Function of Specific K
1
Channels in Sustained High-Frequency
Firing of Fast-Spiking Neocortical Interneurons
A. ERISIR,1 D. LAU,2 B. RUDY,2 AND C. S. LEONARD1 1
Department of Physiology, New York Medical College, Valhalla 10595; and2Department of Physiology and Neuroscience and Department of Biochemistry, New York University School of Medicine, New York, New York 10016
Erisir, A., D. Lau, B. Rudy, and C. S. Leonard. Function of specific
K1channels in sustained high-frequency firing of fast-spiking neo-cortical interneurons. J. Neurophysiol. 82: 2476 –2489, 1999. Fast-spiking GABAergic interneurons of the neocortex and hippocampus fire high-frequency trains of brief action potentials with little spike-frequency adaptation. How these striking properties arise is unclear, although recent evidence suggests K1 channels containing Kv3.1-Kv3.2 proteins play an important role. We investigated the role of these channels in the firing properties of fast-spiking neocortical interneurons from mouse somatosensory cortex using a pharmacolog-ical and modeling approach. Low tetraethylammonium (TEA) con-centrations (#1 mM), which block only a few known K1channels including Kv3.1-Kv3.2, profoundly impaired action potential repolar-ization and high-frequency firing. Analysis of the spike trains evoked by steady depolarization revealed that, although TEA had little effect on the initial firing rate, it strongly reduced firing frequency later in the trains. These effects appeared to be specific to Kv3.1 and Kv3.2 channels, because blockade of dendrotoxin-sensitive Kv1 channels and BK Ca21-activated K1 channels, which also have high TEA sensitivity, produced opposite or no effects. Voltage-clamp experi-ments confirmed the presence of a Kv3.1-Kv3.2–like current in fast-spiking neurons, but not in other interneurons. Analysis of spike shape changes during the spike trains suggested that Na1channel inactiva-tion plays a significant role in the firing-rate slowdown produced by TEA, a conclusion that was supported by computer simulations. These findings indicate that the unique properties of Kv3.1-Kv3.2 channels enable sustained high-frequency firing by facilitating the recovery of Na1 channel inactivation and by minimizing the duration of the afterhyperpolarization in neocortical interneurons.
I N T R O D U C T I O N
Inhibitory GABAergic interneurons play essential roles in cortical function. They are implicated in the formation and reorganization of receptive fields, in the refinement of cortical connections during development, and in the generation and spread of cortical rhythmical activity (Chagnac-Amitai and Connors 1989; Freund and Buzsaki 1996; Gilbert 1993; Gray 1994; Jacobs and Donoghue 1991; Jones 1993; Martin 1988; Sillito 1984; Singer and Gray 1995; Steriade 1997; Traub et al. 1996; Vidyasagar et al. 1996). Moreover, their dysfunction may be responsible for promoting seizure activity (Hosford 1995; Jefferys and Whittington 1996). Understanding the mechanisms underlying the electrical activity of cortical GABAergic interneurons is therefore critical for understanding
both the normal functioning and pathophysiological processes of the cerebral cortex.
Cortical GABAergic interneurons display diverse intrinsic electrophysiological properties, morphology, connectivity, and neurochemical features (Connors and Gutnick 1990; Huettner and Baughman 1988; Kawaguchi and Kubota 1998; Keller 1995). The interneurons that contain the Ca21binding protein parvalbumin constitute more than half of the cortical interneu-rons (Kubota and Kawaguchi 1994), and a strong correlation between parvalbumin expression and the “fast-spiking” (FS) phenotype has been established in rat neocortex and hippocam-pus by immunocytochemical staining and by single-cell RT-PCR (Cauli et al. 1997; Freund and Buzsaki 1996; Kawaguchi 1995; Kawaguchi and Kubota 1997). These neurons are char-acterized, in vitro and in vivo, by a striking ability to fire sustained high-frequency trains of brief duration action poten-tials with little spike-frequency adaptation in response to sus-tained depolarizing inputs (Azouz et al. 1997; Baranyi et al. 1993; Connors and Gutnick 1990; McCormick et al. 1985; Mountcastle et al. 1969). These distinctive firing properties suggest that FS neurons may express distinct types of ion channels compared with other interneurons and pyramidal cells (Huettner and Baughman 1988), although a provocative anal-ysis of realistic neuronal models suggests that the differential expression of ion channels may not be necessary to account for the diversity of cortical neuron firing properties (Mainen and Sejnowski 1996). In a whole cell patch-clamp study of disso-ciated cortical neurons, Hamill et al. (1991) found that FS neurons had significantly larger K1 currents than pyramidal cells and suggested that a higher K1 channel density might contribute to the differences in firing properties. Recently, the findings that the products of two potassium channel genes, Kv3.1 and Kv3.2, are prominently expressed in parvalbumin-containing fast-spiking cortical interneurons have refined this view and focused interest on the possibility that channels formed from these subunits play a special role in fast-spiking (Chow et al. 1999; Du et al. 1996; Lenz et al. 1994; Martina et al. 1998; Massengill et al. 1997; Moreno et al. 1995; Perney et al. 1992; Sekirnjak et al. 1997; Weiser et al. 1995).
Kv3.1 and Kv3.2 channels display unusual properties when expressed in heterologous expression systems. They are fast-activating delayed rectifiers that require large membrane de-polarizations (above210 mV) to produce significant activation and they deactivate very quickly on repolarization (for review see Rudy et al. 1999). Their rates of deactivation are at least 7–10 times faster than those of other known voltage-gated K1 channels (Coetzee et al. 1999), except for Kv1.7, a
nonneuro-The costs of publication of this article were defrayed in part by the payment of page charges. The article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
nal member of the Kv1 family that deactivates only two to three times slower than Kv3 channels (Kalman et al. 1998). Based on these properties and their distribution patterns in CNS neurons, it has been proposed that Kv3.1 and Kv3.2 channels function in the repolarization of action potentials of short-duration and in facilitating high-frequency firing (Du et al. 1996; Lenz et al. 1994; Martina et al. 1998; Massengill et al. 1997; Moreno et al. 1995; Perney et al. 1992; Perney and Kaczmarek 1997; Sekirnjak et al. 1997; Wang et al. 1998; Weiser et al. 1995). Consistent with this hypothesis, we and others have shown that low doses of 4-aminopyridine (4-AP), which block heterologously expressed Kv3.1-Kv3.2 channels, block a similar current and impair spike repolarization in cortical fast-spiking interneurons (Du et al. 1996; Massengill et al. 1997). In a recent study, Martina et al. (1998) confirmed the differential expression of Kv3.1 and Kv3.2 transcripts in fast-spiking hippocampal basket cells and showed that the major component of the K1 current in these cells is similar to heterologously expressed Kv3.1 and Kv3.2 currents.
