J. Exp. Biol. (1963), 40, 393-401 2 9 3 With 3 text-figura
Printed in Great Britain
DELAYED RESPONSES IN OCTOPUS
BY P. N. DILLY
Department of Anatomy, University College London
(Received 11 February 1963)
INTRODUCTION
Since Hunter's (1912) original experiment with rats, it has been shown that many animals can learn to choose correctly between identical shapes even though the signal indicating the correct choice is removed before the animal is allowed to make the choice. The period during which an animal can be kept waiting after the signal has been removed and still make the correct choice varies from species to species and with the conditions (Konorski, 1959). It is difficult to judge the significance of these differences since the experimental situation is usually different in each species tested. The times vary from 5 to 10 sec. in rats to several hours in some primates.
The cephalopods possess relatively and absolutely larger brains than any other invertebrate, and many experiments have demonstrated the ability of Octopus to learn to attack one of a pair of shapes and to avoid the other.
The only experiments that have so far considered the capacity of the octopus to make a delayed response are those of Schiller (1949). He concluded that correct per-formance was correlated with the maintenance of bodily orientation during the time for which the animal 'remembers' the correct direction.
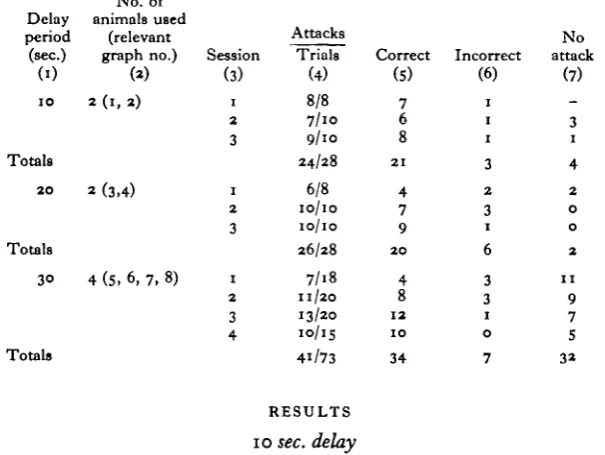
In the present experiment the animals were shown two identical shapes, one of which had been accompanied by a crab. The crab was then removed and after a fixed time the octopus was allowed to attack the shapes. The situation allowed for a dis-turbance to be introduced during the delay period, in order to disturb the 'bodily orientation'.
METHODS
Octopuses were isolated in rectangular tanks, 100 x 40 x 40 cm., with opaque sides and lids. They were given bricks arranged as a home in the middle of one end of the tank. The apparatus used is a modification of that used by Sutherland and Muntz (1959) for their simultaneous discrimination experiments. An opaque screen with a large rectangular aperture in the middle of its bottom half was fixed one-third of the way along the tank, in front of the home. The aperture in the screen was fitted with slides so that a transparent door could be placed across it. The lid of the tank consisted of two parts, one (outer) part could be removed completely, while the remaining part covered the home and extended as far forwards as the screen.
At the end of the tank opposite to the home two identical white plastic rectangles were fixed with their long axes vertical against a black matt screen. They were separ-ated from one another by a black matt partition extending lengthways along the tank and large enough to prevent the octopus from reaching both shapes simultaneously.
394
-The screen, partition and shapes were all stuck together and moved as a unit from tank to tank during the testing of a series of animals.
In order to obscure the shapes from the animal during the delay period an opaque black plastic mask, larger than the aperture in the screen, was used. This mask was mounted firmly on a handle, and could be pushed rapidly about the tank (Fig. i).
Octopuses that had previously been rewarded for attacks on crabs or vertical rectangles were used. The outer lid was removed and the circulating water switched off. The transparent door was then closed and the shapes were introduced.
Glass door removed after delay period
Opaque shutter removed after delay period
Parcit i<
Shape
[image:2.451.151.349.160.485.2]Crab removed during delay period
Fig. I. Diagram of apparatus. The animal is provided with a home made of three bricks. The 'home' part of the tank is covered with a lid and separated from the rest of the tank by an opaque screen, in which the aperture is closed by a transparent door. The open part of the tank has no lid and contains the screen, partition and shapes, o.t., open part of tank.
A live 20 g. crab tied to a piece of cotton was introduced alongside one of the shapes. This was usually sufficient to cause the octopus to attempt forcefully to get past the door. However, during the early stages of the experiment it was occasionally necessary to show the crab immediately behind the door to provoke this type of response. On these occasions as soon as the attack was provoked the crab was towed slowly across the floor of the tank until it lay alongside one or other of the shapes.
