CLONING AND THE CHARACTERIZATION OF THE REGULATORY ELEMENTS OF THE LY-6E.1 (SCA-1) GENE
Angus McLeod Sinclair May 1994
Research Thesis Submitted in Partial Fulfilment of the Requirements of the University of London for
the Degree of Doctor of Philosophy.
Laboratory of Gene Structure and Expression, National Institute for Medical Research,
ProQuest Number: 10106860
All rights reserved
INFORMATION TO ALL USERS
The quality of this reproduction is dependent upon the quality of the copy submitted.
In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uest.
ProQuest 10106860
Published by ProQuest LLC(2016). Copyright of the Dissertation is held by the Author.
All rights reserved.
This work is protected against unauthorized copying under Title 17, United States Code. Microform Edition © ProQuest LLC.
ProQuest LLC
789 East Eisenhower Parkway P.O. Box 1346
To my parents
'Trom the lone sheilding of the misty island,
Mountains divide us, and the waste of the
seas-Yet still the blood is strong, the heart is Highland,
As we in dreams behold the Hebrides. ”
Abstract
Pluripotent haem atopoietic stem cells (HSCs) differentiate into all m a tu re b lo o d cell lin eag es a n d h av e b een c h a ra c te riz e d as the su b p o p u la tio n of m u rin e bone m arro w cells ex p ressin g cell surface m arkers Sca-1+, Thy-1^° and Lin". Stem cell antigen Sca-1, is one of the earliest differentiation m arkers detected on the surface of both fetal liver and ad u lt bone m arrow HSCs. Expression of this antigen also occurs on th y m o c y te p r o g e n ito r s , a c tiv a te d ly m p h o c y te s a n d o n so m e n o n -h aem ato p o ietic tissues. The allo an tig en s d etected by the Sca-1 m onoclonal antibody are encoded by strain specific Ly-6E.l and Ly-6A.2 alleles, m em bers of the Ly-6 m ultigene family. The characterization and isolation of the regulatory elem ents of Ly-6E.l and Ly-6A.2 th at govern tissue specific an d high level expression in the haem atopoietic system , particularly stem cells, are therefore of considerable interest.
Contents Pages
Title 1
Abstract 4
C ontents 6
List of Figures 12
List of Tables 14
A cknow ledgem ents 15
C hapter 1. Introduction 16
1. 1. H aem atopoietic stem cells, Sca-1 and the Ly-6 antigens 18 1. 1. 1. Characterization of the haem atopoietic stem cell (HSC) 18
1. 1. 2. HSC surface antigens 22
1. 1. 3. The Ly-6 alloantigen family 28
1. 1. 4. The Ly-6 A.2 and Ly-6E.l antigens 32
1. 1. 5. Ly-6A.2 and Ly-6E.l antigen function 35
1. 2. Regulation of gene expression 39
1. 2. 1. C hrom atin structure and function 40
1. 2. 1. I. C hrom atin structure: nucleosom es and solenoids 40 1. 2. 1. II. Nucleosom e assembly and transcriptional regulation 40
1. 2. 1. III. N ucleosom al positioning 42
1. 2. 1. IV. Nucleosom es d uring transcriptional elongation 43 1. 2. 1. V. DNase I hypersensitive sites (HS) 44
1. 2. 1. VI. M ARs/SA Rs and m éthylation 47
1. 2. 2. T ranscriptional initiation 49
1. 2. 3. U pstream Prom oter Elements (UPEs) 51
1. 2. 4. Enhancer elem ents 52
1. 2. 5. Locus Control Regions (LCRs) 54
1. 2. 6. Silencers 55
1. 2. 7. Transcription factors 57
1. 2. 8. Interferon action on gene expression 60
1. 2. 8. IL Interferon responsive elem ents and binding factors 1 . 2 . 9. Gene structure and regulatory elem ents of Ly-6A.2 and
Ly-6E.l alleles 1. 3. Project aims
Pâggfi 62
66
69
2. 1. Nucleic acid procedures 71
2. 1. 1. DNA purification 71
2. 1. 1. I. Plasm id m iniprep 71
2. 1. 1. II. Plasm id m axiprep 71
2. 1. 1. in. Cosm id m axiprep 73
2. 1. 1. IV. Genom ic DNA 73
2. 1. 1. V. High m olecular w eight genomic DNA 74
2. 1. 1. VI. DNA fragm ents 75
2. 1. 2. RNA purification 76
2. 1. 2. I. Tissue RNA 76
2. 1. 2. II. C ultured cell RNA 77
2. 1. 2. in. Peripheral blood RNA 77
2. 1. 3. Restriction enzym e digestion 77
2. 1. 4. Electrophoresis 78
2. 1. 4. I. Agarose 78
2. 1. 4. II. D enaturing polyacrylam ide 78
2. 1. 5. Blotting procedures 78
2. 1. 5. I. Colony blots 78
2. 1. 5. II. Slot blot analysis of DNA 79 2. 1. 5. in. Slot blot analysis of RNA 80
2. 1. 5. IV. Southern blot analysis 80
2. 1. 6. H ybridization 81
2. 1. 6. I. Colony hybridization 81
2. 1. 6. IV. Oligo-labelling (random prim ing) 82
2. 1. 7. A utoradiography 82
2. 1. 8. Cosm id library construction and screening 83
2. 1. 9. DNA sequencing 83
2. 1. 10. DNase I hypersensitive site m apping 84
2. 1. ll.P C R am plification 85
2. 1. 11. I. O ligonucleotides 85
2. 1. 11. II. D N A PC R 85
2. 1. 11.in. RT-PCR 86
2. 1. 12. C onstruct form ation 87
2. 1. 12.1. Ligations 87
2. 1. 12. II. C om petent bacteria / transform ation 88
2. 1. 12. III. Bacterial culture 90
2. 1. 12. IV. C onstructs 90
2. 1. 13. SI Nuclease protection analysis 93
2. 1. 13.1. Kinasing of probes 93
2. 1. 13. II. 81 analysis 93
2. 2. Cell culture, transfection and transgenic procedures 94
2. 2. 1. Tissue culture 94
2. 2. 1. I. BW5147 94
2. 2. 1. II. EL4 95
2. 2. 1. in. FDCP-1 95
2. 2. 1. IV. MEL 95
2. 2. 1. V. NIH3T3 95
2. 2. 1. VI. TMIOG 96
2. 2. 1. VII. YAC-1 96
2. 2. 1. VIII. y-Interferon induction 96
2. 2. 1. IX. Cell storage 96
2. 2. 2. Transfection of cultured cells 97
2. 2. 2. I. MEL 97
Faggs
2. 2. 2. in. TMIOG 99
2. 2. 3. Fluorescence-Activated Cell Sorter (FACS) analysis 99
2. 2. 4. H um an grow th horm one assay 100
2. 2. 4. I. Cell preparation for assay 100
2. 2. 4. II. H um an grow th horm one assay 101
2. 2. 5. Transgenic m ouse generation and analysis 101
2. 3. Reagents: Buffers and solutions 102
C hapter 3. C loning and the Characterization of the Ly-6E.l G ene 105
3. 1. In tro d u ctio n 106
3. 2. R esults 106
3. 2. 1. Cosmid cloning of the Ly-6E.l gene 106
3. 2. 2. PCR and sequence analysis of Ly-6E.l 107
3. 2. 3. Restriction analysis of the cosmid clone 111 3. 2. 4. The Ly-6E.l gene is functionally active 114
3. 3. C onclusion 116
C hapter 4. DNase I H ypersensitive Site M apping of the Ly-6A.2 and
Ly-6E.l Genes 118
4. 1. In tro d u ctio n 119
4. 2. R esults 120
4. 2. 1. DNase I hypersensitive sites are present in the 5' and 3'
regions of the endogenous Ly-6A.2 gene in T cells 120 4. 2. 2. Ly-6A.2 DNase I hypersensitive sites are found in
haem atopoietic precursor cells 127
4. 2. 3. DNase I hypersensitive site m apping in Ly-6E.l transfected
MEL cells 130
4. 2. 4. Kinetic induction of Ly-6E.l hypersensitive sites on y-IFN
induction in transfected MEL cells 134
4. 2. 5. Allelic differences between Ly-6E.l and Ly-6A.2 y-IFN
Pages
4. 3. C onclusion 142
Chapter 5. Functional Analysis of Ly-6E.l DNase I Hypersensitive
Sites 146
5. 1. In tro d u ctio n 147
5. 2. R esults 148
5. 2. 1. In vitro deletion analysis of the 5' Ly-6E.l gene
hypersensitive sites 148
5. 2. 2. In vitro deletion analysis of 3' Ly-6E.l gene hypersensitive
sites 152
5. 2. 2. I. Functional analysis of the 3' region from +2.94 kb
to +5.54 kb 152
