0032-0889/91/95/0077/05/$01 .00/0 Accepted August 11, 1990
Similarity between Cytokinin and Blue Light Inhibition of Cucumber Hypocotyl Elongation'
Lila Cohen2, Shimon Gepstein, andBenjaminA.Horwitz*
Department of Biology, Technion-lsrael InstituteofTechnology, Haifa32000, Israel
ABSTRACT
Thecytokinin benzyladenine inhibitedendogenoushypocotyl elongation in intact etiolated seedlings of cucumber(Cucumis sativus L.). In hypocotyl segments, theinhibitory effect of ben- zyladenineongrowthwasclearly detectable inthe presence of indoleacetic acid.Fusicoccin-inducedelongationwasunaffected by the presence ofcytokinin.The effect ofcytokininonelongation of the segmentswasdetermined bymeasuringchanges in fresh weight, a linear function of extensiongrowth. The effect of ben- zyladenineonhypocotyl growthwas atleastaslargein segments prepared from red-light-grownseedlingsasinthose from seed- lings grownintotal darkness. Acomparison wasmade between theinhibitory effects ofcytokinin and bluelight. Theuseof the calcium chelator ethyleneglycol-bis(jB-aminoethyl ether)-N, N'- tetraacetic acid indicatedthatcalcium ionsarerequiredforman- ifestationofbenzyladenine-induced inhibition.
on cucumber cotyledon expansion, and on inhibition of greening, than either inducer alone (4). In thesame system, red lightor cytokininalonecaused maximal acceleration of greening (4). The presence of cytokinin in the incubation medium was found to inhibit auxin-promoted extension growth ofsegmentsinseveral differentspecies (7, 18, 24, 26).
We havestudied theeffects of BA3, asynthetic cytokinin, on stem elongation in dark-grown cucumberseedlings. We investigated whether BA would inhibit IAA-induced exten- siongrowthofhypocotylsegments.Inaddition,wecompared suppression ofgrowth by cytokinin andby blue light. Fur- thermore, weexaminedwhetherthecytokinin-inducedinhi- bition of hypocotyl elongation depends on calcium, as re- ported for inhibitionby bluelight (23).Inall thesestudieswe adopted thetechnicallyeasyandaccuratemethoddeveloped forassaying the effect of IAA on elongation based on cuc- umberhypocotylweightincrease(10).
MATERIALS AND METHODS Thesuggestion thatplant hormonesmightserve assecond
messengersin thesignalresponsecascadeissupported bythe fact that many light-regulated plant growth responses are
reportedeithertoinvolve changesin hormone levelor tobe mimicked byhormoneapplication(for review,see6, 15, 17).
Photocontrol ofstem elongation has been widely explored anddocumented(seerefs. 6, 13).Theeffect ofcytokininson shootelongationis less well known. Photoinhibition ofstem
elongationoperatesthroughatleasttwophotoreceptors,phy- tochromeandcryptochrome(s) (1, 5, 11, 12, 14). Theeffec- tivenessofred andbluelight varies accordingtospeciesand thehistoryoflighttreatments,with,atleast insome species, redlight increasingineffectivenessasseedlingsbecomegreen (6, 13). Cytokininshave been showntoinduce, indarkness, anumberofprocessesnormally controlled by light. Examples include amaranthin synthesis, chloroplast development, and photomorphogenesis (2-4, 25). It is not yet clear to what extentlightandcytokininshareacommon responsepathway.
Inmustardseedlings, kinetin and Pfracted togetherin three differentways toinduce three photoresponses: cotyledon ex- pansion, enzyme induction, and carotenoid accumulation (25).Cytokininand redlight giventogether had a larger effect 'L. C. was supported by a Lady Davis postdoctoral research fellowshipat Technion. B.A. H. holdsfellowships from the Allon Foundationandfromthe Bat-Sheva de Rothschild Fund for Science andTechnology.
2Present address: DepartmentofBotany, George S. Wise Faculty ofLifeSciences,Tel AvivUniversity,RamatAviv 69978, Israel.