Delayed responses in Octopus 395
After varying lengths of time, 10, 20 or 30 sec. in different experiments, both the transparent door and the mask were removed together, and the octopus was then free to attack the shapes. The time taken to attack was measured with a stop-watch. At-tacks on the shape of the side on which the crab had been shown were scored positive, and rewarded with a 0-5 g. piece offish. Attacks on the other side were scored negative, and punished with an 8 V. d.c. electric shock. Record was also made of those trials in which the animal only made as if to attack the crab while it was shown together with the shape, but did not attack when the shutters were removed. These responses were distinguished from the 'no-response' reaction where the octopus did not attempt to attack the crab even when shown it in close proximity through the transparent door. The side upon which the crab was shown was randomly varied to exclude any pos-sibility of the animal learning to attack on a specific side. In critical experiments the experimenter remained hidden from the animal (p. 399).
The octopuses were isolated, and tested for two or three sessions each day, with five trials in each session, each trial being at least 5 min. after the previous trial.
Normal animals
Eight octopuses that had been in the laboratory for some time were isolated in tanks, as described. They were divided into three groups. Two animals were delayed for
[image:3.451.70.373.379.606.2]10 sec., two for 20 sec., and four were delayed for 30 sec.
Table 1. Responses by normal animals
(Column 7 shows the number of times that there was no attack although there was an active response whilst the crab was shown.)
Delay period (sec.)
(0
10 Totals 20 Totals 3° Totals No. of animals used (relevant graph no.) <»)2 (1, 2)
2 (3,4)
4 (5- 6, 7, 8)
Session (3) 1 2 3 1 2 3 1 2 3 4 Attacks Trials (4) 8/8 7/10 9/10 24/28 6/8 10/10 10/10 26/28 7/18 11/20 13/20 10/15 4i/73 Correct (S) 7 6 8 21 4 7 9 20 4 8 12 10 34 Incorrect (6) 1 1 1 3 2 3 1 6 3 3 1 0 7 No attack (7) 3 1 4 2 0 0 2 11 9 7 5 32 RESULTS 10 sec. delay
396
especially those of the last two sessions, the octopuses swam directly to the shape and attacked it, remaining firmly attached until rewarded. During the first session the animals sometimes walked along the bottom of the tank to attack the shape.
The results of the 10 sec. delay experiments are shown in Table i and Fig. 2, graphs (i) and (2).
20 sec. delay
These two animals rapidly showed the correct response. One of them (octopus 4) first made three incorrect responses and then had a run of nine correct responses. The other (octopus 3) after six correct responses had one long-time and one short-time incorrect response before making further correct responses.
Fig. 2 Fig. 3
Figs. 2 and 3. The graphs show the individual performance of each octopus. Each point shows the time taken to attack the correct ( + ) or incorrect (—) figure at a single trial. A point on the base line represents a 'no attack', and a O represents 'no response to crab' (p. 39s). \ S represents ' shaking up' of the octopus (p. 397). The ordinates are calibrated in seconds.
Delayed responses in Octopus 397
30 sec. delay
In this group four animals were used. In the first session of eighteen trials there were only seven attacks, four of them correct and three incorrect. The remaining •eleven trials provoked attacks when the crabs were shown, but the animals did not
attack after the screens had been removed.
During the second session of the day there were twenty trials with eleven attacks, •eight of them correct. One animal (octopus 6) made definite swimming attacks rapidly and directly at the correct shape, wheareas the others more frequently attacked either positive or negative sides by walking along the bottom of the tank and then tentatively grasping the shape with one or two tentacles. They rapidly swam back into the home if disturbed. Introducing the fish reward into the tank was often sufficient to cause the animal to swim rapidly into its home without seizing the reward.
During this session the attacks of one animal (octopus 6) had been so accurate and rapid that in the third session the animal was made to retreat from the doorway at the end of the crab-showing phase of the trial by a rapid movement of the black shutter towards the transparent door. This was sufficient to cause the animal to withdraw into its home. This stage of the experiment is indicated by 4. S in Fig. 2, graph (6). The introduction of this disturbance did not seem to affect the animal's performance, except sometimes to prolong the response time.
Next morning a further session was undertaken. Unfortunately, one of the animals (octopus 7) had escaped from its tank overnight, and died. In the fifteen trials with the remaining animals, there were ten attacks all of which were correct. Most of the attacks in this session were swimming attacks.
