Copyright © 1997, American Society for Microbiology
A Case of Tick-Borne Encephalitis in Japan and Isolation of
the Virus
IKUO TAKASHIMA,1* KOUICHI MORITA,2† MISAKO CHIBA,1DAISUKE HAYASAKA,1
TATSURO SATO,3CHIKAKO TAKEZAWA,3AKIRA IGARASHI,2HIROAKI KARIWA,1
KUMIKO YOSHIMATSU,4JIRO ARIKAWA,4ANDNOBUO HASHIMOTO1
Laboratory of Public Health, Department of Environmental Veterinary Sciences, Graduate School of Veterinary
Medicine,1and Institute for Animal Experiment, School of Medicine,4Hokkaido University, Sapporo,
Department of Virology, Institute of Tropical Medicine, Nagasaki University, Nagasaki,2and Department of
Neurology and Department of Respiratory Disease, Hakodate Medical Association Hospital, Hakodate,3Japan
Received 24 February 1997/Returned for modification 26 March 1997/Accepted 24 April 1997
A case of tick-borne encephalitis (TBE) has not been reported for many years in Japan, although a serological survey of sera from domestic animals suggested the presence of TBE foci in Hokkaido, the northern island of Japan. Studies were conducted to prove the presence of an endemic focus of TBE virus in Japan by means of serology and virus isolation. In October 1993 in Hokkaido, a severe case of encephalitis in a dairy farmer’s wife was diagnosed as TBE. Serological examination of paired serum specimens showed a rise in the neutralization antibody titer to Russian spring summer encephalitis virus. A seroepizootiological survey of dogs showed that the TBE-related virus was prevalent in the area. Three virus isolates were obtained from the blood of sentinel dogs, and antigenic analysis grouped the isolates into TBE-related viruses. Sequence analysis of the envelope protein gene identified one of the isolates as being of the same subtype as the Russian spring summer encephalitis (Far Eastern TBE) virus. The results provide evidence that TBE is endemic in a certain area of Japan.
The family Flaviviridae includes nearly 70 antigenically dis-tinct viruses, many of which cause severe illness in humans and animals (15, 22). Ecologically, they are divided into mosquito-and tick-borne viruses. The latter comprise a complex of vi-ruses including tick-borne encephalitis (TBE), louping ill, Lan-gat, Powassan, Negishi, and Omsk hemorrhagic fever viruses (3, 5), often called the TBE complex of viruses. Like mosquito-borne yellow fever virus, dengue virus, and Japanese enceph-alitis (JE) virus, TBE virus represents one of the flaviviruses that is a major pathogen for humans, causing thousands of cases of severe neurological disease per year. TBE is thus a significant public health problem in areas of Asia and Europe where the organism is endemic. Two subtypes of TBE virus, the Western (central European) subtype and the Far Eastern subtype (the prototype is Russian spring summer encephalitis [RSSE] virus strain Sofjin), are prevalent on the Eurasian continent (4, 9). The latter subtype is known to be highly pathogenic for humans, with a mortality rate of about 30% (17).
In Japan, Negishi virus had been isolated from a patient with encephalitis during the JE epidemic in 1948 in the Tokyo area, and the virus was later identified as a member of the TBE complex by antigenic analysis (1). Since then, no cases of TBE have been reported, nor have further studies of Negishi virus proven the presence of endemic foci or identified the species of the tick vector. After the genomic sequence of Negishi virus was reported recently and compared with those of other TBE complex viruses, the virus showed such a high degree of
ho-mology with louping ill virus that it was judged to be a strain of louping ill virus (21, 23). Since no further isolation of Negishi virus was recorded in Japan and no clear explanation can be given for the introduction of the virus from the British Isles to Japan, the origin of the virus became controversial.
