1144.
28. Welsh, J., and M. McClelland.1990. Fingerprinting genomes using PCR
with arbitrary primers. Nucleic Acids Res.18:7213–7218.
29. White, T. G., and R. D. Shuman.1961. Fermentation reactions of
Erysipe-lothrix rhusiopathiae. J. Bacteriol.82:595–599.
30. Williams, J. G. K., A. R. Kubelik, K. J. Livak, J. A. Rasfalski, and S. V.
Tingey.1990. DNA polymorphisms amplified by arbitrary primers are useful
as genetic markers. Nucleic Acids Res.18:6531–6535.
31. Wood, R. L.1999. Erysipelas, p. 419–430.InB. E. Straw, S. D’Allaire, W. L.
Mengeling, and D. J. Taylor (ed.), Diseases of swine, 8th ed. Iowa State University Press, Ames.