5. 2. 2. II. Functional analysis of the 3' region from +5.54 kb
to +10 kb 158
5. 2. 3. The Ly-6E.l 3' region, w ith sites HS+8.7 and HS+8.9, contains
a y-IFN responsive elem ent 162
5. 2. 4. N ucleotide sequence and putative factor binding analysis
of sites HS+8.7 and HS+8.9 164
5. 2. 5. In vivo analysis of the 3' Ly-6E.l deletion constructs LGH-7
and LGH-18 166
5. 3. C onclusion 174
Chapter 6. Discussion 180
6. 1. M apping and sequence analysis of the Ly-6E.l gene 181 6. 2. Correlation of DNase I hypersensitive site w ith function and
expression of Ly-6E.l 183
6. 2. I. 5' Regulatory sequences 184
6. 2. II. 3' R egulatory sequences 188
6. 3. In vivo function of 3' regulatory elem ents 192
Pages
List of Figures Pages
Fig 1.1: Schematic representation of spedHc cell surface antigen
expression during murine haematopoiesis 24
Fig 3.1: Comparative Southern blot analysis of cosmid clone cA3.1
and genomic BALB/c (Ly-6E.l) and C57/B110 (Ly-6A.2) DNA 108 Fig 3.2: Sequence analysis of a PCR generated fragment of Ly-6E.l
exon 4 from clone cA3.1 109
Fig 3.3: Restriction map of clone cA3.1 and regions of homology
within the clone 112
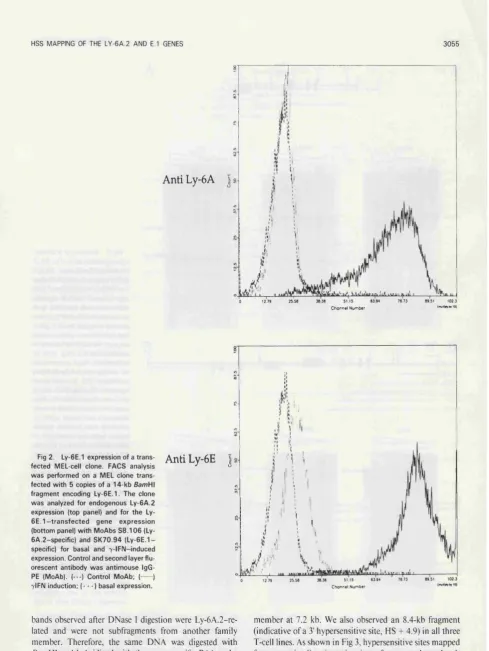
Fig 3.4: FACS analysis of Ly-6E.l and Ly-6A.2 basal and y-IFN induced
expression of a MEL cell clone transfected with the Ly-6E.l gene 113 Fig 3.5: Restriction maps of the Ly-6E.l and Ly-6A.2 alleles 115 Fig 4.1: Mapping of 5' DNase I hypersensitive sites of the Ly-6A.2 allele
A). 3' Probe NBl 121
B). 5' Probe BAl 122
Fig 4.2: Mapping of 3' DNase I hypersensitive sites of the Ly-6A.2 allele
A). 5'ProbeEx4 124
B). 3 'Probe 4.3 125
Fig 4.3: Control mapping of Thy-1 thymocyte specific (intron 3) DNase I
hypersensitive site 126
Fig 4.4: DNase I hypersensitive site mapping of Ly-6A.2 in
multipotential cell line FDCP-1 129
Fig 4.5: Mapping of y-IFN inducible Ly-6E.l hypersensitive sites
A). Probe NBl; B). Probe Ex 4 131
C). Probe 4.3; D). Hypersensitive site map 132 Fig 4.6: Kinetics of Ly-6E.l 5' hypersensitive site induction with y-IFN 135 Fig 4.7: Kinetics of Ly-6E.l 3' hypersensitive site induction with y-IFN 137 Fig 4. 8: y-IFN induced changes within 5' and 3' chromatin of the
Eagea
Fig 4. 9: y-IFN induced changes within 5' and 3' chromatin of the
endogenous Ly-6E.l allele 141
Fig 4 .10: Summary of the 5' and 3' hypersensitive sites of the Ly-6A.2
and Ly-6E.l alleles found in haematopoietic cells 144 Fig 5.1: 5' Ly-6E.l-hCH reporter deletion constructs 150 Fig 5.2: Functional analysis of 5' Ly-6E.l promoter deletions in stably
transfected MEL and TMIOG populations 151
Fig 5.3; 3' Ly-6E.l-hCH reporter deletion constructs 153 Fig 5.4: Functional analysis of the Ly-6E.l 3' region from +2.94 kb to
+5.54 kb in transfected MEL populations 155
Fig 5 .5: Functional analysis of the Ly-6E.l 3' region from +2.94 kb to
+5.54 kb in transfected NIH3T3 populations 156 Fig 5. 6: Functional analysis of the Ly-6E.l 3' region from +2.94 kb to
+10 kb in transfected MEL populations 159
Fig 5. 7: Functional analysis of the Ly-6E.l 3' region from +2.94 kb to
+10 kb in transfected NIH3T3 populations 160
Fig 5. 8: Functional analysis of the Ly-6E.l 3' region containing
hypersensitive sites HS+8.7 and HS+8.9 in transfected MEL cells
and NIH3T3 Hbroblasts 163
Fig 5.9: Sequence and putative factor binding analysis at HS+8.7 and
HS+8.9 165
Fig 5.10: LCH-7 and LGH-18 transgenic expression in the peripheral
blood of founder mice 167
Fig 5.11: Transgene copy number quantitation and analysis for
mosaicism within transgenic founder tissues 169 Fig 5.12: Founder tissue distribution analysis of transgene expression
by SI nuclease protection of RNA 171
Fig 5.13: Summary of the fold increase in expression of the 3' deletion
List of Tables Pages
Table 1.1: Com parison of the tissue distributions of the alloantigens
from the Ly-6 family 30
Table 1. 2: Com parison of the surface expression patterns of
alloantigens Ly-6A.2 and Ly-6E.l 33
Table 5.1: Quantification of tissue specific expression levels of the
Acknowledgements
First and forem ost I w ould like to thank Elaine D zierzak for her supervision, ideas, encouragem ent and enviable enthusiasm , and also for trusting her instincts w hen deciding to take me on as her first student even though Frank and Dimitris hadn't met me ! I am especially grateful to all the members of our group Colin, Bernadette, Hugh, Sharon, Dan (Tracy), Albrecht, David and Sasha for m aking Rm 105 a great and fun (and interesting !) lab to work in. Special thanks m ust go to Bernie for kindly and rapidly sequencing the 3' elements and to Colin for looking after my cells from time to time, taking off blots and for making the transgenics (you can now take over the rest of the room !), and Carol for your sequencing advice and expertise and for being a shoulder to moan to incessantly.
I am especially grateful to all my proof readers and reviewers: Elaine, James Ellis, Ali Immam, Dimitris Kiousiss, Selina Raguz, David Abrahams and M artin Turner. Thank you also Frank Grosveld for putting me through my paces on a M onday morning, Jacky for keeping the departm ent functional and operational, Cora for delivering all my junk mail, Alex for her tissue culture expertise and discussions about holidays, Frank and others in photography for all their work and Chris Atkins for help with the FACS. I m ust also thank John (Stroubie) for letting me finish my thesis before him (Hal), Val for all her laughs, Selina for all her growls. Camélia for our "discrete" conversations, all in GSE and my friends, especially Alice and Cath, not to forget the bakery down Barnet High Street for providing me with my daily cakes !
Chapter 1,
The m aintenance of the haem atopoietic system requires complex signals for the constant renew al of m ature blood cell types during developm ent, and for the extensive proliferation required at times of haematologic stress. All blood cells are derived from single totipotent haem atopoietic stem cells (HSCs), which are thought to reside in the em bryonic yolk sac, fetal liver and finally the adult bone m arrow in mammals (for review: Medvinsky, 1993). The specific cell types, lymphoid (B and T cells) and myeloid (erythrocytes, granulocytes, megakaryocytes, macrophages, eosinophils and mast cells), and the level of o utput of these cells from progenitors, appear to be governed by exogenous signals produced w ithin strom al cell m icroenvironm ent (review: Dexter et al.,
1990). However, the mechanisms of this regulation are poorly understood. The examination of the regulatory mechanisms of haematopoiesis and the factors influencing the decision of HSCs to divide and differentiate, have been the major focus of many investigators and have major implications for cancer, im m unological, tran sp la n ta tio n and gene rep lacem en t therapies.