Hokkaido, the northern island of Japan, is located close to the Maritime Province of Siberia in Far Eastern Russia, where TBE is prevalent. Both areas, once connected by land, share the same biogeographical and ecological environments. Ixodes
persulcatus, the tick vector of TBE virus in Siberia, is a
pre-dominant species in Hokkaido, and some rodent species, which are reservoir hosts of the virus, are also common in both areas. From the viewpoint of an ecological setting for the virus, it is not surprising that TBE virus of the Far Eastern subtype is endemic to Hokkaido. The presence of scattered TBE foci in the area has been suspected from our previous seroepidemio-logical surveys in 1978 with cattle sera (18).
In 1993, a case of human viral encephalitis in Kamiiso, the southern part of Hokkaido, was diagnosed as TBE. This report describes the findings for patients with TBE, the results of a seroepidemiological survey, virus isolation, and sequence anal-ysis of the virus envelope (E) protein gene in order to dem-onstrate that TBE is endemic in Japan.
MATERIALS AND METHODS
Cells and viruses.The baby hamster kidney (BHK; ICN Biomedicals Inc., Aurora, Ohio) cell line was grown at 37°C in Eagle’s minimum essential medium (MEM; Nissui Pharmaceutical Co. Ltd. Tokyo, Japan) supplemented with 8% fetal calf serum (FCS; ICN Biomedicals Inc.).
The viruses used were JE virus (strain JaGAr-01) RSSE virus (strain Sofjin; the prototype of the Far Eastern subtype), Langat virus (strain TP-21), Negishi virus, and Apoi virus. The viruses were propagated in 1-day-old suckling mice by intracerebral inoculation, and the brains from moribund mice, after they were killed with an overdose of ether, were homogenized into a 10% suspension in phosphate-buffered saline (PBS; pH 7.2) containing 10% FCS and were frozen at
280°C until they were used. BHK cells were infected with a 10% suckling mouse brain suspension of each virus, and culture fluids were collected on days 3 to 5, frozen at280°C, and used as the working virus stock.
* Corresponding author. Mailing address: Laboratory of Public Health, Department of Environmental Veterinary Sciences, Graduate School of Veterinary Medicine, Hokkaido University, Sapporo 060, Japan. Phone: 81-11-706-5211. Fax: 81-11-706-5211.
† Present address: Regional Office for Western Pacific, World Health Organization, Manila, Philippines.
1943
on May 15, 2020 by guest
http://jcm.asm.org/
Sera and blood.Patient sera were obtained 6 days (acute phase) and 43 days (convalescent phase) after the onset of illness in 1993. Cerebrospinal fluid was obtained on day 52 of the illness (convalescent phase). Sera from other residents in the affected area were obtained in 1994. Blood was obtained in 1994 from nine dogs kept on four farms in the area. In 1995, 10 dogs born in December 1994 were kept on a neighboring farm where the index case of TBE occurred. The dogs were bled once a week from April to July 1995. These dogs were allowed to freely roam the farm to avoid the attacks of predacious brown bears inhabiting the nearby forests and were considered excellent sentinel animals because they were infested with many ticks. The importance of the dogs in the transmission cycle of the virus was not known at the onset of the study. Blood samples were used for antibody examination and/or virus isolation. Blood samples for virus isolation were frozen immediately by placing them in a cooling box with dry ice and were later kept at280°C.
Titration of viruses.Titers of the working virus stock were determined by the focus count method combined with the peroxidase-antiperoxidase (PAP) proce-dure. BHK cell monolayers were formed in 96-well plates (Costar, Cambridge, Mass.) and were inoculated with the serially diluted virus solutions. After 90 min of virus adsorption, the viral inoculum was removed and the cells were incubated in MEM containing 1.5% caboxymethyl cellulose (CMC; Wako Pure Chemicals Ltd., Osaka, Japan) and 2% FCS (CMC-MEM) for 24 h at 37°C. Thereafter, the monolayers were rinsed with PBS and fixed for 20 min with methanol. The plates were air dried and stored at280°C until focus staining.