Plant Materialand Growth Conditions
Seeds ofcucumber(Cucumis sativus L. cvElem; Hazera, Haifa, Israel)weresowninvermiculitein4 cm x4 cmplastic pots irrigated from below with tap water. The seeds were
germinated for4dat25°Candmaintained either incomplete darknessorunder red light (2 ,umol m-2 s-'). Redlightwas provided by filtering light from cool-white fluorescenttubes throughonelayer ofyellowandonelayerof redacrylic plastic (YavinPlast, Haifa).Thetransmissionofthis combined long- passfilterwas80% above660nm,half-maximalat628nm, andundetectable below 560nm.Fluencerates weremeasured with a quantum photometer (185B; Lambda Instruments, Lincoln,NB). IAAandBAwerefrom Sigma; FC was agift fromE.Marre.
Treatment ofIntact Plants
Variousconcentrations ofBA wereappliedbysprayingthe solution, approximately 15mL/pot,ontotheseedlings. Con- trol pots weresimilarly sprayed withwater.
PreparationandHandling of Segments
Hypocotyl sections approximately 1 cm long were excised using a razor blade. Sections were cut from dark grown seedlings justbelow the hypocotylhook. At 4 d after planting,
3Abbreviations:BA, benzyladenine;FC, fusicoccin.
77
TableI. Hypocotyl Length of Intact Cucumber Seedlings Treated with Various Concentrations of BA
Mean lengths ± SEofthe mean are indicatedforn plants.
BA Hypocotyl Length Inhibition
/AMcm n control
44 11.9±0.45 84 51.0
4.4 19.1 ±0.50 81 21.4
0.44 20.4±0.41 84 16.0
0 24.3±0.82 10
the hypocotyl hooks of red-light grown seedlings had com-
pletely straightened, and sections were cut just below the cotyledons. While preparing sections from dark-grown seed- lings, a green lightata maximum fluencerateof0.35 Mmol m-2 s-' was used for up to 10 min work. For red-grown seedlings, the entire experiment including excision of seg-
ments was performed under the red background light. The sectionswereimmediately placed intotap waterandafterthe desired numberwasobtained,theywere washedin running tap water at 25°C for 30 min. Groups of 10 segmentswere
carefully placed ontoa papertowel, then gently blotted be-
tweentwo pieces of filterpaperbefore being weighedtothe nearestmgusingataredelectronic balance(10). Thesegments were placed in glass scintillation vials containing 3 mL
aqueous medium. Triplicate vials were prepared for each treatment.The data shown in each figure are fromatypical experiment. Means in the figures thus correspond to 30 segments;theerrorbarsareSEof themeanforthe triplicates.
Allexperimentswererepeatedatleast three times.InFigures 3 to 8, growth is expressed as percentage of fresh weight increase, calculatedas
100 x (final weight- initialweight)/(initial weight)
Figure 1. Inhibition of extension growth byvarious concentrations of BA. Seven-day-old dark-growncucumber seedlings were spray- treated with BA after 4 d germination. Concentrations used were
(beginningatleft): 44, 22, 4.4, 2.2, 0.44, 0.22,0.11AMBA,and zero
(control seedlings sprayedwithwater).
30- A
25- -C
0
20+
154-
5
I,1
,,
0-_O
length/I
/
0 / *&
4-,o o
0
fresh weight /
a1, 5--
-IAA -3 -2 -1 0
log [IM, AM]
1
zu I
0
a
0~
C"
4._
-C0
N)U-oL
LLi
15+
l0o
12 16 20 24 28 32
% Length increase
Figure 2. Effect of various concentrations ofIAAonlength and fresh weight of hypocotyl segments. A, relative growth, definedas(final- initial)/(initial), measured by each method. B, Linear dependenceof fresh weight increaseonlength increase, plotted from the datain(A).
Growthwasmeasured after 3 h incubation.
Thegrowthratevaried betweenindependent experimentson
differentsetsofplants (compare growthof controls inFigs. 3, 7,and8). Therefore,whenattestwasused tocomparemeans
for different treatments,thegrowth differential,definedas
(% fresh weightincrease without BA)
- (% fresh weightincrease withBA)
wascomparedbetweenexperiments.
Treatment ofSegments
Astandard mediumcontaining2 mm KCI,0.1 mM CaCl2
wasused,exceptinexperimentswithEGTA,in whichcase a
B .
.
:) i I i It
'I,
A.9...