The role that these channels play in repetitive firing is less clear. In a study of cultured neocortical neurons (Massengill et al. 1997), 4-AP (0.1 mM) reduced the firing rate of neurons expressing Kv3.1 transcripts, but these cells had very low maximal firing rates (25 spikes/s) compared with those re-ported for FS neurons in slices at the same temperature (104 spikes/s) (Cauli et al. 1997), leaving open the role of Kv3.1-Kv3.2 channels in high-frequency firing. In hippocampal bas-ket cells, application of 4-AP (0.2 mM) in the presence of Ca21-channel blockade interfered with repetitive firing and produced large spike-afterdepolarizations, at least at the single current strength reported (Martina et al. 1998). These observa-tions, along with a recent report showing that tetraethylammo-nium (TEA), presumably by blocking Kv3 channels, affected the ability of auditory neurons to respond to high-frequency stimuli (Wang et al. 1998), are consistent with a general role in high-frequency firing. However, a systematic study of the effects of Kv3.1-Kv3.2 channel blockade on the firing patterns of cortical interneurons, which may lead to an understanding of the mechanisms by which these channels regulate fast-spiking, is still lacking.
We have therefore utilized a pharmacological and computer modeling approach to investigate the specific roles played by a Kv3.1-Kv3.2-like current in the generation of the FS pheno-type in neocortical interneurons. The data showed that a Kv3.1-Kv3.2–like current is necessary to maintain sustained, but not early high-frequency firing. Analysis of the spike shape changes occurring during a train of action potentials suggested that Kv3.1-Kv3.2 currents facilitate sustained high-frequency firing by limiting the accumulation of Na1 channel inactiva-tion, an hypothesis that was supported by computer modeling. M E T H O D S
Brain slices were prepared from 14- to 32-day-old C57/Bl6 mice (Taconic Farms, Germantown, NY). All procedures complied with National Institutes of Health guidelines for ethical use of animals. Following the induction of deep anesthesia with Halothane, the mice were decapitated and the brains were rapidly removed into an ice-cold, oxygenated Ringer solution that contained (in mM) 121 NaCl, 2.5 KCl, 1.25 NaH2PO4, 2 CaCl2, 1 MgCl, 26 NaHCO3, 20 dextrose,
and 4.2 lactic acid. The cerebrum was blocked at a coronal or parasagital plane, and vibratome-sectioned into 250- to 300-mm-thick
sections. Somatosensory cortex slices were incubated at 35°C for 20 min in oxygenated Ringer solution and then were stored at room temperature, until they were transferred to a submerged recording chamber, which was perfused at 3–5 ml/min with the same Ringer solution at room temperature. Drugs were applied by superfusion. Dendrotoxin I and K (DTX), and tetrodotoxin (TTX) were purchased from Alomone Labs (Jerusalem, Israel), TEA from Research Bio-chemicals, and charybdotoxin (CTX) from Sigma (St. Louis, MO); iberiotoxin (IbTX) was a gift from Dr. Maria L. Garcia (Merck Laboratories).
Neurons were visualized at 3160 –200 magnification with near infrared light (.775 nm) transillumination, using a nuvicon tube camera (VE-1000, Dage, Michigan City, IN) and the DIC optics of a fixed-stage microscope (BX50WI, Olympus, Melville, NY). Cells were selected for whole cell recording from mainly layer II/III on the basis of a nonpyramidal shape and multipolar dendrites. Neurons were recorded in current-clamp mode for the analysis of action potential shape and repetitive firing properties, using an electronic bridge amplifier (Axoclamp 2A, Axon Instruments, Foster City, CA) with the output filter set at 10 kHz. Voltage-clamp measurement of ionic currents was obtained from outside-out macro-patches pulled from neurons that were first characterized under current-clamp conditions using an Axopatch 200A amplifier (Axon Instruments). Macro-patches were obtained by slowly backing the pipette from the cell surface along the long axis of the pipette while monitoring the uncompensated capacitative transients and cell input resistance (Keros and McBain 1997). Once resealing occurred, the pipette tip was further withdrawn to just above the surface of the slice. Ringer containing TTX (1 mM) was superfused to block voltage-gated so-dium currents. Recorded currents were filtered at 5 kHz with a four-pole Bessel filter. Pipettes fabricated from Corning 7052 glass were used directly for current-clamp recordings and were coated with silicone elastomer (Sylgard 184, Dow, Midland, MI) and fire-polished immediately before use for macro-patch voltage-clamp recordings. Current-clamp studies were conducted using a pipette solution con-taining (in mM) 144 K-gluconate, 0.2 EGTA, 3 MgCl2, 10 HEPES,
0.3 NaGTP, and 4 Na2ATP. Voltage-clamp studies were conducted
with a pipette solution containing (in mM) 100 K-gluconate, 10 K4– bis-(o-aminophenoxy)-N,N,N9,N9-tetraacetic acid (BAPTA), 5
KCl, 3 MgCl2, 10 HEPES, 0.3 NaGTP, and 4 Na2ATP to limit any
contribution to the current from Ca21-activated K1 currents that might have been present. Biocytin (0.1%; Sigma) was added to the pipette solution just before use. Potentials were recorded with respect to a Ag/AgCl reference electrode located near the outflow of the chamber. Liquid junction potentials were estimated to be213 and 211 mV for each internal solution, and because the difference was small, the values of membrane potentials were not corrected.
Membrane currents and voltages were controlled and recorded with a computer running PCLAMP7 software (Axon Instruments). Current and voltage signals were sampled at 20 kHz, and analysis of these waveforms was performed using Igor Pro (Wavemetrics) software. Action potential shape parameters were measured from action poten-tials evoked by just-suprathreshold 200-ms current steps from a mem-brane potential near270 mV. Spike amplitude was measured as the difference between the peak and the threshold of the action potential. Spike threshold was determined by finding the potential at which the second derivative of the voltage waveform exceeded 3 times its standard deviation in the period preceding spike onset. The afterhy-perpolarization (AHP) was measured as the difference between the spike threshold and voltage minimum following the action potential peak. Maximum rates of rise and decay of the action potential were computed from the maximum and minimum of the smoothed first derivative of the voltage waveform. Spike width was measured at half the spike amplitude. Spike times were measured by determining the time at which the rising phase of the action potential crossed a fixed threshold potential. Instantaneous frequency (1/interspike interval) was computed from trains of action potentials evoked by 600 ms
duration pulses for the 1st, 2nd, 4th, and the last intervals. Steady-state firing rate was computed as the average of instantaneous frequency for the last five intervals of a train. Instantaneous frequency for the 1st, 2nd, 4th, and the last intervals along with steady-state firing rate were plotted as a function of the injected current strength to construct rate-frequency curves. Current strength was increased at 50-pA incre-ments until spike failure occurred within the 600-ms duration pulse. The maximum steady-state firing rate was computed by averaging the steady-state firing rates from trains evoked by the three current strengths before that which produced spike failure. Spike frequency adaptation was measured during the first 200 ms of such trains [A2005(Freq1st2Freq200 ms)/Freq1st] and the reported A200was an
average of the A200values measured from the traces used to compute
the maximum steady-state firing. Statistical analyses were conducted using the program DataDesk 6 (Data Descriptions, Ithaca, NY). Re-sults are reported as means6SE.