Foci were visualized by the PAP procedure as described previously (11). Briefly, each well was reacted with mouse ascitic fluids immune to the viruses, followed by reaction with anti-mouse immunoglobulin G (IgG) goat antiserum, mouse PAP (Polysciences Inc., Warrington, Pa.), and substrate solution. The substrate solution was prepared by mixing 0.5 mg of 3,39-diaminobenzidine (Nacalai Tesque Inc., Kyoto, Japan) per ml and 0.05% (vol/vol) hydrogen per-oxide in PBS. Virus foci as brown spots were counted under a dissecting micro-scope with incident illumination. The virus titer of the stock solution was calcu-lated from the virus dilution showing 20 to 60 foci per well.
ELISA.Sera and cerebrospinal fluid were tested for IgG and IgM antibody titers to JE virus by enzyme-linked immunosorbent assay (ELISA). The proce-dures described previously (2) were followed.
NT.Sera were tested for antibody by the neutralization test (NT) by the procedure described previously (11). Serially diluted sera (50ml) were mixed with an equal volume of stock virus (200 focus-forming units/50ml), and the mixture was incubated at 37°C for 90 min. Monolayers of BHK cells grown in 96-well plates were inoculated with the mixtures (50ml/well) and were incubated at 37°C for 90 min. After the virus inocula were removed, CMC-MEM was layered onto the cells, and the mixture was incubated in a CO2incubator at 37°C for 24 h. The monolayers of the inoculated cells in the plate were washed with PBS and fixed with methanol. The focus was visualized by the PAP procedure as described above. The NT titer was determined to be the highest serum dilution that reduced the virus focus counts by at least 80%.
Virus isolation and identification.Dog blood was added to an equal volume of PBS containing 10% FCS, and the mixture was made into a homogenate. The homogenate was centrifuged at 2,0003g for 30 min, and the supernatant was used as the inoculum. One-day-old suckling mice were inoculated with 0.02 ml of the blood samples by the intracerebral route and were observed for 14 days. Dead or moribund mice were removed, and the 10% brain suspensions were prepared as described above. BHK cell monolayers formed in a 50-ml plastic culture bottle were inoculated with the brain suspension and were incubated at 37°C for 72 h. The inoculated monolayers were treated with 0.1% trypsin (Difco Laboratories, Detroit, Mich.) and 0.02% EDTA disodium salt (Wako Pure Chemicals Ltd.) in PBS, dispersed in MEM with 8% FCS, and seeded into 24-well glass slides. After a 4-h incubation in a CO2incubator, the cells were fixed with cold acetone and stored at280°C as antigen slides.
For the serological identification of the virus isolates, an indirect immunoflu-orescent-antibody assay (IFA) was performed with the antigen glass slides pre-pared as described above and monoclonal antibodies provided by Franz X. Heinz of the Institute of Virology, University of Vienna, Vienna, Austria. These mono-clonal antibodies were prepared against the Western subtype of TBE virus, and the reaction patterns to various flaviviruses were described (8, 10, 12). The antigen slides were incubated with monoclonal antibodies for 1 h at 37°C and washed with PBS three times. Fluorescein isothiocyanate-conjugated antibody to mouse IgG (Organon Teknika, Durham, N.C.) was then added to the slides. After incubation for 1 h at 37°C and washing with PBS, the plates were observed under a fluorescence microscope. The IFA titer was determined to be the highest dilution of the antibody showing a positive fluorescent reaction.
Sequence analysis of E-protein gene.RNA was extracted from BHK cells infected with one of the isolates (isolate Oshima 5-10) by the procedure de-scribed previously (11). The RNA was subjected to reverse transcription-PCR targeting for the E-protein gene as described previously (11). Amplified DNA fragments were sequenced by a direct sequencing procedure with a Taq dideoxy terminator cycle sequencing kit and a fluorescence autosequencer (Applied Biosystems, Inc., Foster City, Calif.).
Phylogenetic analysis.The nucleotide sequences of the E-protein genes were analyzed with a genetic analysis software package, Phylip at DNA Data Bank of Japan, National Institute of Genetics, Japan, by the unweighted pair group method with arithmetic means (UPGMA) and the neighbor-joining method. The
reliability of the results was ascertained by majority rule and a strict consensus tree program (copyrighted in 1986 and 1993).