_6%
4) U)
a
4,
U)
-4,
T
26- 0
9 X0
24-X
220- /
0
20 //
-BA -3 -2 -1 0 log [BA, 4M]
50
4)
U) 0 4,
c
-C 0)1
UL) I,
40-
30+
20-
10-
0
Figure 3. Inhibition by BA of IAA-promoted growth of hypocotyl segments.Segmentswereincubated for 3 h with 57AiMIAA.Assay
wasby fresh weight determinations;errorbars,shown wherelarger than the symbols,are SEof themeanfor thetriplicates.
medium containing5 mm K phosphate (pH 6.0)was used.
IAA,BA, FC, EGTA, andCaCl2wereaddedtothe medium where indicated in the different experiments before the seg- mentswereintroduced into the vials. The vialswere placed inacoveredreciprocalwaterbath shaker(70strokes/min)at 25°Cfor 3 h, afterwhich time the sectionswereremoved from themedium,blottedgentlyasdescribedabove,andweighed.
Intheblue light experiments,the vialswereincubatedat25°C with gentle shaking in a controlled temperature darkroom wherearedlight backgroundwasmaintainedduringthe 3-h bluelighttreatment. Blue irradiation(30 ,gmol m-2 s-')was
produced by a projector equipped with a 150 W tungsten halogen lamp, heat filters (coating number 116 [Schott, Mainz, Federal Republic ofGermany], 10 cm of distilled water,wide band hot mirror[Optical Coating Lab, Inc.,Santa Rosa, CA]), and a dichroic blue filter. The transmission of this filter combination is less than 1% below 395 nm and above 530nm,andmorethan 60% between410 and 505nm.
/'
O control
X 44 uM BA ,\
U)
30 C:
0'20
10- I T// I I I
- IAA 0.1 1.0
IAA, AM
10 100
Figure 4. Inhibition of IAA-promoted growth by BA,as afunctionof the IAA concentration. Assay was byfresh weight determinations, afteragrowth periodof 3 h inthepresenceof57 AM IAA.
noFC 0.1 1.0
FC concentration (A.tM)
Figure 5. Failure ofBAtoinhibit thegrowth promoted byFC.Assay conditionswereasforFigures 3 and 4, exceptthat IAAwasabsent from the medium and growth of the segmentswasinducedby FC.
RESULTS AND DISCUSSION
Table I shows the hypocotyl length of intact 7-d-old dark
growncucumberseedlings thatweretreated 72 h earlier with various concentrations of the synthetic cytokinin BA. In addition to inhibiting hypocotyl elongation strongly, appli- cation ofhigh concentrations of BA(22to44 fLM) produced hypocotyl thickeningandcotyledon expansion (Fig. 1). Ten millimeterlongsections ofhypocotyl,excisedjustbelow the hypocotyl hook of dark-grown seedlingsand incubated in a
simple medium containing different concentrations ofBA, failedtorevealanyinhibitoryeffect ofcytokininonhypocotyl growth (datanot shown). Whether growthwas measured as
increaseinlengthorfreshweight,about 6,uMIAAwasoptimal forpromoting growthof thehypocotylsegments inthis cul- tivar (Fig. 2A). The fresh weight increase showed a linear dependence onlength increase (Fig. 2B). With the addition of IAAtothemedium,it becamepossibletodetect,after3 h incubation, an inhibition ofgrowth by BA. The inhibition increased withBAconcentrationupto44,M (Fig. 3).At IAA concentrations iround theoptimum, an inhibitoryeffect of 44,uM BAwasobserved(Fig. 4).An IAA concentration of 57 ,uM waschosen forsubsequent experiments.
Todeterminewhether theeffect ofBA onhypocotyl elon- gationwasspecificforgrowth inducedby auxin, growthwas
promoted by the fungal toxin fusicoccin (22). FC, likeIAA,
TableII. Comparisonof theEffect ofBAonGrowth of Hypocotyl SegmentsofDark-Grown and Red-Light Grown Seedlings
Means and SE of the mean are indicated for six replicates, 10 segmentseach, fromtwoexperiments.