The results of the 30 sec. delay experiments are shown in Table 1 and Fig. 2, graphs
(5H8)-Operated animals
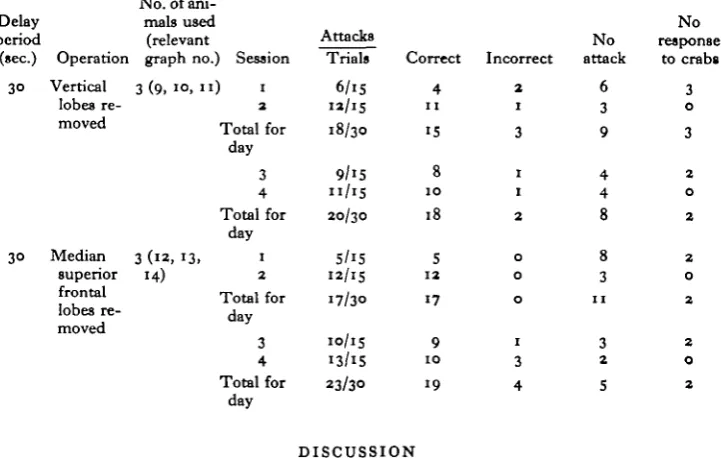
Six animals that had previously only been trained to attack crabs were operated upon, three for the removal of the vertical lobes and three for the removal of the median superior frontal lobes. The operations for the removal of the lobes and tech-nique for assessment of the amounts removed were as described by Boycott & Young (1955). In no animal did more than 25 % of the operated lobe remain.
The animals were replaced in their tanks with the screens in place and crabs were shown to them until the fourth post-operative day, when the experiment was begun as for the normal animals. In each case the delay period was 30 sec.
No vertical lobes [three animals)
During the first session of fifteen trials there were only six attacks, four of them correct. In three trials one animal (octopus 10) showed no response even when crabs were shown. In the second session of the day there were twelve attacks, eleven of them correct. On the first day there were thus thirty trials with eighteen attacks, fifteen of them correct.
398
No median superior frontal lobes {three animals)
In the first session there were only five attacks, but they were all correct. In the second session, there were twelve attacks, all correct. The animals were attacking so rapidly and consistently that it was decided to ' disturb' them during the delay periods of the next day's trials. This was done in the same way as for the 30 sec. delay animal (octopus 6) (p. 397). During the second day's sessions this procedure had little effect on the number of attacks made, although there were now four incorrect attacks in a total of twenty-three attacks. The over-all time for a correct attack was also slightly increased. The results of these experiments are shown in Table 2 and Fig. 3, graphs ( i z H H )
-Table 2
(Columns and indices as for Table i, except the final column, which shows the number of occasions when the animals showed no response to crabs.)
Delay period (sec.) 30 3° Operation Vertical lobes re-moved Median superior frontal lobes re-moved
No. of ani-mals used (relevant
gTaph no.) Session
3 (9, 10, 11) 1
2 Total for day 3 4 Total for day
3 (12, 13, 1
14) 2
Total for day 3 4 Total for day Attacks Trials 6/15 n/15 18/30 9/i5 n/15 20/30 5/15 12/15 17/30 10/15 13/15 23/30 Correct 4 11 15 8 1 0 18 5 1 2 17 9 1 0 19 DISCUSSION Incorrect 2 1 3 1 1 2 0 0 0 1 3 4 No attack 6 3 9 4 4 8 8 3 11 3 2 5 No response to crabs 3 0 3 2 0 2 2 0 2 2 0 2
It is clear that an octopus can choose the correct response even after the object stimulating that response has been removed from the visual field. In the 10 sec. delay experiments the correct response was made immediately, whereas with the 20 and 30 sec. delay experiments there was some sign that the animals had to learn to make the delayed response (Table 1).
The maximum delay period that can be tolerated has not been established. The numbers of attacks made compared with the total possible did not vary much between the 10 and 20 sec. delay groups, but was considerably reduced in the 30 sec. delay group. One interesting feature in all groups was the small number of incorrect attacks (Table 3).
[image:6.451.47.408.251.480.2]Delayed responses in Octopus 399
The possibility that the observer was introducing some extra signal into the ex-perimental situation was controlled by the observer retiring and remaining completely hidden from the animal during the critical experiments. This was possible by adopting such a position that the octopus only came into the observer's field of view when it had reached the shape.