Haematopoiesis commences at early stages of embryogenesis (7.5 days gestation in the mouse) and continues throughout the lifetime of the adult. This requires the correct tem poral ord er of developm ental sw itching, from the production of embryonic, to fetal and finally adult specific cell types, in addition to switching the sites of haematopoiesis and location of stem cells. The haematopoietic system provides an ideal system in which to examine gene regulation in a developm ental context, in addition to providing an insight into the genetics of stem cells. In an attem pt to further understand gene regulation in HSCs, the regulatory elem ents of an early stem cell m arker have been investigated. The regulatory elem ents of the m urine, strain specific alleles Ly-6A.2 and Ly-6E.l, encoding the surface antigen recognized by the Sca-1 monoclonal antibody, are of particular interest as Sca-1 has been identified as one of the earliest cell surface markers of HSCs (Spangrude et ai, 1988a) .
section describes the attem pts to characterize and purify HSCs by the Sca-1 m onoclonal an tibody and ou r p resen t u n d e rstan d in g of the expression p a ttern an d function of the Ly-6 A.2 and Ly-6E.l antigens. The second review s the general m echanism s that are involved in gene transcriptional regulation, w ith an em phasis on exam ples from w ithin the haem atopoietic system , and prelim inary analysis of the transcriptional regulation of the Ly-6A.2 and Ly-6E.l alleles. The aims section of this thesis is follow ed by the results of m y studies to characterize the regulatory elem ents of the Ly-6A.2 and Ly-6E.l alleles directing expression w ithin the haem atopoietic system .
1. 1. H aem atopoietic stem cells, Sca-1 and the Ly-6 antigens
1. 1. 1. C haracterization of the haem atopoietic stem cell (HSC)
Stem cells are defined as cells possessing high self-renewing capacity w hilst providing the steady state production of differentiated cell types to replace cells w ith a lim ited lifespan. The haem atopoietic system has such pluripotential stem cells which can generate m ature cells of both m yeloid a n d ly m p h o id lineages th ro u g h o u t the lifetim e of an anim al. It is generally accepted th at there is a hierarchy w ith in the haem atopoietic system , in cluding lym phoid and m yeloid p ro g en ito r cells w ith varying d e g re e s of m u ltip o te n tia lity . These cells are fu rth e r a lo n g the differentiation p athw ay than the long-term rep opulating HSCs, and only h av e a lim ited ability to co n trib u te to m u ltilin eag e h aem atopoiesis. Further m ature progenitors can only contribute to few lineages of cells, eg. the granulocyte and m acrophage progenitor (review: Keller, 1992).
As the m ost prim itive HSCs exist at low frequencies, approxim ately one in 10^ -10^ nucleated bone m arrow cells (Jones et al., 1987; Boggs et a l,
ability to lodge in the spleen, pro d u cin g m acroscopic colonies (CFU-S). These colonies, derived from single cells, w ere su b seq u en tly show n to consist of erythroid, m acrophage, granulocyte and m egakaryocyte cells types (Till an d M cCulloch, 1961; Becker et al., 1963; W u et al., 1967). The p lu rip o te n tia l n a tu re of the cells resu ltin g in CFU-S form ation w as exam ined by transplantation of both single CFU-S and cells from colonies from an entire spleen, into secondary irrad iated recipients, resu ltin g in secondary CFU-S (Sim inovitch et al ., 1963, 1964). The ability of prim ary CFU-S to generate CFU-S in double tran sp lan tatio n s w as found to be d e p en d e n t on the tim e of harvesting the original CFU-S. Later colonies (day 14) had an increased ability to form CFU-S w ithin the secondary recipients when compared to the earliest colonies (day 8). This was the first indication th at the cells form ing CFU-S were a heterogeneous population. It is now accepted th at the later CFU-S are derived from m ore prim itive p rogenitor cells (W orton et al., 1969; Magli et fl/.,1982). Increasing evidence, h ow ever, indicated th at the donor cells in CFU-S w ere lim ited in their ability to sustain pluripotential and longterm reconstitution in secondary tran sp lan ts, over extended periods in vivo (H arrison et al., 1978). It was therefore likely that CFU-S were representative of m ore m ature precursor cells and not the long-term reconstituting HSC (Paige et a l, 1979; Kitam ura
et a l, 1981; Magli et al., 1982; Ploem acher and Brons, 1989). Reanalysis of p rim a ry re c ip ie n ts d e m o n s tra te d th a t the m o st p rim itiv e HSC preferentially hom ed to the bone m arrow cavity rath er than the spleen (Barnes et al.,1968; H odgson et al., 1982; van Zant, 1984; Bertoncello et a l,
1985; W olf and Priestley, 1986).
The analysis of haem atopoietic reconstitution in irradiated mice has p ro v id ed a greater u n d erstan d in g of the characteristics of the prim itive HSC, providing a system for the quantitative analysis of prim itive HSCs in ad d itio n to the potential for m utilineage contributions from single HSCs o v er ex ten d ed p erio d s of tim e (Wu et al., 1968; A bram son et al.,1977;
W illiam s et a l, 1984; Dick et a l, 1985; Lem ischka et a l, 1986; Keller and Snodgrass, 1990; Jordan and Lemischka, 1990). This system m easures both the m atu re precursor cells that protect the anim al from radiation induced death, together w ith the short- and long-term repopulating HSCs (Visser et a l, 1984; M üller-Sieburg et al., 1988; Spangrude et al., 1988a). In order to detect the long-term reconstituting HSCs, cells of a m ore m ature phenotype are also transplanted to transiently sustain the m ouse through the initial crisis stage. The ad m in istratio n of p u re HSCs alone is insufficient to p ro tect the anim al from rad iatio n in d u ced death , as the HSCs cannot proliferate and differentiate rapidly enough to su p p o rt the m ouse through th e r a d ia tio n in d u c e d h a e m a to lo g ic c ris is (Jones et al.,1990).
In ord er to quantitate the long-term repopulating HSCs, H arrison (1980) devised a competitive repopulation strategy by transplanting a mix of d o n o r HSCs from mice w ith p a rticu la r h aem oglobin g en o ty p es into irra d ia te d recip ien ts of a n o th er haem o g lo b in g e n o ty p e , alo n g w ith u n m an ip u lated recipient bone m arrow cells. Assays of the haem oglobin type in erythrocytes after extended periods, dem onstrated that the higher the ratio of d o n o r to recipient bone m arrow used the higher the donor haem oglobin type. Further transplantation experim ents to quantitate the lo n g -term re p o p u la tin g cells have been p erfo rm ed in the genetically anem ic m ouse strain W /W ^ mice using lim iting num bers of cells (Russel, 1979). As norm al transplanted m arrow has a selective advantage over the m u tan t m arrow , lethal irradiation is not required and thus relatively small n u m b ers of cells can be assayed. These experim ents have led to the estim ated frequency of long-term repopulating cells as being in the order of one in 3 X 10^ cells in the femur (Boggs et a l, 1982, 1984).
w as unclear w hether HSCs w ere im m ortal or had a finite life span. If im m ortality w as the case, the haem atopoietic system w ould be m ade u p of haem atopoietic cells arising from a lim ited proportion of HSCs, that w ould be active throughout the lifetime. H ow ever, if the life span of HSCs were fin ite, a c o n stan t tu rn o v e r of HSCs w o u ld occur w ith the m atu re h aem ato p o ietic cells arising from different clones, a m odel of clonal succession p ro p o se d by Kay (1965). T herefore ex p erim en ts w hereby individual HSCs could be m arked were desirable .
Early stem cells m arking ex p erim en ts u se d rad ia tio n in d u ced chrom osom al translocations to in d iv id u ally m ark the HSCs (Wu et al., 1968; A b ram so n et a l.,1977). M ore recen tly , rep lica tio n d eficien t retro v iru se s have been used to in d iv id u ally m ark HSCs in vitro th u s allow ing specific retroviral integration p attern s to delineate the progeny from a m arked HSC ( M intz et al, 1984; Keller et a l, 1985; Dick et a l, 1985; Lem ischka et al., 1986). Both m ethods directly dem o n strated th at few plu rip o ten tial HSCs are capable of m ultilineage long-term reconstitution. These experim ents provide a further u n d erstan d in g of the types of m ore m ature progenitors w ithin the haem atopoietic hierarchy.
