A genetic approach to the prediction of drug side effects: bleomycin induces concordant phenotypes in mice of the collaborative cross
Full text
Figure
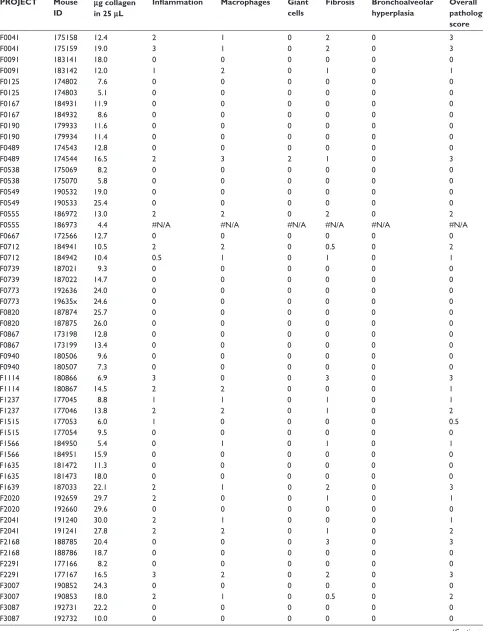
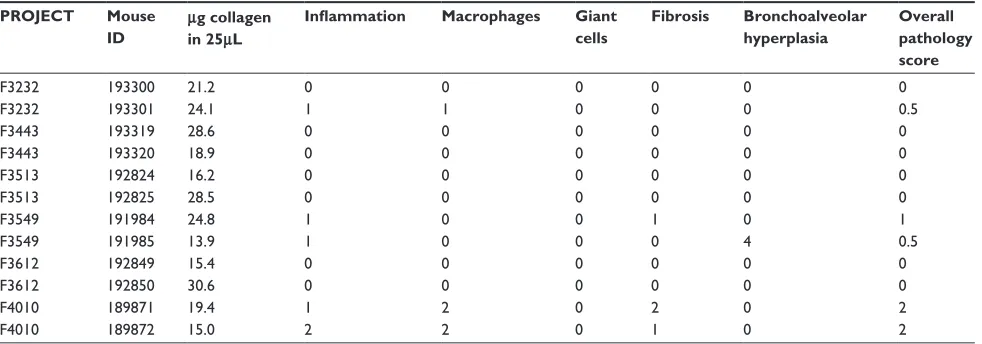
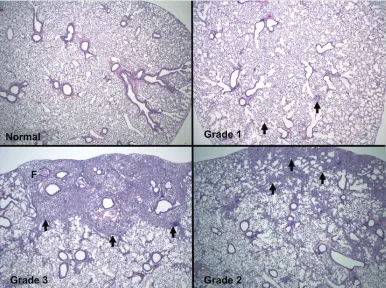
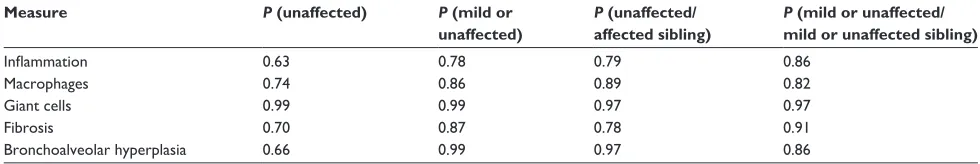
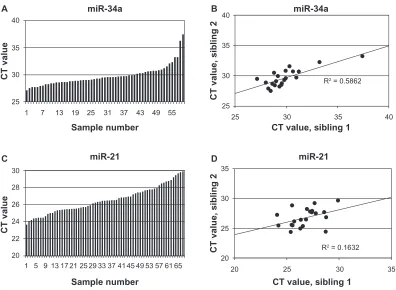
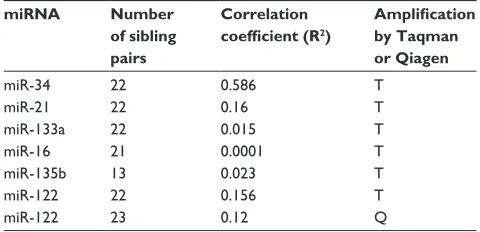
Related documents
The proposed Peyton Slough Hydraulic Relief Project consists of removing an existing hydraulic restriction in Peyton Slough to improve water exchange between McNabney Marsh and Suisun
The instrument contained 13 attitudinal statements related to the newsletter received, measured on a seven-point Likert scale: reactions to the site (newsletter frequency, drop
These included intended purposes of the mission statement, standards for faculty staff, requirements for clinical training by students, program budgetary autonomy and transparency,
infrastructure resources is designed the system composed of a virtual network of virtual machines capable of live migration across multi- domain physical
They would not only become aware of their own inner strengths and characteristics (Seligman et al., 2009), but they would also feel the use of true partnership of the family
The paper assessed the challenges facing the successful operations of Public Procurement Act 2007 and the result showed that the size and complexity of public procurement,
After successfully supporting the development of the wind power technology, an approach is needed to include the owners of wind turbines in the task of realizing other ways, other