For histochemical characterization of recorded cells, slices were fixed in 4% paraformaldehyde for 2 days at 4°C and then stored refrigerated in 0.01 M phosphate-buffered saline (PBS) containing 30% sucrose and 0.05% sodium azide. A freezing microtome was used to resection slices at 50- to 100-mm thickness before incubation in a monoclonal parvalbumin antibody (Sigma; 1:400 dilution in PB containing 1% bovine serum albumin, 0.75% Triton at room temper-ature for 2 days), followed by FITC-conjugated secondary antibody (anti-mouse, 1:50 dilution in PBS; Fisher Chemicals) to visualize Parvalbumin, and by Texas Red conjugated avidin (1:50 in PBS containing 0.7% Triton) to visualize the biocytin-filled cell. Sections were then mounted on gelatin subbed slides and examined on a confocal microscope.
A computer model of an FS cell was implemented using Nodus 3.2 software running on a Power Macintosh computer. The neuron con-sisted of an isopotential sphere with 16 mm diam, a membrane capacitance (Cm) of 1.0mF/cm
2
, a membrane resistance (Rm) of 10
kVzcm2
, a cytoplasmic resistance (Ri) of 100Vzcm, and a resting
membrane potential of270 mV. The leakage conductance was cho-sen to be 10 nS with a reversal potential of270 mV to approximate the average input resistance of recorded FS neurons. The voltage dependence of each current was modeled using a Hodgkin-Huxley formulation for a transient Na1current, a Kv3.1-Kv3.2, current and a Kv1.3 current. The Na1 current model was derived from currents recorded from hippocampal basket cells (Martina and Jonas 1997) and neocortical interneurons (Huguenard et al. 1988). A two-state Kv3.1-Kv3.2 model was derived from fits of Kv3.1 currents expressed in HEK293 cells (Rudy and Leonard, unpublished data), and the Kv1.3 model was derived from “n”-type currents measured in human T-lymphocytes (Cahalan et al. 1985). The maximum value of gNawas
900 nS, of gKv3.1-Kv3.2was 1,800 nS, and of gKv1.3was 1.8 nS. The
Na1conductance was proportional to m3
h, the Kv3.1-Kv3.2 conduc-tance was proportional to n2
, and the Kv1.3 conductance was propor-tional to n4
. The rate constants for the Na current were
am5~3,020240V!/$exp@~751V!/213.5#21%
bm51.2262/exp~V/42.248!
ah50.0035/exp~V/24.186!
bh52~0.871210.017V!/$exp@~51.251V!/25.2#21% The rate constants for the Kv3.1-Kv3.2 current were
an5~952V!/$exp@~2951V!/211.8#21%
bn5~0.025!/exp~V/22.222! The rate constants for the Kv1.3 current were
an52~0.61610.014V!/$exp@~441V!/22.3#21%
bn50.0043/exp@~441V!/34#
R E S U L T S
Interneurons in mouse somatosensory cortex have diverse firing properties
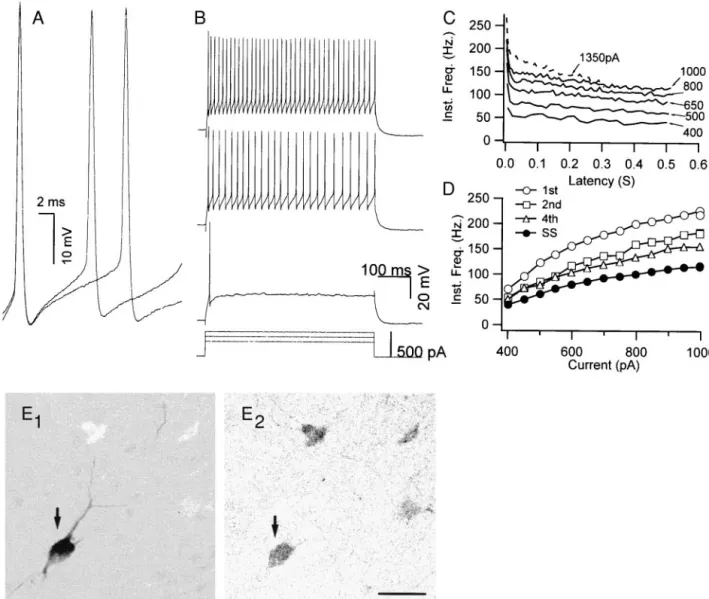
Based on responses to current injection, the recorded non-pyramidal cells were classified into two groups: fast-spiking and regular-spiking-nonpyramidal (RSNP) cells, the latter of which also included cells showing a low-threshold response similar to that described in rat frontal cortex (Kawaguchi and Kubota 1997, 1998). FS neurons (Fig. 1A; n 5 19) had an average resting potential of 260.6 6 1.9 (SE) mV and an average input resistance of 110.4 611 MV. They had short-duration action potentials (0.60 6 0.04 ms) and large AHPs (216.461.2 mV). In response to sustained current injection, FS cells began firing repetitively with abrupt onset and were able to fire at high frequencies with relatively little spike frequency adaptation (A2005 0.34 6 0.02) that occurred mainly over the first few intervals (Fig. 1, B and C). The instantaneous firing rate increased monotonically with current strength (Fig. 1D), and in some cases, this relation reached a clear plateau at current strengths below those that caused spike failure. The average maximum steady-state firing rate for FS neurons was 123.1611 Hz.
It has been shown in quantitative studies both in rat (Sekirn-jak et al. 1997; Weiser et al. 1995) and in mouse (Chow et al. 1998, 1999) that all parvalbumin-containing neurons in the layers I–IV of the neocortex express Kv3.1 proteins, whereas all PV-containing neurons in layers V–VI coexpress both Kv3.1 and Kv3.2 proteins, probably in heteromeric complexes. We therefore used post hoc immunohistochemistry to parval-bumin (Kawaguchi and Kubota 1998) to confirm that the cells having fast-spiking characteristics are Kv3.1-Kv3.2 containing cells (Fig. 1, E1 and E2). These findings also confirmed the strong correlation between parvalbumin expression and the FS phenotype as previously established in rat neocortex (Kawagu-chi and Kubota 1998).
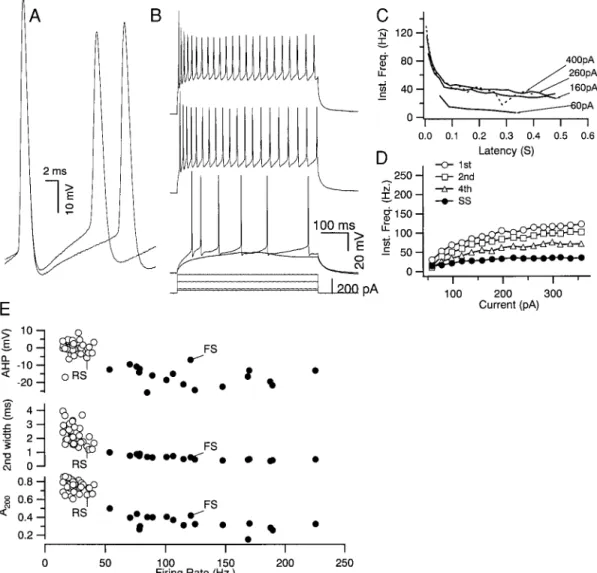
In comparison to FS cells, RSNP neurons (Fig. 2; n537) had significantly longer duration action potentials (1.7160.062 ms;
P, 0.001) and smaller amplitude AHPs (0.16 0.7 mV; P, 0.001), showed much greater spike frequency adaptation (0.756 0.01; P,0.001), and had a much lower maximum frequency of firing (24.8 6 1.1; P , 0.001). RSNP neurons also showed substantial spike broadening (27.864.5%; P,0.001) between the first and second action potentials of a train that was greater than that observed for FS neurons (4 6 0.9%; P , 0.001). Biocytin labeling of some of these RSNP cells (n512) verified that they were nonpyramidal in morphology (data not shown), but none were positive for parvalbumin.