The retroviral m arking experim ents carried o u t by Lemischka et al.
into a secondary host, indicating that HSCs could return to a quiescent state after replication in vitro (Lemischka et a l, 1986).
Further analysis of anim als over 5 m onths post-transplantation has identified th at single or oligoclonal HSCs predom inantly reconstitute the haem atopoietic system (Jordan and Lemischka, 1990; Keller and Snodgrass, 1990), w ith very little fluctuation in clonal contribution. Initial clonal flu ctu atio n m ay th erefo re re su lt from the infection of b o th m atu re precursors an d transiently functioning prim itive HSCs w hich are active early after transplantation due to haem atopoietic disequilibrium ( Jordan and Lemischka, 1990; Keller, 1992) .
A lth o u g h HSCs can now be m arked and their contribution to specific lineage of the h aem atopoietic system an aly sed , the physical identification and purification of the m ost prim itive HSC has yet to be realized. This is essential for the characterization of HSC properties at a clonogenic level. Techniques to separate HSCs from m ore com m itted precursors (review: Visser and van Bekkun, 1990; Spangrude, 1989) have involved physical properties, including density g rad ien t centrifugation, counter-flow centrifugal élu triatio n and adherence p ro p erties, staining properties w ith a m itochondrial dye (Rhodam ine 123) and a D N A -binding dye (Hoechst 33342), in addition to cell surface antibody and lectin binding affinities.
1. 1. 2. HSC surface antigens
(Berman and Basch, 1985; G oldschneider et a/., 1978) dem onstrated that this a n tig en m ark ed the HSC co m p artm en t. F u rth er en rich m en t stu d ies dem o n strated that cells expressing Thy-1 w ith the absence of m ature lin e a g e m a rk e rs (Lin") for B cells (B220), g ra n u lo c y te s (Gr-1), m yelom onocytic cells (Mac-1) an d T cells (CD4, CD8), en rich ed for p ro g en ito rs of m y elo ery th ro id an d ly m p h o id lineages 50 - 200 fold (Müller-Sieberg et a l, 1986).
M ore recently, this enriched, though heterogeneous subset, w as further subdivided by m onoclonal antibody E13 161-7 (Aihara et a l, 1986). The antibody, recognising an antigen designated stem cell antigen Sca-1, w as found to detect antigens expressed on several subsets of lymphocytes in ad d itio n to p lu rip o ten t HSCs (Figure 1. 1.) (Spangrude et a l , 1988b). T ran sp lan tatio n analysis of cells from bone m arro w selected for the expression of Sca-1, in conjunction w ith Thy-1.ll° and Lin", dem onstrated th a t the T hy-1.ll° Lin" Sca-1 + p o p u latio n rep resen ted the only subset con tain in g HSCs th at could co n trib u te to m u ltilin eag e reco n stitu tio n ( S p a n g r u d e et a l, 1988a; Ikuta et a l, 1990). This p o p u latio n , w hich constitutes 0.05% of the bone m arrow cells, was also found to have the ability of repopulating the thym us, to form late CFU-S, to initiate long-term c u ltu r e s (LTC-IC) (W e ilb a e c h e r et a l , 1991), an d to e n rich for ra d io p ro te c tio n by a factor of 2000 fold, resu ltin g in long term reconstitution of both m yeloid and lym phoid lineages (Smith et al., 1991; S p an g ru d e et a l, 1988a; Li and Johnson, 1992). U chida and W eissm an (1992) also have unequivocally dem onstrated th at this po p u latio n is the only ad u lt bone m arrow population containing pluripotent HSCs.
Figure 1,1;
Schem atic re p re se n ta tio n of specific cell su rface a n tig e n expression d u rin g m urine haem atopoiesis.
ACM
f gUl Liyer
Aorta, gonad mesonephro* derived HSC Sca-17 AA4.17 Embryonic
Thy-1 lo Sca-1-
AA4.1 +
AA4.1 +
Lin +
I
B Linwgt
Precursor Fro B cell Fre B « I I
A dult Bone Marrow
Lymphoid Progenitor
Sca-1 + Thy-1 + AA4.1 + Lin
-Sc»-1 7 Thy-1 + AA4.1 + Lin +
Sca-1 + Thy-1 - AA4.1 + Lin +
M ature
Sca-1 + Thy-1 II
U n - T Lineage
Precursor
Im m ature Thymocyte
Pluti potent HSC
CD4+CD8- Sca-1 + Thy-1 + Sca-1+
Thy-1 lo U n - AA4.1 +
Sca-1 lo Thy-1 + Lin
-Sca-1 - Thy-1 + CD4+CD8+
CD4-CD8+ Sca-1 - Thy-1 +
M ature M yeloid Cells
Sca-1 - Thy1
-Sca-1- Thy-1 - U n +
subsets of Kit expressing and non-expressing cells illustrated that only the Kit+ su b set (70 - 80% of the original p o p u latio n ) form ed CFU-S, and long-term m ultilineage reconstitution was achieved w ith as few as 1 0 - 2 0 injected cells (Ikuta and W eissman, 1992). A lthough Kit is expressed on prim itive HSCs, transplantation experim ents using Sca-1+ and Sca-1" Kit+ L in" bone m arro w cells have conclusively d e m o n stra te d th a t Sca-1 expressing cells are absolutely required for long-term reconstitution (Qkada
et af.,1992). Low levels of CD4, generally a T cell specific m arker, has also been recently found on m urine HSCs (W inem an et aL, 1992). How ever, fu rth er analysis of these cells needs to be carried o u t w ith respect to the levels of detectable expression and sorting param eter stan d ard isatio n as d ata from other groups suggest that lineage m arkers are n o t expressed w ithin the population containing HSCs (Smith et aL, 1991; Spangrude et aL,
1988a; Li and Johnson, 1992).
Further attem pts to purify HSCs has been approached by the low sta in in g of m ito ch o n d rial dye R hodam ine 123, the low sta in in g of D N A -binding dye H oechst 33342, and counter-flow centrifugal élutriation in com bination w ith the exp ressio n of T h y - 1 . L i n " Sca-1+ surface antigens has been carried out (review: Uchida et aL, 1993). These additional m eth o d s have fu rth er d iv id ed the subset into cells th at only p rovide radioprotection and those which provide both radioprotection and contain the HSCs for long-term m arrow reconstitution (Uchida et aL, 1993 and references therein).
HSCs (Ikuta and W eissman, 1992), it has yet to be proven that these are m ark e rs for em b ry o n ic HSCs. A nalysis of lo n g -term re p o p u la tin g em bryonic HSCs from cells w ithin the yolk sac appears to suggest that these cells d o n o t express the Sca-1 an tig en (H uang an d A uerbach, 1993). H ow ever, both em bryonic and fetal HSCs express a m arker recognized by the AA4.1 m onoclonal antibody (Jordan et al., 1990; H uang and Auerbach, 1993). Populations of AA4.1+ cells, in com bination w ith Lin" in fetal liver (Jordan et al., 1990), and w heat germ agglutinin ( W G A h r i g h t ) m arkers in
yolk sac cells (H uang and A uerbach, 1993), contain HSCs capable of m u ltilin eag e co n trib u tio n an d lo n g -term reco n stitu tio n of irra d ia te d re c ip ie n ts. In fu rth e r a tte m p ts to d efin e the e arlie st em b ry o n ic haem atopoietic progenitors, investigators have utilized differentiation of em bryonic stem (ES) cells for the in vitro and in vivo assays of HSCs activity (reviews: M üller and Dzierzak, 1993; Chen, 1992). Even though Sca-1, Thy-1 (Schm itt et al., 1991) and AA4.1 (M üller an d D zierzak, u n p u b lish ed results) were found to be expressed on a low percentage of cells in differentiating ES cystic bodies, it has yet to be proven that these are functional HSCs.