Nucleotide sequence accession number.The nucleotide sequence data for the E-protein gene from isolate Oshima 5-10 reported here will appear in the DDBJ, EMBL, and GeneBank nucleotide sequence databases under accession no. AB001026.
RESULTS
On 27 October 1993, a dairy farmer’s wife in the town of Kamiiso, Japan, suddenly suffered a febrile illness with a tem-perature of 39°C. She was hospitalized on the 3rd day after the onset of illness. Double vision was reported on the 2nd day, and convulsions were reported on the 5th day. She was on a respirator from days 5 to 20. Remaining motor paralysis of the arms and neck was present 3 years later. Cerebrospinal fluid was examined at the acute and convalescent phases, and clin-ical data for the respective phases were cell counts of 468 and 6 cells/mm3, protein concentrations of 167 and 88 mg/dl, and
glucose concentrations of 76 and 69 mg/dl. The results showed that she suffered from encephalitis or encephalomyelitis of viral origin (20).
The dairy farm of the patient was located in the town of Kamiiso in the Oshima District, Hokkaido, Japan (Fig. 1). Four farms were distributed within a 1.0-km radius of that area. The area was deforested 50 years ago. The sera and cerebrospinal fluid were examined by ELISA for JE virus (Ta-ble 1). The cerebrospinal fluid collected during the acute phase was insufficient for serological examination. The ELISA IgG titer for JE virus in sera increased significantly from the acute phase to the convalescent phase. However, the ELISA IgM titer, which is essential for the diagnosis of JE, was negative. Therefore, another flavivirus infection was suspected as the cause of the encephalitis in this patient, and a more specific NT was performed, including NTs for JE virus strain JaGAr-01, Negishi virus isolated in 1948 in Japan (1), Apoi virus isolated from rodents in Hokkaido in 1954 (16), and RSSE virus strain Sofjin. Among the flaviviruses tested, NT antibody titers to RSSE virus increased significantly at the acute and the conva-lescent phases. The results indicated that the patient was in-fected with a TBE-related virus.
[image:2.612.319.557.71.270.2]In 1994, a serological examination of cattle and dog sera kept on farms of a case study area was conducted to confirm
FIG. 1. Geographical location of a case study area.
on May 15, 2020 by guest
http://jcm.asm.org/
that the virus is endemic in the affected area and to examine cows’ milk as a possible source of infection. Positive antibody was not detected in 54 serum samples collected from cows on the farm of the patient and an adjacent farm, suggesting no connection of cows’ milk with the infection source. Dogs in the area were selected as indicator animals on the basis of the observations that dogs were infested with ticks. Sera from nine dogs from four farms in the affected area were collected in July 1994, and NT titers to JE virus, RSSE virus, and Langat virus of the TBE complex were determined (Table 2). Eight of nine dogs showed significantly higher titers to RSSE virus than to JE virus. Although the titer to Langat virus was lower than that to RSSE virus, it was higher than that to JE virus. The results indicated that the TBE-related virus was prevalent in the area. Comparison of NT titers to three flaviviruses demonstrated that Langat virus could be a substitute for the highly patho-genic RSSE virus in the assay.
A serological survey was extended to the residents of the affected area. Five serum samples from humans, including four family members of the patient and a neighboring farmer, were tested. A negative titer (,1:20) to Langat virus was obtained for the four family members. However, the neighboring farm-er’s serum had a specific NT titer to RSSE virus (1:100). Thir-ty-five years earlier he had suffered from severe encephalitis clinically suspected of being caused by JE virus and was hos-pitalized for 2 months. He claimed to have suffered motor paralysis of the neck for 1 year.