Treatment Growth Rate Inhibition 44 MmBA mgh-1
control
Dark + 7.78± 1.4 28.4
- 10.87±0.4
Redlight + 1.72±0.06 39.0
- 2.82±0.16
I= control 44AMBA
+Ig + g~~
,, ,,.Iiz,,j
1
12-
En 10-
U
1)
0 8-
E- L-Q)
2L-
0
-BLUE +BLUE
@303 0
-c
.5
c-(n
@3
Figure 6. Comparisonof blue light and BA inhibition of IAA-promoted growth of hypocotyl segmentsfromred-lightgrownseedlings. The entireexperimentwasdoneinthepresenceofcontinuous red light (2 ,molm-2s-1). Blue light(30 ,molm-2s-1) and/or BA (44AM)were
applied for the entire 3-h incubation period, afterwhich growthwas
measured by fresh weight determinations. The IAA concentration
was57AM.
31) U)a
31)
C:
c
c-
U) 31) LL.
40(-5--- T
- -.-
I- 0 35
.~~~~~~~~ \
0' 30
_1
25 i
O control, 57u.MI'M
* +44 MBA 20
0 0.01 0.1 1.0 EGTA (mM)
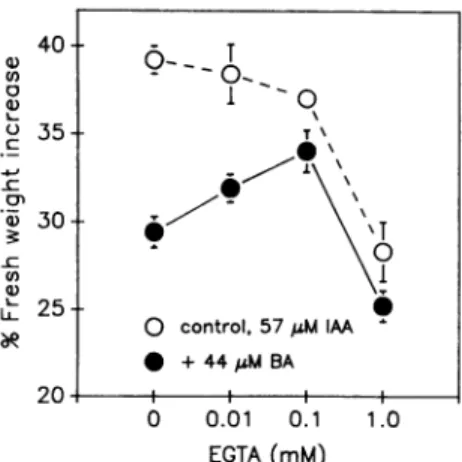
Figure7. Effectof EGTAontheabilityof BA to inhibitIAA-promoted growth.Increasingconcentrations of EGTAwereusedinaCa2l-free
incubation medium(see "Materials and Methods"). Growth of seg- ments from dark-grown seedlings was assayed by fresh weight increase, after3 h incubation in thepresenceof 57 MmIAA. Atypical experimentisshown; symbolsindicate theaveragegrowthof three sets of 10 segments; error bars, shown where larger than the symbols, are SE of the mean for the three replicates. The mean
growthdifferential inducedby BA, defined as(percentfreshweight increase withoutBA)minus(percentfreshweightincreasewithBA)
was:8.64±0.75%(SEofthemeanfor fiveindependentexperiments) withoutEGTA; 1.32 ±0.66%(seven experiments)withEGTA. The effect of 0.1 mm EGTA is significant ata probability level P =2.6 10-5(ttest).
36 ~~~~~0--C 57suMIAAand0.1 mMEGTA
34 6A *- +44/M BA
32 T
T
+BAJ \\
24 I0~~~~~~~~~~~~~
20~~~~~~~~
18~~~~~~~~~~~~~~~~~~
-EGTA 0 0.1 1.0
CoCO2 (mM) 10
Figure 8. Reversal of theeffect of EGTA chelation by addition of Ca2+. Bargraph (left) illustratestheeffect of BA in the absence of EGTA. Forthecurves(right), theCa2+concentrationwasincreased
asindicated,in thepresenceof0.1 mmEGTA.Otherconditionswere asforFigure7. Atypical experiment is shown:meanandSEofthe
mean areforthree replicates of10segmentseach.Themeangrowth differentialsattributed to BAwere4.73± 0.66%and5.6± 1.4%at 0.1 and 1.0mmCa2+, respectively (three independent experiments each). By comparison (t test)with the meangrowth differentialfor 0.1 mM EGTA withnoaddedCa2+(seeFig. 7),the reversal byCa2+
is significantatP=0.015andP=0.012,respectively.
induces cellelongation, butmaydosobyadifferent mecha- nism (19, 24). FC promoted rapid growth (Fig. 5). In the
presence of FC, BA failedto inhibit growth of the cucumber hypocotylsegments(Fig. 5),as wasshown also forsegments ofpea epicotyl (24), revealing the selective nature of the inhibitory effect of cytokininoncellexpansion.