S tra in sp e cific a n tig e n s (a llo a n tig e n s ) su c h as Thy-1 ( M üller-S ieburg et a/.,1986; Jotereau et a l, 1987; Spangrude et a l, 1988a; Spangrude and Brooks, 1992) and Ly-6A.2/Ly-6E.l (Spangrude et a l, 1988a; Li and Johnson, 1992; Jurecic et al, 1993; Spangrude and Brooks, 1993) have been w id ely u tilized in tran sp lan tatio n experim ents to so rt for HSCs. H ow ever, discrepancies exist in defining HSC p o p u latio n s w ith certain a llo a n tig e n s w ith in d iffe re n t m o u se stra in s. The m ajo rity HSC reconstitution experim ents using the Thy-1 antigen as a positive selectable m arker, have utilized HSCs from strains expressing the Thy-1.1 specific allele of the Thy-1 gene (Spangrude et a l, 1988a; 1988c; M üller-Sieburg et
1992). In contrast the Thy-1 alloantigen T cells expression patterns in the tw o strains of mice were found to be identical.
Strain specific p attern s of expression also occur w ith the Ly-6 antigen on HSCs as recognized by the Sca-1 antibody. Serial transplantation and CFU-S analysis using HSCs purified from m ouse strains expressing the Ly-6.1 and Ly-6.2 haplotypes (Spangrude and Brooks, 1993), identified that the HSCs were not confined to positive cell subset from mice of the Ly-6.1 haplotype, b u t were also found in the negative subset. However, the subset containing HSCs from anim als of the Ly-6.2 haplotype was found to be confined p red o m in an tly to the positive p o p u latio n . This im plies th at there m ay be strain specific differences in gene regulation w ithin the HSC com partm ent, a n d /o r strain differences in haem atopoiesis.
Such differences w ithin haem atopoiesis have been studied by van Z ant et al, (1983, 1990, 1992). By aggregating embryos at the eight cell stage, m ice from tw o strain s w ith differing allogenic m arkers (D B A /2 and C57BL/6) w ere created. It was found that HSCs from the D B A /2 strain d o m in a ted the first half of the chim era’s life, an d was su b seq u en tly overtaken by HSCs from the C57BL/6 strain. This dual phase pattern was again seen w hen bone m arrow from the chim eras w as tran sp lan ted into prim ary and secondary hosts. This suggests that there are strain specific program m es of genetic behaviour for HSCs.
Analysis of hum an HSCs has also utilized cell surface expression of hum an antigens. H um an HSCs do not appear to express Thy-1 or a Sca-1 hom ologues, but have been characterized as expressing the antigen CD34 in the absence of CD38 and CD33 expression, and also to possess extensive self renew al properties (Siena et al., 1989; Buhring et a l, 1989; A ndrew s et a l,
potential of the fetal CD34'*' cell subsets (Péault et aL, 1991; Baum et al.,
1992). The pluripotential nature of this cell subset has also been supported by the fin d in g th at CD34+ bone m arro w cells can rep o p u late lethally irrad iated prim ates (Berenson et al., 1988) and on tran sp lan tatio n into preim m une sheep, can lead to long-term m ultilineage contribution to the h aem ato p o ietic system (Srour et al., 1993). H ow ever, it has yet to be elu cid ated w hether the CD34 surface antigen defines the m u rin e HSC co m p artm en t.
1. 1. 3. The Ly-6 alloantigen family
As d escrib e d above, Sca-1 a n tig e n is one of the e arlie st differentiation antigens found to be expressed on HSCs. A ttem pts have been m ade to characterize the Sca-1 antigen and identify the gene which encodes it. Im m unoprécipitation analysis revealed that the surface antigen detected by the Sca-1 m onoclonal antibody also reacted w ith an Ly-6A.2 m onoclonal antibody. In addition, Ly-6A.2 mRNA was detectable only w ithin the Sca-1 + stem cell subset, strongly suggesting that the antigen detected by the Sca-1 antibody was Ly-6 A.2 (van de Rijn et al., 1989). This analysis also dem onstrated that the Sca-1 antibody detected the allelic form of Ly-6A.2, the Ly-6E.l antigen. Since this original analysis, stem cell purification and function has been dem onstrated using the Sca-1 antibody on m ouse strains expressing the alternate alloantigen, Ly-6E.l (Spangrude and Brooks, 1993; Jurecic et al., 1993).
differences in alloantigens w ithin this thesis will be referred to as Ly-6.1 an d Ly-6.2 (Palfree, 1991a), and the in dividual alleles detected by Sca-1 an tibody as Ly-6A.2 and Ly-6E.l, and Ly-6A.2/Ly-6E.l w hen referring to both.
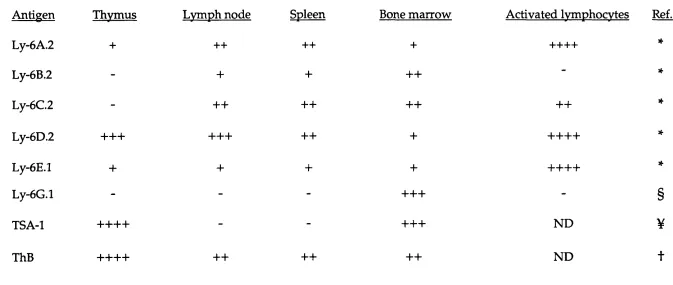
The initial analysis of the antisera id en tified th a t at least six discernible Ly-6 related antigens; Ly-6 A. 2, Ly-6B.2, Ly-6C.2, Ly-6D.2, ThB, an d Ly-6E.l (Kimura et al., 1980, 1984; H ogarth et al., 1984; Yutoku et al.,
1974). Recently, an additional Ly-6 antigen Ly-6G.l has been defined serologically (Flem ing et a l, 1993a). All of the antigens, including the allelic Ly-6A.2 and Ly-6E.l, d em onstrate distinct tissue specific surface expression patterns w ithin the haem atopoietic system (Table 1. 1.).
Initially, it w as u nclear w h eth er the large n u m b er of related a n tig en s w ere d eriv e d from m u ltip le copies of rela te d genes, the differential splicing of a single transcript or w hether a single protein was differentially processed. By using genetic linkage analysis of the Ly-6 antigens, LeClair et al (1987) dem onstrated that there w ere m ultiple genes encoding the Ly-6 antigens and that they occupied a single distinct locus on m ouse chrom osom e 15, w ithin a region of band 15E. Further analysis has show n that the locus is closely linked to the IL-2Rp gene (Malek et al, 1993) and the Pdgf and myc protooncogenes (H uppi et al., 1988; M eruelo et a l,
Tablet. 1;
C om parison of the tissue distributions of the alloantigens from the Ly-6 family.
Table 1 .1.
The surface expression patterns of the family of Ly-6 alloantigens w ithin the haem atopoietic system.
Antigen Thvmus Lvmph node Spleen Bone m arrow Activated Ivmphocvtes Ref.
Ly-6A.2 + 4-4- 4-4- 4- 4-4-4-4- *
Ly-6B.2 - 4- 4- -f-H - *
Ly-6C.2 - 4-4- 4-4- 4-4- 4-4- *
Ly-6D.2 4-4-4- 4-4-4- 4-4- 4- 4-4-4-4- *
Ly-6E.l 4- 4- 4- 4- 4-4-4-4- *
Ly-6G.l - - - 4-4-4- - §
TSA-1 4-4-4-4- - - 4-4-4- ND ¥
cDNAs have also been identified w ithin the rat (Friedman et al., 1990) and dem onstrated substantial predicted protein sequence sim ilarity to Ly-6A.2 and Ly-6C.2. The Ly-6.2 locus has now been m apped and has been show n to exceed 1600 kb, w ith at least eighteen closely linked hom ologous genes localized in three clusters (Kamiura et a l, 1992).
The use of the Ly-6E.l cDNA has also facilitated the cloning of the com plete (Sinclair and Dzierzak, 1993 and herein) and partial (Khan et a l,
1990) Ly-6E.l genomic sequences in addition to the Ly-6 A.2 (McGrew and Rock, 1991a; Stanford et a l , 1992) and the Ly-6C.l genom ic sequence (Bothwell et a l, 1988). Also, the genomic sequences of two new members, Ly-6F.l and Ly-6G.l have been cloned w ith an Ly-6A.2 cDNA probe (Fleming et al, 1993b).
glycosylation has been observed (Rock et al, 1989; Gumley et al, 1992).