A scatter-plot comparison of spike-shape parameters and maximal average firing frequency revealed a bimodal distribu-tion for all cells with little overlap (Fig. 2E). Thus FS and RSNP neurons were reliably distinguished on the basis of firing frequency, adaptation, and spike shape parameters. Properties of FS and RSNP cells are summarized in Table 1.
Low concentrations of TEA produce striking changes in the action potential of FS neurons
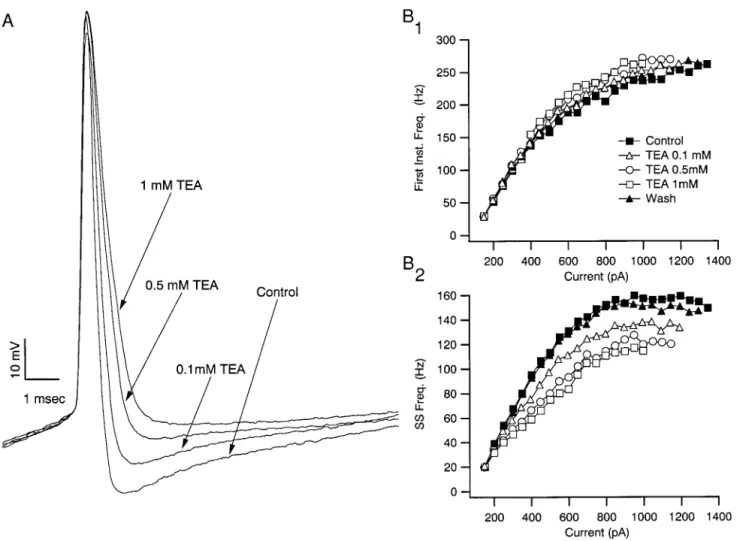
To investigate the effects of blocking Kv3.1-Kv3.2 channels, we exploited their relatively high sensitivity to TEA (extracellular application 1 mM TEA blocks.80% of Kv3.1-Kv3.2 channels
expressed in mammalian heterologous expression systems) (Chandy and Gutman 1995; Grissmer et al. 1994; Herna´ndez-Pineda et al. 1999; Rudy et al. 1999; Vega-Saenz de Miera et al. 1994). Bath application of 1 mM TEA produced a nearly com-plete and reversible elimination of the AHP (Fig. 3A) in all FS neurons tested (212.561.6 vs. 0.861.9 mV, P,0.001; n5 7). TEA also increased the action potential width from 0.646 0.04 to 1.1660.08 ms (P,0.001), suggesting that the current(s) generating the AHP play an important role in repolarizing the action potential. This was supported by the observation that the maximum spike repolarization rate of the first spike during a train of action potentials generated by a just suprathreshold stimulus
was reduced from2148.7628.6 to262.7610.3 mV/ms (P, 0.001), whereas the maximum spike depolarization rate of the first spike remained unaffected (295.7633.6 vs. 295.56 43.6 mV/ms; P50.995). These findings indicate that one or more K1 currents with a high sensitivity to TEA play an important role in repolarizing the action potential and generating the AHP of FS neurons.
TEA impairs the ability of FS neurons to fire high-frequency trains of action potentials
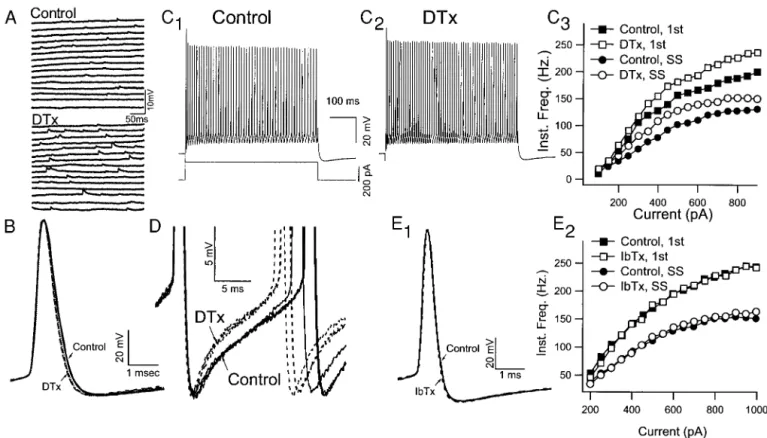
Rather than increasing the firing rate, as might be expected from blocking the AHP, TEA reversibly decreased the steady-FIG. 1. Properties of fast-spiking (FS) neurons recorded from layer II/III somatosensory cortex of mouse. A: FS neurons had
brief duration action potentials and showed little variation in shape between the 1st and 2nd spikes. Action potentials were evoked by just-suprathreshold currents (400 and 500 pA). Traces are superimposed and aligned on the 1st spike. B: repetitive firing of FS neurons for 3 current steps of increasing magnitude (300 –500 pA). Repetitive firing had an abrupt onset in FS cells. In this case, the FS neuron fired only a single spike in response to a 300-pA depolarization but then fired at;50 spikes/s in response to a 400-pA current step. Spike amplitude decreased slightly following the 1st spike. C: instantaneous firing frequency was plotted as a function of time from onset of the current pulse for selected current strengths (solid lines). FS neurons showed only a small amount of spike frequency adaptation. Dashed line indicates the frequency-latency curve for the minimum current strength that produced spike failure during the pulse. D: instantaneous frequency (1/interspike interval) was plotted as a function of injected current. The frequencies of the 1st, 2nd, and 4th intervals and the steady-state frequency (SS) were monotonically proportional to the current strength. The amount of adaptation increased with current intensity, and the relation between firing rate and current showed evidence of reaching a plateau at the highest frequencies. E1 and E2: biocytin-labeled FS neurons (E1) were immunoreactive for parvalbumin (E2). Data from A–E are all from the same FS neuron. Scale bar540mm.