Since totally dark-grown seedlings can alter their growth rateinresponsetoextremely smallamountsof light (very low fluence range; ref. 21), the question naturally arose as to whether preparingsegmentsfrom dark-grown cucumber seed- lings under green light and subsequently exposing them to dimlaboratory fluorescent light from the washstageon,might override some of the inhibitory effects of BA. In order to separate the effects ofBA onthe growth of segments from
any phytochromeresponse, cucumberseedlings weregrown
under a constant low level of red light, so as to keep the photostationary level ofPfratahighandnearlyconstantlevel (16). This strategy has also been used to separate crypto- chromefromphytochromeeffects in the measurement ofblue light suppression of growth (20, 23, 27). Under continuous red light, the cotyledons were larger and partially greened, and thehypocotylswereshorterthan those of etiolated seed- lings; such seedlings are thus often referred to as "de-etio- lated." Although growth was slower, the relative inhibitory effect ofBA was at leastas large when hypocotyl segments
wereprepared from de-etiolatedseedlingsaswhentheywere cut from dark-grown seedlings (Table II; Fig. 6). Red-light
grown seedlingswere usedtocomparethe inhibitory effects ofBA (44 ,M)and blue lighton hypocotyl growth. BAand bluelightinducedinhibitoryeffectsonhypocotyl growththat weresimilarinmagnitude(Fig. 6). Furthermore,the inhibi- tory effects were not enhanced when BA and blue light inductionswerecarriedoutsimultaneously.
= control, 57 ,uM IM IE +44MMBA
r/f/I 1,rfsSta
I
I
I
2-1
d
I
I
d d 1
Blue-light inhibition of hypocotyl segment elongation showed a specific requirement for Ca" (23). Furthermore, calmodulin-binding compounds have been found to inhibit several cytokinin-regulatedprocesses(8, 9). An obviousques-
tiontoask, therefore,waswhetherornotCa2+ is required in order for BA to inhibit hypocotyl growth. Addition of the specific Ca" chelator EGTA to the incubation medium (5
mMKphosphate,see"Materialsand Methods") showed that Ca>2 was also required for BA-induced growth inhibition.
EGTAataconcentration of 0.1 mM diminished the capacity of BAtoinhibit theIAA-promoted growth ofsegments(Fig.
7). EGTA at 0.1 mm stimulated the growth of BA-treated segments, while atthe sametime thegrowth of control (no BA) sections decreased slightly. The highest concentration of EGTA, 1.0mm,wasinhibitory for growth of both BA-treated and nontreated segments(Fig. 7). Addition of CaCl2 tothe incubation medium containing 0.1 mM EGTA partially re-
versed the effect of the chelator: segmentsincubated with 0.1
or 1.0mM CaCl2 exhibited aninhibitoryresponsetoBA. At
10mmCaCl2 and 0.1 mmEGTA, the growth of both control and BA-treatedsegmentswasinhibited, and the effect ofBA
was nolongerapparent(Fig. 8).
Blue light and cytokinin elicited similar inhibitory growth
responses. Ca>, perhaps in the cell wall (23), may act as a
signal transmitter in both inhibitory effects. Although the mechanism of cell elongation is not completely understood, themimickry of blue-light inhibition by cytokinin raises the possibility that this hormoneactsas amessenger ortransducer for blue light, though the fastest blue light responses leave little time forasignaling moleculeto act(5, 6). Alternatively, cytokinin and blue lightmayindependentlydecrease therate of extensiongrowth.
LITERATURE CITED
1. Attridge TH,BlackM,GabaV(1984)Photocontrol ofhypocotyl elongation in light-grown Cucumis sativus L. A synergism between theblue-lightreceptorandphytochrome.Planta 162:
422-426
2. Bianco-Colomas J(1984) Effectofcytokinin antagoniston cy-
tokinin and light-dependent amaranthin synthesis inAmar- anthus tricolorseedlings.PlantGrowthRegul2:281-287 3. Bianco-ColomasJ,Peaud-LenoelC,Bulard C (1988) Use of 5,6-
dichloro-l-b-D-ribofuranosylbenzimidazoletodistinguish light stimulation fromcytokininstimulation of amaranthinsynthe- sis in Amaranthus tricolor. PlantGrowthRegul7: 19-27 4. CohenL,ArzeeT,Zilberstein A(1988) Mimicry by cytokininof
phytochrome-regulatedinhibitionof chloroplast development inetiolated cucumber cotyledons. Physiol Plant 72: 57-64 5. CosgroveD(1981) Rapid suppression ofgrowth byblue light.