The search for the presence of Ly-6 related proteins w ithin other sp ecies h as in d ic a te d th a t the s tru c tu re of th e Ly-6 a n tig e n s is evolutionarily conserved, with Ly-6 hom ologous protein sequences found w ithin h u m an CD59 (Davies et a l, 1989; Stefanova et al, 1989), hum an urokinase-type plasm inogen activator receptor (uPAR) (Palfree, 1991b) and squid neuronal protein gp-2 (Williams et al, 1988), all of which are also GPI lin k e d m olecules. M ore recen tly the Ly-6 m o lecu les hav e been d e m o n s tra te d to hav e p ro te in sequence sim ilarities to n eu ro to x in s (Fleming et a l, 1993b; MacNeil et al, 1993). The hom ology betw een these proteins and Ly-6 antigens is prim arily w ithin the conservation of the 10 cysteine residues. A lthough there ap p ears to be no DNA sequence hom o lo g y of the Ly-6 coding sequences to the genes encoding these stru c tu ra lly related m olecules, it is in terestin g to sp ecu late th at the functions of the Ly-6 proteins are evolutionary conserved.
1. 1. 4. The Ly-6A.2 and Ly-6E.l antigens
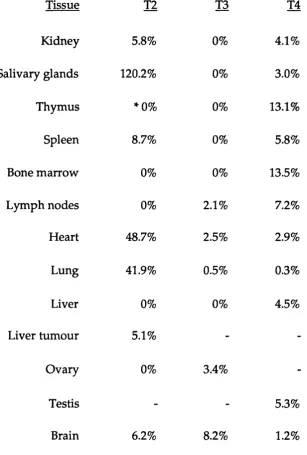
As described above, the Ly-6 A.2 and Ly-6E.l genes have been identified as strain specific alleles of the sam e gene, even though the tissue specific patterns of expression of the alloantigens, differ som ew hat (Table 1. 1.). Initially, it was thought that the Ly-6A.2 and Ly-6E.l molecules were d istin c t m em bers of the Ly-6 fam ily as the tissue d istrib u tio n of the m olecules, determ ined by serological analysis, varied considerably (Kimura
Table 1.2;
C om parison of the surface expression patterns of alloantigens Ly-6A.2 and Ly-6E.l.
D ata is presented as the percentage of cells w ithin the particular tissue that express the antigens, unless stated. Symbols represent the source of the data. Data w as com piled from Kim ura et al, 1984 (*); Codias et a l, 1989 (§); Spangrude and Brooks, 1993 (¥) and Cray et a l,
Table 1.2
Com parison of surface alloantigen Ly-6E.l and Ly-6A.2 expression patterns
Tissue Lv-6E.l LV-6A.2 Ref.
Lym ph node 5% 60-70% *
Spleen 10% 59% §
Bone m arrow 5% 5% *
Activated lymphocytes 80-90% 100% *
Thymus: CD4- CD8- 15% 67% §
CD4+ CD8+ 2% 9% §
CD4+ CD8- 8% 39% §
CD4- CD8+ 30% 21% §
Haematopoietic
stem cells 25% 99% ¥
Brain H ippocam pus
al., 1988). Like the other m em bers of the Ly-6 family, these molecules have a m olecular w eight of 10-12 kDa u n d er non-reducing conditions (Palfree and H am m erling, 1986; Rock et al, 1986) and are also linked to the surface m em brane by a GPI linkage m oiety (LeClair et al, 1986; Su and Bothwell, 1989). Biochemical analysis has in d icated th at Ly-6A.2 possesses no N -lin k ed glycosylation b u t su ggests th at the surface an tig en exhibits O-linked glycosylation (Resier e ta l, 1987).
A lthough the Ly-6E.l and Ly-6A.2 genes are allelic, their antigen expression p attern s w ithin the haem atopoietic system are considerably different in strains of mice from Ly-6.1 an d Ly-6.2 haplotypes, w ith the tissu e an d specific h aem ato p o ietic lineage a llo an tig en d istrib u tio n sum m arized in Table 1. 2. Ly-6A.2 is found to be expressed on a greater n u m b er of a d u lt thym ocytes, p erip h eral lym phocytes and splenocytes com pared to its allelic counterpart, Ly-6E.l (Kimura et al, 1984; Rock et al,
1986; Yeh et al, 1986). Both alloantigens are how ever expressed highly on activated lym phocytes (Kimura et al, 1984). D uring T cell ontogeny, both antigens can be detected on early progenitor double negative (CD4"CD8") thym ocytes, b u t w ith a low er frequency on double positive im m ature thym ocytes (CD4+CD8'''). Subsequently, predom inant expression of both antigens is found on the m ature thymocytes, with the specificity of antigen expression on the CD4+CD8" and CD4“CD8+ subsets dependent on the allele ty p e (C odias et al., 1989). The an tig en s have also been sh o w n to dem onstrate expression anom alies on stem cells (Spangrude and Brooks, 1993). This indicates that complex developm ental control m echanism s are likely to be recru ited in the reg u latio n of both Ly-6A.2 an d Ly-6E.l expression.
brain expression, with Ly-6A.2 lim ited to the vasculature, and Ly-6E.l to the hippocam pal and m idbrain regions (Cray et al, 1990).
Investigations of the cytokines regulating antigen expression has elucidated th at both IFNs y and o/p dram atically upregulate Ly-6E.l and Ly-6A.2 on both T and B cells in vivo and in vitro (D um ont an d Boltz, 1987; D um ont and Coker, 1986; D um ont et al, 1986, 1987; Snapper et al.,
1991a). By in vivo im m u n isatio n it has been d e m o n stra te d th at the augm entation of the surface antigens on B cells is exclusively m ediated by both type I and type II IFNs (Snapper et al, 1991a), w ith an endogenous source of y-IFN required for expression by activated T cells (D um ont and Boltz, 1987). Both types of interferons can act synergistically to further enhance surface expression (Altmeyer et al, 1991). This suggests that these molecules m ay be im portant in im m unoregulation.
A d d itio n al cytokines are able stim u late an d enhance surface expression of the Ly-6 A.2 and Ly-6E.l antigens. T um our necrosis factor (T N F -a) and interleukin 1 (IL-1) increase the induction of Ly-6E.l and Ly-6A.2 e x p re ssio n in vitro ( A l t m e y e r et a l, 1991), h o w ev er the inducibility by IL-1 requires y-IFN to potentiate its effect (LeClair et a l, 1989). A lthough TN F-a alone is ineffective in induction, T N F -a synergizes with y-IFN in vitro to stim ulate Ly-6E.l and Ly-6A.2 expression w ithin bone m arrow and thymocytes, but not B cells (Malek et al, 1989a). However, the synergistic effect of enhancem ent of TN F-a with y-IFN in T cells was only found w ithin strains of mice w ith the Ly-6.1 haplotype, indicating that up reg u latio n of the Ly-6E.l antigen expression, w ith in T cells, is at the tran scrip tio n a l or th e m RNA stab ilisatio n level (M alek et a l, 1989a; A ltm eyer et al., 1991) and is likely to be m ore complex than the regulation of expression of Ly-6A.2 (Malek et al, 1989a).
1. 1. 5. Ly-6A.2 and Ly-6E.l antigen function
developm ental and cellular processes have been investigated b u t are poorly understood and have often been contradictory. N o investigations into the function of the antigens on HSCs have been perform ed. Strong evidence h as been p ro v id ed to suggest an invo lv em en t of Ly-6E.l an d Ly-6A.2 a n tig en s in B and T lym phocytic signal tran sd u ctio n . Studies have dem onstrated th at the binding of specific m onoclonal antibodies to both Ly-6E.l and Ly-6A.2 can induce interleukin 2 (IL-2) production and can resu lt in m atu re T an d B cell activation and proliferation (Codias and Malek, 1990; M alek et al, 1986; Rock et al, 1986; Snapper et al, 1991b). A ntibody binding of the antigens on early fetal thym ocyte progenitor cells has also resulted in cellular proliferation in vitro (Yeh et a l, 1986). The analysis of TCR/CD 3 negative T cell hybridom a m utants by Bamezai et al
(1988), has show n that the TCR/CD3 complex was required for Ly-6A.2 and Ly-6E.l m ed iated activation. This finding has been su p p o rted by the findings of Sussm an et al. (1988) and has been further defined to specifically involve the CD3 Ç chain of the TCR complex and the TCR a /p chains, w hich have been dem onstrated to be absolutely required for the induction of IL-2 production and T cell activation by the antigen specific monoclonal antibodies (Sussm an et al., 1988; W egener et al., 1992). C o-stim ulation of the alloantigens w ith im m obilized anti-CD 3 m onoclonal antibodies has also resulted in the activated phenotype of a T cell hybridom a (Codias et a l,
1990).