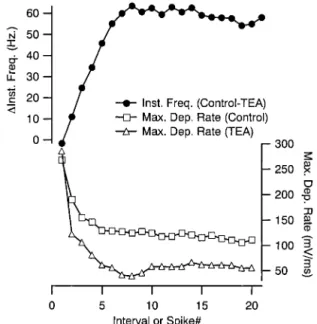
state firing rate of FS neurons (Fig. 3B). This occurred at all current strengths tested, indicating that strong depolarization could not overcome the effect of channel blockade. The aver-age maximum steady-state firing rate was reduced (from 104.66 10.8 to 65.6610 Hz; P, 0.001), and spike failure occurred at lower current strengths than observed in the control condition. Interestingly, no systematic reduction in the instan-taneous firing rate of the first interval was observed (Fig. 3C), and in two cases, an initial burst of action potentials occurred at high current strengths (data not shown). Instead, the TEA suppression of firing rate developed during the spike train, and only reached a maximum by;10 intervals (Fig. 4, top). This resulted in a large increase in the amount of spike-frequency adaptation after the application of TEA (from 0.37 to 0.61; P, 0.001). These results suggest that the processes underlying
firing frequency suppression took time to accumulate. One such process is Na1 channel inactivation, which was also implicated by the observation of spike failure at lower current strengths in the presence of TEA. Consistent with a role for such a mechanism, we observed a decrease in the maximum depolarization slope of each action potentials in a train that was much larger in the presence of TEA and that had a time course that matched the TEA produced changes in firing frequency (Fig. 4, bottom). These effects of TEA were not due to a direct action on Na1 channels because the drug did not affect the maximum depolarization slope of the first action potential in a spike train (see previous section) but did decrease it for the second action potential (249.66 42.9 vs. 209.56 39.2 mV/ ms; n57; P,0.01, measured with just suprathreshold current pulses).
FIG. 2. Properties of layer II/III regular-spiking-nonpyramidal (RSNP) neurons compared with FS neurons. A: RSNP neurons had broader 1st action potentials and showed greater spike broadening between the 1st and 2nd spike than did FS neurons (compare with Fig. 1A). B: just-subthreshold current produced a “low-threshold” response for this RSNP neuron. Suprathreshold current produced repetitive firing at comparatively lower frequencies and with substantially more spike frequency adaptation than observed for FS neurons (compare to Fig. 1B). C: instantaneous firing frequency vs. time from onset of current pulse (—) revealed a large amount of frequency adaptation and a lower minimum current producing spike failure (- - -). D: the relation between instantaneous frequency and current plateaued for all intervals at lower frequencies than those observed for FS neurons. Data from A–D are from the same neuron. E: summary of spike and firing parameters for RSNP (E) and FS (●) neurons. The afterhyperpolarization (AHP) amplitude of the 1st spike, width of 2nd action potential (2nd width), and the fraction of adaptation at 200 ms (A200) are plotted
against the maximum steady-state firing frequency (Firing Rate). These data revealed that neurons were naturally grouped into 2 populations. FS and RS indicate the loci of the neurons illustrated in Fig. 1 and Fig. 2, respectively.
Given that Kv3 channels are blocked by TEA with a Kd
;200mM, it was expected that the effects of TEA on action potential shape and repetitive firing would be dose-dependent over the range of 0.1–1 mM, if they were mediated by antag-onizing Kv3 channels (Fig. 5). The amount of inhibition of the AHP, the degree of action potential broadening, the amount of inhibition of spike repolarization rate, and the suppression of steady firing rate, were all dose-dependent and approached saturation as the TEA concentration reached 1 mM. Finally, spike failure during repetitive firing occurred at lower current strengths as the TEA concentration increased over this range
(Fig. 5B2), whereas only minor effects were observed on the early intervals (Fig. 5B1) at all concentrations of TEA.
Specific K1channel types mediate the effects of low TEA concentrations on FS interneurons
Three additional heterologously expressed K1channels are known that have TEA sensitivities similar to that of Kv3 channels (Kd ; 200 mM): the large conductance, Ca
21 -acti-vated K1(BK) channels containing proteins of the Slo family (Kd80 –330mM), and two voltage-gated K1channels, Kv1.1 (Kd; 300mM) and KCNQ2 (90% blocked by 1 mM TEA) (Coetzee et al. 1999). Because KCNQ2 subunits form very slowly activating and deactivating channels (time constants of hundreds of milliseconds to seconds) (Yang et al. 1998), which would not be significantly activated during short action poten-tials, they were not further examined. Although there are no known specific toxins for Kv3.1 and Kv3.2 channels, we were able to examine the contribution of the other two known TEA-sensitive K1channels using available specific toxin an-tagonists.
DTX, which blocks several Kv1 channels including Kv1.1 (Chandy and Gutman 1995; Coetzee et al. 1999; Robertson et al. 1996), produced an irreversible increase in background synaptic activity (Fig. 6A) but had no significant effects on action potential shape (Fig. 6B). Nevertheless, DTX produced significant increases in steady-state firing rate (maximum steady-state firing rate increased from 156.6614.0 to 172.66 TABLE1. Significance levels and within group comparisons
P FS RS
Resting potential, mV 260.6 6 1.9 261.5 6 2.1 Input resistance, MV * 110.4 611 141.1 612.4 Spike amplitude, mV † 78.5 6 2.5 90.4 6 1.8 AHP amplitude, mV † 216.4 6 1.2 20.2 6 0.7 Depolarization slope, mV/ms * 290.9 617.9 255.2 612.0 Repolarization slope, 1st spike † 2161.5 613.9 243.3 6 2.3 Repolarization slope, 2nd spike † 2142.2 614.1 228.8 6 1.8 † Spike width, ms, 1st spike † 0.606 0.04 1.7 6 0.6 Spike width, ms, 2nd spike † 0.646 0.04 2.2 6 0.1 † Maximum SS firing rate, Hz † 123.2 611.1 24.9 6 1.1 A200 † 0.346 0.02 0.756 0.01
Significance levels between the FS and RS groups are indicated in P. Within group comparisons are indicated by brackets. Number of cells in FS is 19 and in RS is 37. FS, fast spiking; RS, regular spiking; AHP, afterhyperpolarization; SS, steady-state. *P,0.05. † P,0.001.
]
]
]
]
FIG. 3. Tetraethylammonium (TEA; 1 mM) reversibly inhibits the AHP and slows repetitive firing of FS neurons. A: action potential of FS neuron before (control), during (TEA), and after (Wash) the application of 1 mM TEA. Low concentrations of TEA broadened the action potential, decreased the maximum rate of repolarization, and blocked the AHP. B1: TEA reversibly slowed the firing frequency evoked by a constant current pulse. Notice the smaller AHP achieved during the interspike interval. B2: 1 mM TEA also increased the susceptibility to spike failure. A current pulse of 1,350 pA evoked a train of action potentials under control conditions (left). Spike amplitude decreased but achieved a stable amplitude. The same current pulse in the presence of 1 mM TEA resulted in a more rapid reduction of spike amplitude and failure (right). C: instantaneous frequency vs. current for the frequencies measured in the 1st interval (1st;■andh) and the steady-state (SS;●andE). Control conditions,■and●. TEA (1 mM;handE)
13.5 Hz; P,0.05). This increase was apparent at both initial and late intervals and over a large range of currents (Fig. 6C). Closer examination of the voltage trajectories between action potentials during steady-state firing revealed that the AHP recovered more rapidly following the application of DTX (Fig. 6D). These data are consistent with an effect of DTX on a K1 current contributing to the late portion of the AHP and indicate that DTX-sensitive channels do not significantly contribute to spike repolarization and the early phase of the AHP.
Application of IbTX (10 –50 nM, n55) or CTX (100 nM,
n51), which block BK channels (Coetzee et al. 1999), did not produce significant changes in the spike shape or repetitive firing properties of FS neurons (Fig. 6E). This finding was further supported by our observation that blocking Ca21 chan-nels with bath application of both cadmium (500 mM) and nickel (500mM) produced only minor changes in the AHP or in the maximum steady-state firing rate (n 5 2; data not shown). The changes in spike shape and repetitive firing pa-rameters produced by TEA, DTX, and IbTX/CTX are summa-rized in Fig. 7.