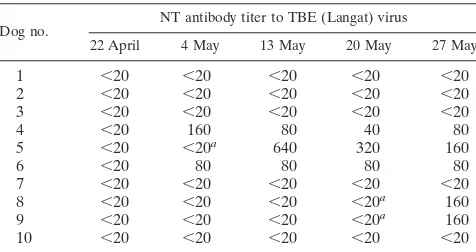
In 1995, 10 sentinel dogs were kept in the area in a free-living environment, and serum samples were collected once a week from April to July. NT titers to Langat virus in the dogs’ sera were determined, and virus isolation was carried out (Ta-ble 3). On 22 April, all 10 dogs were negative for antibody to Langat virus. Two animals converted to positivity on 4 May, three converted to positivity on 13 May, and five converted to positivity on 27 May. The results indicate that the virus was
prevalent in the area in 1995. The blood was inoculated into suckling mice by the intracerebral route. Three viral strains were isolated from the blood of sentinel dogs 5, 8, and 9 which were obtained 7 days prior to the day of seroconversion. The virus strains from dogs 5, 8, and 9 were designated Oshima 3-6, 5-10, and 5-11, respectively. The antigenicities of the virus isolates were examined by IFA with monoclonal antibodies supplied by Franz X. Heinz (8, 10, 12). Three of the isolates showed high titers to TBE complex-specific 2E7 and TBE type-specific 7G7 monoclonal antibodies (Table 4). The viral strains isolated were identified as TBE-related virus.
Sequence and phylogenetic analyses of one of the isolates, isolate Oshima 5-10, was performed, with targeting of the E-protein gene. The whole E-protein gene was amplified by reverse transcription-PCR and was directly sequenced. The sequence of Oshima virus was compared with those of TBE complex viruses (Table 5). The nucleotide sequence identities of Oshima virus with RSSE virus (Far Eastern subtype) and Western tick-borne encephalitis (WTBE) virus were 95.7 and 84.3%, respectively. Oshima virus showed extremely high amino acid sequence identities with RSSE virus (99.0%) and WTBE virus (95.8%).
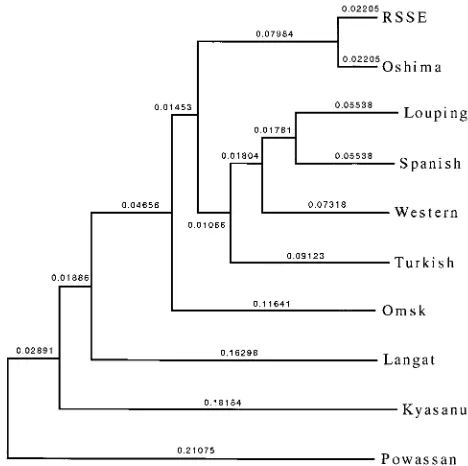
[image:3.612.55.558.81.146.2]Phylogenetic analysis of the E-protein gene was carried out to study the genetic relationship of Oshima virus and other TBE complex viruses. Oshima virus showed the closest rela-tionship to RSSE virus among the other TBE complex viruses tested by the UPGMA method (Fig. 2), and a similar result was also obtained by the neighbor-joining method (data not shown). The high degree of reliability of the tree was ascer-tained by majority rule and a strict consensus tree program; RSSE virus versus the Oshima isolate was 1,000/1,000. TABLE 1. Serological examination of sera and cerebrospinal fluid from a patient with TBEa
Sample (day)
Anti-JE virus antibody titer Anti-flavi NT antibody titer
IgG-ELISA IgM-ELISA JE virus RSSE virus Negishi virus Apoi virus
Serum (6) 1,600 ,100 10 640 40 10
Serum (43) 16,000 ,100 20 2,560 320 10
CSFb(52) 1,600 ,10 NTc NT NT NT
aThe data presented here were published elsewhere (14). bCSF, cerebrospinal fluid.
cNT, not tested.
TABLE 2. Serological examination of dog sera from affected area in 1994
Dog serum
sample no. Farma Age (yr)
NT antibody titer
JE virus RSSE virus Langat virus
1 S 3 80 .1,280 640
2 S 5 20 .1,280 640
3 R 2 40 640 160
4 A 2 10 NTb ,10
5 A 5 80 .1,280 640
6 A 5 10 .1,280 320
7 A 6 ,10 .1,280 80
8 U 2 20 .1,280 .1,280
9 U 10 10 .1,280 320
[image:3.612.317.555.582.708.2]aS, patient’s farm; R, A, and U, adjacent farms. bNT, not tested.