PlantPhysiol 67: 584-590
6. CosgroveDJ (1986)Photomodulation ofgrowth.In RE Ken-
drick, GHM Kronenberg, eds, Photomorphogenesis in Plants.
MartinusNijhoff/Junk, Dordrecht, Netherlands, pp 159-183 7. De Ropp R (1956) Kinetin andauxin activity. Plant Physiol 31:
253-254
8. Elliot DC(1983) Inhibition ofcytokinin-regulated responses by calmodulin-binding compounds. Plant Physiol 72: 215-218 9. Elliot DC, Batchelor SM,Cassar RA, Marinos NG (1983) Cal-
modulin-binding drugs affect responses to cytokinin, auxin andgibberellic acid. PlantPhysiol 72: 219-224
10. Epel BL, Erlanger M, Yahalom A (1987) The etiolated cucumber hypocotyl weight-growth test. A sensitive, easy and ultrafast bioassay for IAA. Plant Growth Regul 5: 3-14
11. Gaba V, Black M (1983) Photocontrol of hypocotyl elongation indeetiolated Cucumis sativus L. Rapid responsestobluelight.
PhotochemPhotobiol 38: 469-472
12. Gaba V, Black M(1985) Photocontrol of hypocotyl elongation in light-grown Cucumis sativus L. Planta 164: 264-271 13. Gaba V, Black M (1983) Thecontrolof cell growth by light. In
Shropshire Jr W, Mohr H, eds, Encyclopedia of Plant Physi- ology.Springer-Verlag,Berlin-Heidelberg, pp 358-400 14. Gaba V, Black M (1979) Two separate photoreceptors control
hypocotyl growthin greenseedlings.Nature278: 51-54 15. Horwitz BA(1989) The potentialforsecond messengers inlight
signalling.In WFBoss, DJ Morre, eds, Second Messengers in PlantGrowth and Development. Alan R.Liss,New York, pp 289-313
16. Iino M, Briggs WR (1984) Growth distribution during first positive phototropiccurvatureofmaizecoleoptiles.Plant Cell Environ7:97-104
17. JohnsonCB (1982)Photomorphogenesis.In HSmith,DGrier- son,eds, The MolecularBiologyof PlantDevelopment.Black- wellScientific Publishers,Oxford/London, pp 365-404 18. Katsumi M (1963)Physiologicaleffectsofkinetin.PhysiolPlant
16: 67-72
19. Kutschera U, SchopferP(1985) Evidence for the acid-growth theoryof fusicoccinaction. Planta 163: 494-499
20. Laskowski MJ, Briggs WR (1989) Regulation ofpeaepicotyl elongationby bluelight.PlantPhysiol89:293-298
21. MandoliDF, Briggs WR(1981) Phytochrome controloftwolow irradiance responsesin etiolated oat seedlings. Plant Physiol 67: 733-739
22. Marre E(1979) Fusicoccin. A tool inplant physiology. Annu RevPlantPhysiol30: 273-288
23. Shinkle JR, Jones RL(1988) Inhibition ofstem elongationin Cucumisseedlingsby bluelightrequirescalcium. PlantPhysiol 86:960-966
24. TheologisA (1987) Possible link between auxin regulated gene expression,H+secretion,and cellelongation.Ahypothesis.In D Cosgrove, D Knievel, eds, Physiology of Cell Expansion duringPlantGrowth.AmericanSocietyof Plant Physiologists, Rockville, MD, pp 133-144
25. Tong A, Kasemir H, Mohr H (1983) Coaction of light and cytokinininphotomorphogenesis.Planta 159: 136-142 26. Vanderhoef L, Stahl C, Siegel N, Zeigler R(1973)Theinhibition
bycytokinin of auxin-promoted elongationinexcised soybean hypocotyls.Physiol Plant 29: 22-27
27. Warpeha KMF, Kaufman LS (1989) Blue light regulation of epicotylelongationin Pisum sativum. PlantPhysiol 89: 544- 548