Low concentrations of TEA block a Kv3.1-Kv3.2-like current in FS neurons but not in RSNP neuron
To confirm that Kv3.1-Kv3.2–like currents were present in neocortical FS neurons and to determine their contribution to the total somatic K1current, membrane currents were recorded from outside-out macropatches pulled from the somata of physiologically identified FS neurons. The use of outside-out macropatches was imperative because the very large whole cell currents recorded from these neurons precluded adequate
volt-age control and temporal resolution. Macropatches were also pulled from RSNP neurons for comparison (Fig. 8). Patches obtained from FS neurons had significantly larger outward currents (534.6 6 140.5 pA steady state, n 5 5) than those from RSNP neurons (182.7664.2 pA steady state, n55; P, 0.05). TEA (1 mM) blocked the majority of the current (69.36 8.4%, n54) from FS neurons but only a smaller portion of the current from RSNP neurons (23.369.8%, n54; P,0.01). Moreover, the tail currents measured at 240 mV from FS neurons decayed much more rapidly than those from RSNP neurons (Fig. 8, A2 and B2). To examine the voltage depen-dence of the TEA-sensitive component, current-voltage (I-V) curves were constructed from subtraction currents (Fig. 8, A3 and B3). In FS neurons, the resulting TEA-sensitive current showed significant activation at potentials more positive than
220 mV, whereas the current from an RSNP neuron activated at more negative potentials (Fig. 8C). The tail currents of the TEA-sensitive components were also very different (Fig. 8D): the current from the FS neuron deactivated as a single fast exponential (t 5 5.9 ms), which compared well to that of Kv3.1 (t;3 ms) and Kv3.2 (t;6 ms) channels measured in heterologous expression systems (Grissmer et al. 1994; Her-na´ndez-Pineda et al. 1999). In contrast, a double exponential function with longer time constants was necessary to fit the sensitive current from the RSNP neuron. Thus the TEA-sensitive currents obtained from the somatic membrane of FS neurons behaved like Kv3.1-Kv3.2 currents, whereas those from layer II/III RSNP neurons, which do not express Kv3.1-Kv3.2 subunits, did not. These data confirm the conclusions from studies in other neurons showing that native Kv3.1 and Kv3.2 channels have properties remarkably similar to those in heterologous expression systems (Du et al. 1996; Herna´ndez-Pineda et al. 1999; Martina et al. 1998; Rudy et al. 1999; Wang et al. 1998) and suggest that factors such as associated subunits or cell-specific postranslational modifications do not signifi-cantly change the electrophysiological properties of native neuronal channels containing Kv3 proteins.
Kv3.1-Kv3.2 channels enabled high-frequency firing by speeding the recovery of sodium-conductance inactivation while minimizing the duration of the afterhyperpolarization
The previous experiments suggest that Kv3.1-Kv3.2 currents facilitate sustained high-frequency firing of FS neurons, in part, by reducing the amount of Na1 channel inactivation that accumulates during the spike train. To test this mechanism further, we constructed a single compartment Hodgkin-Hux-ley–like model. The model included voltage-gated Na1 chan-nels, Kv3.1-Kv3.2 chanchan-nels, and Kv1-like channels (seeMETH -ODS) and was studied under current-clamp conditions with
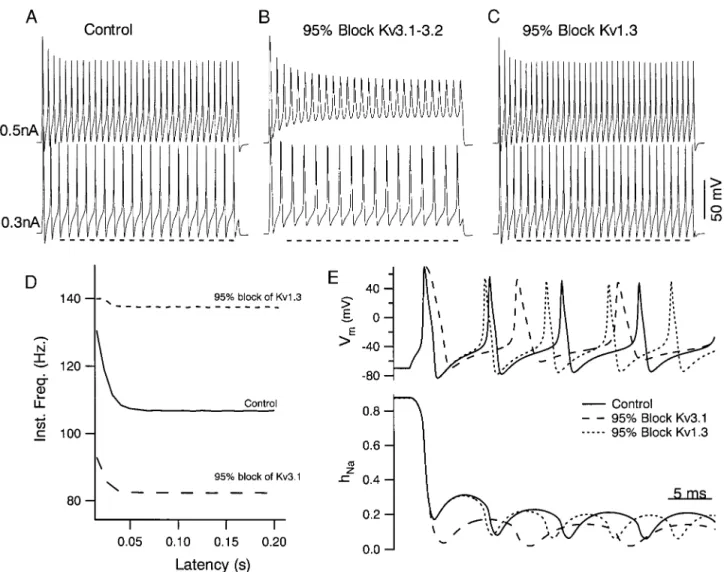
200-ms current pulses (Fig. 9). Under control conditions, when none of the channels were blocked, depolarizing currents pro-duced repetitive firing with an abrupt onset (initial steady-state frequency 62 Hz). Like our recorded FS neurons, the model displayed fast spiking with slight, early, spike frequency ad-aptation. The firing rate varied monotonically with injected current strength, and this relation approached an asymptote as current increased, as observed for FS neurons (data not shown). Blocking the Kv3.1-Kv3.2 channels in the model mimicked the effects of low TEA concentrations on FS neurons: the AHP was decreased, the action potential broadened, and
repolariza-FIG. 4. Firing rate slowdown produced by TEA accumulated during the spike train in tandem with a decrease in spike depolarization rate. The differ-ence between the instantaneous frequency for a control train and one evoked in the presence of 1 mM TEA (1,100-pA current step) is plotted as a function of interval number (top) and required;10 intervals to reach a steady level. This slowdown was correlated with a progressive decrease in the maximum depolarization rate of the action potential. This decrease in spike depolarization rate was greatly accentuated in the presence of TEA, suggesting a larger accumulation of Na1channel inactivation following blockade of TEA-sensi-tive channels.
tion slowed. This result supports the conclusion that a Kv3.1-Kv3.2 current strongly contributes to spike repolarization and the AHP. In addition, blockade of the Kv3.1-Kv3.2 current decreased the repetitive firing rates for all current strengths (Fig. 9, B and D), and spike failure occurred at lower current strengths. In contrast, blocking the Kv1-like current had no effect on the action potential shape but produced an increase, rather than decrease in the firing rate (Fig. 9, C and D), as we observed experimentally when we applied DTX.
How these opposite effects of K1channel blockade on firing rate arose was investigated by examining the changes in chan-nel parameters during action potential trains. The spike-broad-ening produced by blocking Kv3.1-Kv3.2 channels resulted in greater sodium channel inactivation occurring during the action potential (hNa; Fig. 9E). Moreover, due to the blockade of the AHP, the rate of recovery from Na1 channel inactivation following the action potential was slowed down, resulting in a significant decrease in the amount of recovery during the interspike interval. Thus fewer Na1channels were available to depolarize the neuron in the period leading up to the next spike.