TABLE 3. Monitoring of virus activity and virus isolation by using sentinel dogs in 1995
Dog no. NT antibody titer to TBE (Langat) virus
22 April 4 May 13 May 20 May 27 May
1 ,20 ,20 ,20 ,20 ,20
2 ,20 ,20 ,20 ,20 ,20
3 ,20 ,20 ,20 ,20 ,20
4 ,20 160 80 40 80
5 ,20 ,20a 640 320 160
6 ,20 80 80 80 80
7 ,20 ,20 ,20 ,20 ,20
8 ,20 ,20 ,20 ,20a 160
9 ,20 ,20 ,20 ,20a 160
10 ,20 ,20 ,20 ,20 ,20
aViruses were isolated from the blood of sentinel dogs by inoculation into
suckling mice.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:3.612.59.298.591.709.2]DISCUSSION
This report described an emerging case of TBE and virus isolation from sentinel dogs in Hokkaido, Japan. The case of encephalitis was diagnosed as TBE since in the patient sera the NT titers were higher to TBE virus (RSSE virus) than to other flaviviruses. The NT titers to TBE virus in paired serum spec-imens also increased from the acute phase to the convalescent phase, and the NT titers to JE virus were very low.
It was suspected that the patient acquired the virus infection in the area in Hokkaido, and the virus may have been indige-nous to the area for at least several years. This assumption was made for the following reasons. (i) The patient had no history of travel abroad. (ii) In the years 1994 and 1995, virus activity was clearly demonstrated in the dog population by serological examination and virus isolation. (iii) TBE virus has been shown to be endemic in certain foci for many years due to its ecolog-ical nature. Namely, the virus can be maintained in the tick transstadially and transovarially, and the reservoir animals are indigenous rodents whose migration is very limited.
Dogs were found to be a superior indicator and/or sentinel animals for the TBE virus survey. A high rate of antibody positivity was detected for sera from dogs from the affected area. The time of virus activity could be monitored by testing consecutive serum samples. The viruses were isolated from the blood of sentinel dogs. The viremic period for the dogs was reported to be 7 days before the neutralizing antibodies were first detected in serum from infected animals (7). Although the importance of dogs in the transmission cycle of the virus was not clear, dogs should not be allowed to freely roam the area where the virus is endemic so as to minimize the source of blood meals for ticks, thus reducing the tick vector population. No clinical signs developed in the experimentally infected dogs or in our sentinel dogs (7). Cows, although asymptomatic, have been known to secrete the virus into milk, resulting in out-breaks of TBE due to oral infection (6).
The isolated virus strains were identified as TBE-related
virus from their reaction patterns with monoclonal antibodies, and one of the isolates was shown to be RSSE virus (Far Eastern subtype) by sequence analysis of the E-protein gene. Monoclonal antibody 6E2 reacts widely to flaviviruses, mono-clonal antibody 2E7 reacts to viruses of the TBE complex, and monoclonal antibody 7G7 reacts only to TBE viruses of the Western subtype and the Far Eastern subtype (8, 10, 12). The E-protein gene of Oshima 5-10 showed the closest relationship to RSSE virus. Phylogenetic analysis also demonstrated that Oshima virus was grouped in the RSSE virus Far Eastern subtype. The reliability of the phylogenetic tree was confirmed by several methods. The relatedness among TBE complex vi-ruses found in the phylogenetic tree in this study was very similar to that reported previously (13).
TBE virus may be endemic to areas other than the case study area in Japan. Sera from dogs from another district in Hok-kaido showed a positive antibody response to TBE virus (19).
Ixodes persulcatus ticks were reported to be a principal vector
[image:4.612.57.556.85.152.2]species of RSSE virus in the far eastern part of Russia (6). However, study of our tick collection shows that Ixodes ovatus
[image:4.612.316.550.415.648.2]FIG. 2. Phylogenetic tree of TBE complex viruses. The nucleotide sequence of the E-protein gene of Oshima virus obtained in this study was compared with those of other TBE complex viruses which were available in DNA databases. The tree was obtained by the UPGMA method. The virus strains used for analysis are as follows; RSSE virus (RSSE), Oshima 5-10 virus isolate (Oshima), louping ill virus (Louping), Spanish sheep encephalitis virus (Spanish), Western TBE virus (Western), Turkish sheep encephalitis virus (Turkish), Omsk hemorrhagic fever virus (Omsk), Langat virus (Langat), Kyasanur Forest disease virus (Kyasanur), and Powassan virus (Powassan).