Table 3
State Normal Normal Normal
No vertical lobes No median superior
frontal lobes
Delay period Seconds
1 0 2 0
30
3°
3°
0 / Attacks / 0
Trials 86
93
56 63 67
0/ Correct attacks
Total attacks 88
77 83
87
9 0
The experiment of Schiller (1949) was comparable only in that the stimulus was not continuously in the field of vision of the animal. The octopus was able to see a crab through a wire-netting screen. On removal of a door the animal was able to pass down an opaque tunnel, and when it emerged from the tunnel it had to turn either to the left or right in order to obtain the crab. With doors at either end of the tunnel the delay between seeing the crab and being able to get it could be varied. There were four animals in the experiment, and if they were allowed to pass through the passage-way at once they practically never made an incorrect response. With delays of 40 sec. 75~8o% correct responses were obtained, and one animal was still able to perform accurately after a delay of 1 min. However, it was then shown that the correct per-formance was correlated with maintenance of bodily orientation; the octopus kept its underside towards the crab and crawled along the tunnel on the same side as the crab. When this postural orientation was disorganized by making the octopus squeeze through a small hole in the door, the animal made only random responses, turning equally often to left or right.
In the present experiments the disorientation produced by disturbing the animals during the delay period without changing the accuracy seems to dispose of Schiller's suggestion that the animals were orientating themselves and positioning themselves to ' record' the correct shape to attack while the crab was being shown, and remaining in this position during the delay period. Besides these critical experiments, in many others during the showing of the crab the octopus was in continuous movement, restlessly attempting to pass the transparent door, and adopting no obvious orientation. However, if the octopus does not maintain a record of the 'correct' direction in this way it must do so by means of some other internal record.
subordinate to that of ensuring that the signals indicating food or pain arrive at the proper addresses during learning.
In view of the evidence that this circuit is involved in ' amplification' of ' rewards' and hence of 'motivation' it might have been expected to influence delayed responses. Evidently in Octopus the capacity to retain a record of the recent position of the prey is not dependent on these lobes, although it has been suggested that they are involved in short-term memory (Boycott & Young, 1955). The ability of the octopus to perform delayed responses is interesting since such situations may occur frequently during the natural life of the animal when its prey disappears behind weeds or rocks. The ability to remember temporarily the position at which the prey disappeared will then be vital. In the experimental situation the octopus has two distinct learning tasks. It must learn to attack the shape on the side upon which the crab had been shown; the capacity to do this is needed again at every trial. It must also at each trial carry a record of one particular side during the delay period and then afterwards erase that record in preparation for the next trial.
It is difficult to understand how the animal is able to choose correctly from the identical shapes after the delay period. There are at least two possibilities: (1) There may be some cues, unsuspected by the experimenter, in the target area, enabling the octopus to distinguish the two shapes. (2) The octopus may have some internal capacity that allows it to distinguish left from right. The nature of this capacity, 'representation' or 'model' remains a puzzle. The internal capacity to distinguish left from right must be quite robust since it is capable of surviving violent disturbance to the animal as a whole during the 'shaking up' experiments.
SUMMARY
1. Octopuses were tested for their ability to perform delayed responses after varying delays and brain lesions.
2. Octopuses delayed for 10 sec. were capable of accurate performance without training.
3. In order to make a correct response after 20 or 30 sec. delays some 'learning' is needed.
4. Animals without vertical lobes or without median superior frontal lobes delayed for 30 sec. responded correctly.
5. The majority of correct attacks were 'swimming attacks' in which the animal swam through the water without contacting the walls of the tank.
6. ' Shaking up' the animals and disturbing them so that any orientating contact between the octopus and the door and sides of the tank was destroyed, did not de-crease the accuracy of the response. The time taken for the response in these experi-ments was slightly increased.
Delayed responses in Octopus 401
REFERENCES
BOYCOTT, B. B. & YOUNO, J. Z. (1955). A memory system in Octopui vulgaris Lamarck. Proc. Roy.
Soc. B, 143, 449-80.
HUNTER, W. S. (1912). The delayed reaction in animals and children. Behav. Monogr. a, no. 1. KONORSKI, J. (1959). Physiological mechanism of delayed reactions. Acta Biologiae Experimentalit, 19,
175-SANDERS, F. K. & YOUNO, J. Z. (1940). Learning and other functions of the higher nervous centres of
Sepia. J. Neuropkytiol. 3, 501-26.
SCHILLER, P. H. (1949). Delayed detour response in the octopus. J. Comp. Pkytiol. Ptychol. 4a, 220-5.
SUTHERLAND, N. S. & MUNTZ, W. R. A. (1959). Simultaneous discrimination training and preferred directions of motion in visual discrimination of shape in Octopus vulgarit Lamarck. Pubbl. Stas.
Zool. Napoli, 31, 109-26.