A greater membrane depolarization was required before there was enough Na1current to begin the next spike and the next spike was delayed. No such effect was observed on blockade of the Kv1-like current. Because this current contributed little to shaping the action potential, there was no significant effect on the amount of Na1 channel inactivation or on its rate of recovery from inactivation (Fig. 9E). Rather, because the Kv1-like current deactivated slowly, it was not completely turned off during the brief AHP. Moreover, because it activates near spike threshold, it actually grows prior to the next action potential and functions to lengthen the interspike interval. Hence, blocking the Kv1-like current shortens the AHP with-out decreasing the peak amplitude, thereby increasing the firing rate.
Although our model qualitatively reproduces the repetitive firing behavior of FS neurons, there are some quantitative differences. For example, the slow-down in firing frequency produced by Kv3 blockade in the model (Fig. 9D) occurred faster than the slow-down observed in our TEA experiments. This may result from differences between the actual and
mod-FIG. 5. TEA actions on spike shape and repetitive firing occur at low doses. A: 0.1, 0.5, and 1 mM TEA resulted in progressively larger effects on the AHP, spike width, and maximum spike repolarization rate with little effect on the amplitude or rate of rise of the action potential. B1: for this cell there was no effect of TEA on the instantaneous frequency measured from the 1st interspike interval for firing frequencies lower than;150 spikes/s (,400 pA). For larger current strengths, TEA produced a slight increase in the instantaneous frequency that was dose dependent and reversible. B2: the same range of TEA concentration produced a progressively larger decrease in the steady-state firing rate. Similarly, spike failure during repetitive firing occurred at progressively lower current strengths with increased TEA dose. This is indicated by the last point of each curve, which marks the greatest current strength that did not produce failures. These data are consistent with TEA acting on channels having a Kdnear that reported for
FIG. 6. Dendrotoxin I and K (DTX) and iberiotoxin (IbTX) do not mimic the effects of TEA on FS neurons. A: DTX-I (100 nM) application resulted in an increase of spontaneous synaptic potentials. B: 3 superimposed action potentials before (Control; solid lines) and after (DTX; dashed lines) DTX-I application indicate that DTX had no significant effects on spike shape parameters. C1: response of an FS neuron to 250-pA current in normal Ringer. C2: response of FS to the same current pulse after 15 min application of 100mM DTX-I. In contrast to the effects of TEA, DTX significantly increased the firing frequency of FS neurons. C3: instantaneous frequency vs. current pulse amplitude for the 1st interval (■andh) and the steady state (●andE) before (■and
●) and after DTX (h andE). DTX significantly increased the firing rate for both the early and late intervals in the train. D:
superimposed truncated action potentials from an FS cell recorded before (—) and after (- - -) DTX application. DTX resulted in a slightly faster decay of the AHP. E1: IbTX had no significant effect on spike shape or on repetitive firing (E2) of a representative FS neuron.
FIG. 7. Actions of TEA (1 mM), DTX, and IbTX/ charybdotoxin (CTX) are summarized as percent change from control. A: none of the agents had significant effects on spike amplitude (Spike Amp.) or on the maximum rate of rise of the 1st action potential (Max. Rising Slope). TEA (1 mM) significantly slowed the maximum rate of spike repolarization (Max. Repol. Slope). Neither DTX nor IbTX/CTX had significant effects on the Max Repol. Slope. TEA significantly reduced the amplitude of the AHP (AHP Amp.). Neither DTX nor IbTX/CTX had significant effects on the AHP amplitude. TEA signifi-cantly increased the width of the action potential (Spike Width). Neither DTX nor IbTX/CTX had significant ef-fects on the Spike Width. TEA significantly slowed the maximum average steady-state firing rate (Max. Freq.). DTX significantly increased the Max. Freq., whereas IbTX/CTX had no significant effect. TEA significantly increased the amount of spike-frequency adaptation mea-sured at 200 ms (A200). Neither DTX nor IbTX/CTX had
a significant effect. *P,0.01, **P ,0.001, ***P, 0.0001.
eled Na1channel kinetics or from the presence of additional conductances not included in the model. Knowledge of these factors is required before a more accurate model is attempted. D I S C U S S I O N
Using pharmacological and modeling approaches we have demonstrated that the action potential and repetitive firing properties of fast-spiking interneurons in the mouse somato-sensory cortex are powerfully shaped by a K1current closely similar to Kv3.1and Kv3.2 currents. We found that submilli-molar concentrations of TEA disrupted the fast-spiking phe-notype and that this action of TEA on FS neurons was highly specific. Selective toxins, which antagonize other K1channels having a high sensitivity to TEA, had either no effect or facilitated high-frequency firing of FS neurons. Of all K1 channels known (Coetzee et al. 1999), only those containing subunits of the Kv3 subfamily could account for the results of our pharmacological experiments. We also found that the ma-jority of the somatic K1 current from FS (but not RSNP) neurons resulted from a current that strongly resembles the
current expressed by Kv3.1 and Kv3.2 proteins in mammalian heterologous expression systems (Grissmer et al. 1994; Her-na´ndez-Pineda et al. 1999; Rudy et al. 1999). Taken together, the data strongly support the idea that Kv3.1-Kv3.2 channels play a dominant role in repolarizing the action potential and enabling high-frequency firing in neocortical FS neurons, a conclusion that was further supported and extended by our computer simulations.
Just how do Kv3.1 and Kv3.2 channels function in FS neurons? Based on the requirement for membrane depolariza-tion above220 mV to achieve significant activation of heter-ologously expressed Kv3.1-Kv3.2 channels, it has been sug-gested that these channels, when expressed in sufficient numbers, could repolarize action potentials without influencing their threshold, in contrast to K1channels that activate at more negative potentials (Kanemasa et al. 1995; Weiser et al. 1994, 1995). Our findings strongly support this view of Kv3.1 and Kv3.2 channel function. Blockade of a Kv3.1-Kv3.2–like cur-rent by low concentrations of TEA profoundly slowed action potential repolarization in FS neurons without changing the
FIG. 8. Outside-out macro-patches from the somata of FS neurons had a large Kv3.1-Kv3.2–like current, whereas those from RSNP neurons did not. A1: currents from an FS neuron in control Ringer (—) and after the addition of 1 mM TEA (- - -). Voltage jumps from270 to140 mV resulted in a large outward current that was largely blocked by 1 mM TEA. B1: results from similar experiments performed on macro-patches from RSNP neurons indicated, as shown in this example, that RSNP neurons had smaller outward currents and that a smaller fraction of that outward current was blocked by 1 mM TEA. A2 and B2: the time course of tail currents from FS (A2) and RSNP (B2) macro-patches were measured in response to a voltage jump from140 to240 mV. Both sets of tails were fit well by double exponential functions, but the dominant time constant was close to 5 ms for the FS neuron, whereas it was close to 40 ms for the RS neuron. A3 and B3: the TEA-sensitive currents, computed by subtraction, were larger in macro-patches from FS neurons than those from RS neurons. C: the current-voltage (I-V) relation of the TEA-sensitive current from FS neurons (C) showed significant current at voltages above220 mV and was similar to Kv3.1-Kv3.2 currents in heterologous expression systems. In patches from RSNP neurons, the TEA-sensitive current (●) activated at more negative potentials. D: the time course of the TEA-sensitive tail current from an FS neuron was fit well by a single exponential having a 5.9 ms time constant and was similar to that reported for Kv3.1-Kv3.2 channels. In contrast, the time course of the TEA-sensitive tail current from an RSNP neuron was dominated by an exponential with a much slower time constant (62.7 ms) unlike that reported for Kv3.1-Kv3.2 channels.
threshold, the maximum depolarization rate, or the spike am-plitude. These data imply that the Kv3.1-Kv3.2–like channels become sufficiently activated during the brief spike to contrib-ute selectively to action potential repolarization. This point is directly supported by experiments in which transfected HEK293 cells expressing Kv3.1 or Kv3.2 channels were volt-age clamped to an action potential waveform (Rudy et al. 1999). No current was seen until after the action potential reached its peak. The results of our computer simulations mimicked these findings for FS neocortical neurons, and a recent study of hippocampal basket cells indicates that a native
4-AP–sensitive current, which may arise from Kv3.1 and/or Kv3.2 channels, can be activated by brief action potentials (Martina et al. 1998).