O ther GPI linked surface m olecules, such as Thy-1 and Qa-2, can also activate T cells via the cross linking of specific m onoclonal antibodies to these molecules (Kroczek et al, 1986; Stiernberg et a l, 1987) and suggest th a t the activation p h en o ty p e m ay be a general function of antibody crosslinking. H ow ever, experim ents using T cell lines and lym ph node derived cells w ith absent and reduced Ly-6A.2 expression respectively, have d e m o n stra te d th at Ly-6A.2 is ab so lu tely req u ire d for initial an tig en m ediated T cell activation through the TCR (Yeh et a l, 1988; Flood et a l,
ly m p h o k in e s (Flood et al., 1990). The GPI linkage m oiety has been dem onstrated to be necessary for the T cell activation function of Ly-6A.2 and Ly-6E.l (Su et al., 1991). The stim ulatory m echanism crosses species b o u n d aries since the m u rin e alloantigens can function on hum an T cells
(McGrew and Rock, 1991b).
In contrast to these findings, the binding of other distinct antibodies to the Ly-6E.l and Ly-6 A.2 antigens can m ediate a repressive function on T cell activation and expansion in response to m itogenic stim uli (Flood et a l,
1985; Flaque et a l, 1990; Codias et a l, 1990). This suggests that the Ly-6E.l and Ly-6A.2 antigens m ay be bifunctional. Codias et al. (1990) dem onstrated th at the ability of an anti-Ly-6A.2/Ly-6E.l monoclonal antibody to repress T cell activation d ep en d ed on co-stim ulation of the CD3 surface m olecule w ith soluble anti-CD3 m onoclonal antibody. Further studies have clarified th at the binding of the anti-Ly-6A.2/Ly-6E.l monoclonal antibody, resulting in the inhibition of IL-2 production, occurred in the absence of the CD3 Ç chain of the TCR complex (Codias et al., 1992). In the light of the findings by W egener et al. (1992) th at the CD3 Ç chain was necessary for T cell a ctiv atio n via anti-L y-6A .2/L y-6E .l stim u latio n , Ly-6A.2 and Ly-6E.l m olecules m ay perform different functions depending on the presence of specific CD3 chains, and on the types of co-stim ulatory molecules utilized (C o d ias et al., 1990). It is in te re s tin g to s p e c u la te th a t the anti-Ly-6A .2/Ly-6E.l m ediated inhibition of IL-2 production m ay interfere w ith signalling from other CD3 chains (Codias et a l, 1992).
w ith anti-Ly-6A .2/Ly-6E.l, therefore suggesting that factors w ithin m ature T cells, o th er th an the TCR, are req u ired for the suicide response to antibody binding to Ly-6A.2/Ly-6E.l (Nickas et al., 1992). H ow this data fits w ith the T cell activation data described previously is unclear, b u t it may im plicate Ly-6A.2 and Ly-6E.l in the negative selection of autoreactive thymocytes produced during thym us ontogeny (Nickas et al, 1992).
The m ost convincing ev id en ce th a t the Ly-6A.2 an d Ly-6E.l antigens are involved in im m une m odulation has been illustrated by the
in vivo a d m in istra tio n of anti-L y-6E .l an d anti-Ly-6A .2 m onoclonal antibodies to tum our bearing mice. The experim ents perform ed by Lu et al.
(1989) show ed that the adm inistration of the antibodies to the mice resulted in significant tu m our regression an d su p p ressio n of sarcom a, leukem ia an d m elanom a g ro w th , an d this action w as d e p e n d e n t on the Ly-6 h a p lo ty p e of the h o st and n o t the tu m o u r. This has conclusively d e m o n strated th at the antibodies to Ly-6E.l and Ly-6A.2 are able to stim ulate and upregulate host T cell and NK cell an ti-tu m o u r responses (Lu et al, 1989) and supports the in vitro T cell activation data. Significant u p reg u latio n of the alloantigens has also been observed in thym ocytes d u rin g graft-verses-host reactions (Levy et a l, 1988a), again suggesting that the antigens are involved in im m une m odulation. A lthough m onoclonal antibodies have been used in the stim ulation of the T cells via the Ly-6 alloantigens, the natu ral ligands of Ly-6E.l and Ly-6 A.2 have yet to be identified.
transm it signals across the m em brane has been unclear. By tracing the fate of Ly-6A.2 antigens crosslinked w ith monoclonal antibodies, the molecules were found to be internalized by endocytosis, and the internalisation of the com plex form ed only by soluble an tib o d y w as necessary for T cell stim u latio n (Bamezai et ah, 1989). Recently, tyrosine kinases have been fo u n d to co-purify w ith the L y-6A .2/L y-6E .l an tig en s w ith in large, d e te r g e n t re s is ta n t co m p lex es (B o h u slav et al., 1993) a n d by im m u n o p récip itatio n techniques (Stefanovâ et al, 1991). It is likely that these tyrosine kinases, p erh ap s including p56^^^ ( Stefanova et al., 1991; B ohuslav et a l, 1993), are im p o rta n t in the L y-6A .2/L y-6E .l signal transduction pathw ays, resulting in the observed activation of lymphocytes (Stefanova et al, 1991). The precise kinase molecules that are involved in Ly-6A.2/Ly-6E.l m ediated signal transduction require further investigation.
1. 2. Regulation of gene expression
1. 2. 1. C hrom atin structure and function
I. C hrom atin structure: nucleosom es and solenoids
Eukaryotic chrom osomes consist of a single DNA m olecule, which is arran g ed into a higher o rd er stru c tu re by the com plexing of m any proteins to the DNA, to form chrom atin. The com paction of DNA w ithin the nucleus of eukaryotic cells is req u ired as the single DNA m olecule requires som e degree of organisation and condensation for correct cellular function an d for ease of division, en su rin g the inheritance of the full genom ic com plem ent to d au g h ter cells. One of the rep eated u n its of chrom atin is the nucleosome which consists of a basically charged octamer core of histone proteins, com prising two m olecules each of the histones H2A, H2B, H3 an d H4. The octam er core com plexes w ith DNA to accom m odate approxim ately two left-hand twists of 146 bp of B form DNA, to form the characteristic "beads on a string" structure (Klug and Lutter, 1981). On complexing with the core, the helical periodicity of the DNA is changed from 10.5 bp to a non-B DNA form, which contains 10.0 bp per helical tu rn (Levitt, 1978; Klug and Lutter, 1981; Hayes et af.,1991). This D N A /h isto n e octamer complex is further stabilized by the association of an additional molecule, histone H I, w ith the linker DNA. This association is req u ired for the form ation of a higher ord er chrom atin stru ctu re, the 30 n m fibre or "solenoid" w hich consists of six nucleosom es p e r tu rn (McGhee et al, 1980; Felsenfeld and McGhee, 1986).
IL N ucleosom e assem bly and transcriptional regulation
et al., 1988; H an and G runstein, 1988; D urrin et al., 1992). It was found that gene activation d id n o t req u ire the presence of u p stre am activating sequences (UAS) (H an and G runstein, 1988; D urrin et al., 1992). This suggests th at the presence of nucleosom es on the prom oter prevents the form ation of transcriptional initiation com plexes at the TATA box, and th at clearance of chrom atin proteins from the prom oter region requires the binding of fraws-activating factors at UAS (review Svaren and H orz, 1993).
An im portant issue is the ability of transcription factors to recognize and specifically interact w ith their DNA binding sites. This is particularly interesting given that there is an association of the DNA w ith the histone core which occludes one side of the DNA helix, w hilst the other side of the DNA helix is obscured by the presence of an adjacent superhelix (Klug and Lutter, 1981). A dditionally the DNA is further com pacted into a solenoid stru ctu re by histone H I. It appears that the basic N -term inal tails of the histone core proteins m ay have the ability to interact with the linker DNA a n d the D N A su rro u n d in g adjacent nucleosom es, th u s p rev en tin g or h in d e rin g tra n sc rip tio n factors from b in d in g to the p h o sp h o d ie ste r backbone (Hayes et al.,\99V, Lee et al, 1993 ).
regulation of specific genes (W orkman et al, 1988; Alm ouzni et ai, 1990). The histone N -term inal tail residues have been show n to play a role in the repression of gene activation. This effect has been illustrated by the analysis of the repressive action of the N-term inal tail of histone H4 on the m ating loci in yeast (Kayne et al, 1988). A recent investigation by Lee et a l , (1993) on how transcription factors gain access to their b in d in g site, d e m o n s tra te d th a t th e acéty latio n of the N -te rm in a l tails of the nucleosom al core histones released the rep ressio n of X enopus TFIIIA transcription factor binding to the 5S RNA gene. In o rd er to explain the derepression of the chrom atin, it was suggested that the acétylation of the N -term inal tails directly dissociated them from the DNA, a n d /o r that a DN A conform ational change had been induced, allow ing accessibility of the DNA for transcription factor binding. Experiments involving deletions of histone am ino tails from yeast histones H4 and H3, resulting in the d ereg u latio n of gene expression, have also p ro v id ed evidence th at the am ino tails m ay be targets for proteins that regulate chrom atin structure an d may be the sites of action of transcription factors (Durrin et al., 1991; M ann and G runstein, 1992).