TABLE 4. Identification of virus isolates by IFA with monoclonal antibodies
Specificity (MAb)a
IFA titer to the following virus:
JE Langat Oshima5-10 Oshima 5-11 Oshima 3-6
Group reactive (6E2) .6,400 1,600 .6,400 .6,400 .6,400
TBE complex virus specific (2E7) ,100 .6,400 .6,400 .6,400 .6,400
Type specific (7G7) ,100 ,100 1,600 1,600 1,600
aMAb, monoclonal antibody.
TABLE 5. Identities of the nucleotide and deduced amino acid sequences of Oshima virus with those of TBE complex viruses
Virus % Identity
Nucleotide Amino acid
RSSEa 95.7 99.0
WTBE 84.3 95.8
Louping ill 81.9 91.3
Spanish sheep encephalitis 81.8 92.9
Turkish sheep encephalitis 81.8 3.5
Omsk hemorrhagic fever 80.3 94.0
Langat 75.3 88.7
Kyasanur Forest disease 72.2 81.2
Powassan 69.8 78.1
aFar Eastern TBE virus subtype.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:4.612.58.298.601.718.2]is the predominant species in the case study area and may be the tick vector in this area (19). Since the I. ovatus tick is a common species throughout Japan, epidemiological surveys should also extend to Honshyu, the main island of Japan, to reveal the extent of TBE virus endemicity.
These findings should prompt physicians to consider TBE virus one of the causes of viral encephalitis and to differentiate TBE virus from JE virus in an area such as Japan. JE is prevalent in many Asian countries, but TBE may not have been reported because of clinical symptoms similar to those of JE and the high degree of cross-reactivity of TBE virus and JE virus in serological tests. Prompt preventive measures must be established in Japan by further studies to specify areas where TBE virus is endemic and to develop a vaccine strategy against the prevalent TBE-related virus.
ACKNOWLEDGMENTS
This work was supported by grants-in-aid 07456138 and 08306018 from The Ministry of Education, Science and Culture, a scientific grant from Heiwa Nakajima Foundation, and a scientific grant from the Hokkaido Foundation for The Promotion of Scientific and Industrial Technology.
We thank Franz X. Heinz for providing monoclonal antibodies.
REFERENCES
1. Ando, K., K. Kuratsuka, S. Arima, N. Hironaka, Y. Honda, and K. Ishii. 1952. Studies on the viruses isolated during epidemic of Japanese B enceph-alitis in 1948 in Tokyo area. Kitasato Arch. Exp. Med. 24:49–61. 2. Bundo, K., and A. Igarashi. 1985. Antibody-capture ELISA for detection of
immunoglobulin M antibodies in sera from Japanease encephalitis and den-gue hemorrhagic fever patients. J. Virol. Methods 11:15–22.
3. Calisher, C., H. N. Karabatsos, J. M. Dalrymple, R. E. Shope, J. S.
Poter-field, E. G. Westaway, and W. E. Brandt.1989. Antigenic relationships between flaviviruses as determined by cross-neutralization tests with poly-clonal antisera. J. Gen. Virol. 70:37–43.
4. Clarke, D. H. 1964. Further studies on antigenic relationship among the viruses of the group B tick-borne complex. Bull. W. H. O. 31:45–56. 5. De Madrid, A. T., and J. S. Porterfield. 1974. The flaviviruses (group B
arboviruses): a cross neutralization study. J. Gen. Virol. 23:91–96. 6. Gresikova, M. 1994. Tickborne encephalitis, p. 113–135. In Y. H. Hui (ed.),
Foodborne disease handbook, vol. 2. Marcel Dekker, Inc., New York, N.Y. 7. Gresikova, M., K. Weidneroba, J. Nosek, and J. Rajcani. 1972. Experimental pathogenicity of tick-borne encephalitis virus for dogs. Acta Virol. 16:336– 340.