IIL Nucleosom al positioning
In contrast to this observation, regulatory proteins th at m aintain DN A nucleosom e free have been found. For exam ple, w ithin yeast it found that the GRF2 factor, which is proposed to bind a specific site w ithin th e G A Ll p ro m o te r (Fedor et al., 1988; Fedor an d K ornberg, 1989), m ain tain ed the p rom oter in a nucleosom e free state so th at additional trans-activating factors were able to bind and facilitate gene transcription (Chasm an et a l, 1990; Brandi and Struhl, 1990).
Recent w ork by Schild et al. (1993) has dem onstrated th at precise nucleosom al positioning in the Xenopus V itallogenin B1 gene can also potentiate transcription by bringing a distal horm onal responsive elem ent in closer proxim ity to proximal elem ents and therefore increasing induced expression by 5-10 fold. A nother positive m odel for the interaction of transcription factor binding sites to m ediate transcription in Drosophila
heat-shock gene hsp26 has been proposed (Thomas and Elgin, 1988; Elgin, 1988).
N ucleosom al positioning can also occur along higher eukaryotic DN A at precise locations (review: Gross and G arrard , 1988). This has d em o n strated to be the case for the p-globin genes in vitro and in vivo,
(Benezra et ah, 1986; Kefalas et al, 1988; Buckle et a l, 1991). It will therefore be interesting to determ ine the function of the positioned nucleosom es on gene regulation in higher eukaryotic organisms.
IV. N ucleosom es d u rin g transcriptional elongation
In an actively transcribing gene, the passage of the RNA polymerase requires that the nucleosom es are disru p ted or displaced in some m anner. H ow ever, this m echanism still rem ains controversial and several m odels have been proposed to explain how nucleosom al disruption occurs during transcription (review: van Holde et al, 1992).
(Clark and Felsenfeld, 1991; 1992). Several questions arise as to how the polym erase disru p ts the nucleosomal structure. One possibility is that the p o ly m e rase b rea k s the h isto n e-D N A contacts as it e n co u n ters the n u cleo so m e a n d causes the d isso c iatio n o r the in sta b ility of the n ucleosom e, resu ltin g in tran sfer (C lark and Felsenfeld, 1992). This m echanism seem s probable as RNA polym erase Il-histone interactions h a v e been re p o rte d by Baer an d R hodes (1983). In a d d itio n , or alternatively, the m echanism m ay be provided by transcriptional-induced supercoiling (Liu and Wang, 1987; Clark and Felsenfeld, 1991).
A m odel for nucleosom e d isru p tio n th at has p rev io u sly been suggested is th at the passing polym erase forces positive supercoils ahead, w hich alter or destabilize nucleosom e cores facilitating their dissociation (Lee an d G a rra rd , 1991). N eg ativ e su p erco ils are left b e h in d the polym erase, to which histone octamers preferentially bind, allowing for the re fo rm a tio n of n ucleosom es b e h in d the p o ly m erase. This m odel rep resen ts the "twin supercoiled dom ain m odel" (W ang, 1987). This m ech an ism w o u ld p ro v id e local assistan ce for tran scrip tio n by the supercoiling generated. The build u p of this torsional stress is likely to be c o n tro lled b y the level of the tran scrip tio n a l activ ity of the gene, distribution of topoisom erase sites and the relative efficiencies w ith which the positive and negative supercoils are relaxed. Evidence for this m odel is also provided by experiments on yeast and E.coli by Wu et al. (1988), Tsao et al. (1989) and Caiver and W ang (1988).
V. DN ase I hypersensitive sites (HS)
references therein). These nuclease sensitive regions, hypersensitive sites (HS), are form ed by changes in the nucleosom al com position of the chrom atin d u e to the association of trans-acting factors or additional protein binding to specific sites w ithin the DNA sequence.
The in crease in se n sitiv ity of n u clease d ig e s tio n of h ig h er eukaryotic chrom atin was initially recognized w ithin the loci of the globin and ovalbum in genes in expressing tissues (W eintraub and G roudine, 1976; Garel and Axel, 1976). An increase in sensitivity of the loci to digestion w ith nucleases was correlated w ith the enhanced transcriptional activity of th e g en es in specific tissu e s. It w as th e re fo re su g g e ste d th a t transcriptionally active genes h ad an altered chrom atin stru ctu re w hich was reflected in the DNase I sensitivity of the gene. The presence of specific regions of ch ro m atin th at contained D N ase I HS w as su b seq u en tly rec o g n ize d w ith in the SV40 an d p o ly o m a o rig in s of re p lic a tio n (Varshavsky et al., 1978; Scott and Wigmore, 1978; Waldeck et al., 1978) and later w ithin cellular chrom atin (Wu et al., 1979).
A DNase I HS w ithin a genetic locus can be constitutive, transiently in d u cib le, tissue specific a n d /o r d ev elo p m en tally reg u la te d , an d is indicative of the c/s-acting genetic control elem ents that are recruited to reg u late gene expression. H ow ever the presence of HSs alone is not sufficient to activate transcription. This was dem onstrated by analysis of the m ouse m etallothionein gene p ro m o ter and Drosophila h eat shock in d u cib le genes. W hile co n stitu tiv e sites w ere fo u n d w ith in these prom oters and were suggested to be indicative of "transcriptionally poised" genes (Senear and Palmiter, 1983; Rougvie and Lis, 1988). Constitutive HSs can also be indicative of silencer elem ents (review: G ross and G arrard, 1988).
The a n a ly sis of th e h u m a n p -g lo b in locus for D N ase I hypersensitivity has identified m ajor regulatory elem ents th at control the high level, tissue and developm ental specific expression of P-globin related genes w ithin a m ultigene locus d u rin g ontogeny. Sites th at dem onstrate sensitivity to DNase I digestion, w ithin erythroid cells, have been identified w ith in the prom oter regions of the transcriptionally active globin genes (McGhee et al., 1981; Stalder et al., 1980). In addition to the sites associated w ith the prom oters and enhancers of the individual genes, one 3' and five 5' sites flanking the locus, are found to be present w ithin erythroid cells an d are developm entally stable d u rin g haem oglobin sw itching (Forrester
et al., 1986; Tuan et al., 1985). These sites are extrem ely nuclease sensitive and the four erythroid specific 5' sites contain elem ents that constitute the locus control region (LCR) of the globin locus. The p-globin LCR has the ability to direct high levels of embryonic, fetal and ad u lt globin expression in the correct developm ental and erythroid specific pattern (review: Dillon a n d G ro sv eld , 1993). M oreover, these su p e rse n sitiv e sites confer dom inant, position in d ependent and copy num ber d ep en d en t expression on the genes in vivo (Grosveld et a l, 1987).
The chrom atin structure of the HS has previously been associated w ith regions of DNA that is "nucleosomal free" (Gross and G arrard, 1988 and references therein). More recent studies have how ever show n, w ithin the MMTV prom oter, th at b in d in g of the fraw s-activating factor to the glucocorticoid receptor elem ent does not displace the nucleosom e in vitro
(Perlm ann and W range, 1988). Further studies of this prom oter in vivo, in ad d itio n to the prom oters of other active genes, has d em onstrated th at histone H I is depleted from the DNase I nuclease sensitive regions of the p ro m o ter, b u t n o t core histones (Bresnick et al., 1992; K am akaka and T hom as, 1990; N acheva et a l, 1989; Postnikov et a l, 1991). H ow ever, studies by W orkm an and Kingston (1992) have dem onstrated th at GAL4 can com pletely dissociate nucleosom e cores from DNA tem plates in vitro.
These results therefore suggest that the D Nase I hypersensitivity of some prom oters and enhancers may be the result of nucleosom e reconfiguration a n d /o r depletion of nucleosom al cores m ediated by the binding of factors (review: A dam s and W orkm an, 1993).
VI. M ARs/SARs and m éth y latio n