8. Guirakhoo, F., F. X. Heinz, and C. Kunz. 1989. Epitope model of tick-borne encephalitis virus envelope glycoprotein E: analysis of structural properties, role of carbohydrate side chain, and conformational changes occurring at acid pH. Virology 169:90–99.
9. Heinz, F. X., and C. H. Kunz. 1981. Homogeneity of the structural glyco-protein from European isolates of tick-borne encephalitis virus. Comparison with other flaviviruses. J. Gen. Virol. 57:263–274.
10. Holzmann, H., M. S. Verobyova, I. P. Ladyzhenskaya, E. Ferenczi, M. Kundi,
C. Kunz, and F. X. Heinz.1992. Molecular epidemiology of tick-borne encephalitis virus: cross-protection between European subtypes. Vaccine
10:345–349.
11. Kariwa, H., S. Yoshizumi, J. Arikawa, K. Yoshimatsu, K. Takahashi, I.
Takashima, and N. Hashimoto. 1995. Evidence for the existence of Puumala-related virus among Clethrionomys rufocanus in Hokkaido, Japan. Am. J. Trop. Med. Hyg. 53:222–227.
12. Mandl, C. W., H. Holzmann, C. Kunz, and F. X. Heinz. 1993. Complete genomic sequence of Powassan virus: evaluation of genetic elements in tick-borne versus mosquito-borne flaviviruses. Virology 194:173–184. 13. Marin, M. S., P. M. de A. Zanatto, T. S. Gristsun, and E. A. Gould. 1995.
Phylogeny of TYU, SRE, and CFA virus: different evolutional rates in genus Flavivirus. Virology 206:1133–1139.
14. Morita, K., A. Igarashi, T. Sato, and C. Takezawa. 1994. A case suspected as tick-borne encephalitis in Hokkaido, Japan. Infect. Agents Surv. Rep. 15: 273–274. (In Japanese.)
15. Poterfield, J. S. 1980. Antigenic characteristics and classification of Toga-virideae, p. 13–46. In R. W. Schelesinger (ed.), The togaviruses: biology, structure, replication. Academic Press, Inc., New York, N.Y.
16. Saeki, K. 1961. A neurotropic virus isolated from rodents trapped in the Hidaka area. Nihon Iji Shimpo. No. 1611:1247. (In Japanese.)
17. Shope, R. E. 1980. Medical significance of togaviruses: an overview of dis-eases caused by togaviruses in man and domestic and wild vertebrate ani-mals, p. 47–82. In R. W. Schlesinger (ed.), The togaviruses: biology, struc-ture, replication. Academic Press, Inc., New York, N.Y.
18. Takashima, I., M. Ueda, M. Kiyotake, I. Furuta, and N. Hashimoto. 1992. Epidemiology of tick borne encephalitis. J. Vet. Med. 45:831–835. (In Jap-anese.)
19. Takashima, I. Unpublished data.
20. Takezawa, C., T. Sato, Y. Mizutani, S. Abe, and K. Morita. 1995. Russian spring-summer encephalitis. A case report. Neurol. Med. 43:251–225. (In Japanese.)
21. Venugopal, K., A. Buckley, H. W. Reid, and E. A. Gould. 1992. Nucleotide sequence of the envelope glycoprotein of Negishi virus shows very close homology to Louping ill virus. Virology 190:515–521.
22. Westaway, E. G., M. A. Brinton, S. Y. Gaidamovitch, J. S. Porterfield, P. K.
Russel, and D. W. Trent.1985. Flaviviridae. Intervirology 24:183–192. 23. Zanotto, P. M. D., G. F. Gao, T. Gritsun, M. S. Martin, W. R. Jiang, K.
Venugopal, H. W. Reid, and E. A. Gould.1995. An arbovirus cline across the northern hemisphere. Virology 210:152–159.