In addition to firing brief action potentials, FS neurons have large AHPs compared with other neocortical interneurons and pyramidal cells. Our data strongly indicate that the Kv3.1-Kv3.2–like current is also responsible for this large AHP, because it was abolished by 1 mM TEA but not by blockers of other known channels having comparable sensitivities to TEA. Although a large AHP is often associated with central neurons that fire slowly (Henderson et al. 1982; Leonard and Llina´s
FIG. 9. Blockade of a Kv3.1-Kv3.2 current in a computer model of FS neurons mimics effects of TEA on spike shape and repetitive firing in FS neurons. A: spike trains resulting from 200 ms duration current pulses (0.3, bottom; and 0.5 nA, top) injected into the FS model cell. Spike frequency was related to the strength of current injection. Action potentials in the model showed a small amplitude decrease during the pulse and some early frequency adaptation as observed for FS neurons. B: spike trains resulting from the model after blocking Kv3.1-Kv3.2 channels. The AHP was greatly attenuated (compare to dashed line). Spike frequency for a given current strength also slowed, as observed experimentally with application of low doses of TEA to FS neurons. At higher current strengths, a greater spike amplitude decrease occurred following Kv3.1-Kv3.2 blockade, and spike failure occurred at a lower current strength than for control conditions (data not shown). C: blockade of the Kv1.3 current in the model, after restoring all of the Kv3.1-Kv3.2 current, restored the AHP amplitude and produced an increase in firing rate. Dashed line in A–C is a280 mV reference to help compare the magnitude of the AHP in each condition. D: instantaneous frequency vs. time after pulse onset (Latency) for spike trains evoked by 0.3-nA current steps in each of the conditions illustrated in A–C. E, top: membrane potential (Vm) response to the 0.3-nA current pulse are superimposed at higher temporal resolution for the conditions in A–C to show the
changes in spike shape and repetitive firing that occurred. Bottom: the corresponding time course of Na-channel inactivation (hNa)
for these conditions. Spike broadening and AHP reduction produced by blockade of Kv3.1–3.2 resulted in a greater degree of inactivation and less recovery from inactivation than observed in either the Control condition or with just Kv1.3 blocked. This supports the hypothesis that the spike slowing following Kv3.1–3.2 blockade results from greater accumulation of Na-channel inactivation.
1990; Yarom et al. 1985), where it functions to slow firing, we found that the large AHP (and brief action potential) generated in FS neurons functions to enable high-frequency firing. This function appears to result directly from the voltage dependence and rapid deactivation kinetics of native Kv3.1 and Kv3.2 channels and the apparently low levels of Ca21-activated K1 currents and other K1 currents having slower deactivation kinetics and more negatively shifted voltage dependencies.
Our studies suggest that an important mechanism by which Kv3.1-Kv3.2–like channels enable fast spiking is by limiting the impact of Na1channel inactivation on repetitive firing. In trains of action potentials, the interspike interval is established, in part, by the amount of Na1channel inactivation that accu-mulates during the train. By keeping action potentials brief, Kv3.1-Kv3.2 currents reduce the amount of Na1 channel in-activation that occurs during the action potential. This was evident in our simulations where a substantial increase in the amount of Na1channel inactivation occurred following spike broadening produced by Kv3.1-Kv3.2 channel blockade. Kv3.1-Kv3.2 currents also function to speed recovery from Na1-channel inactivation by generating a large AHP. Results from our simulations support this idea because blockade of Kv3.1-Kv3.2 currents both slowed the recovery of Na1 chan-nel inactivation and reduced the amount of recovery that oc-curred after an action potential. A role for Na1channel inac-tivation in decreasing firing frequency was also evident in our FS recordings. TEA greatly enhanced the reduction in depo-larization rate that occurred with successive spikes in a train. It is also worth noting that another factor by which the blockade of Kv3 channels could slow firing frequency is by an increased activation of other possible conductances in response to the broadening of the action potential. The contribution of these possible factors remains to be investigated.
Finally, as was evident in our simulations, the large magni-tude of the AHP in FS neurons also functions to terminate the Kv3.1-Kv3.2 current, which, because of its rapid deactivation rates, and positive activation voltage, minimizes the duration of the refractory period. The situation was completely different for K1 channels that activate at more negative potentials and have slower deactivation kinetics in our simulations. Due to these factors, the Kv1-like conductance decayed little during the interspike interval and contributed to delaying the onset of the next spike. Hence blocking that current in the model produced an increase in spike frequency. Collectively, these results strongly suggest that the particular activation range and fast deactivation kinetics of Kv3.1-Kv3.2 channels function to enable sustained high-frequency firing in FS neurons.
Kv3 proteins are found in the somata of many other neurons capable of high-frequency firing, including some, but not all, GABAergic neurons, suggesting a similar role in facilitating high-frequency firing for these neurons. Kv3 genes are also prominently expressed in many neurons that process sensory information, including many auditory structures (Perney and Kaczmarek 1997; Perney et al. 1992; Rudy et al. 1992; Weiser et al. 1994, 1995). For example, Kv3.1 and Kv3.3 transcripts are found in neurons of the medial nucleus of the trapezoid body (MNTB). These neurons do not fire sustained high-frequency trains of action potentials in response to steady current injection, however, they can fire action potentials en-trained to very high-frequency inputs (.600 Hz), which pre-serves the timing information contained in auditory signals
(Brew and Forsythe 1995). Clearly, the presence of Kv3 chan-nels alone is not sufficient for the generation of sustained high-frequency firing. Nevertheless, Kv3 channels do appear to function in the high-frequency firing of these neurons because low concentrations of TEA reduced their ability to follow stimulus frequencies.200 Hz. (Wang et al. 1998). The dif-ferent firing properties between MNTB neurons and neocorti-cal FS neurons could be explained, in part, by the different levels of low-voltage–activating DTX-sensitive K1 channels (Brew and Forsythe 1995; Wang et al. 1998).
K1 channel diversity is a main factor contributing to the diversity of the electrophysiological properties of neurons, and it also contributes to the specificity of neuromodulator actions (Adams and Galvan 1986; Baxter and Byrne 1991; Hille 1992; Kaczmarek and Levitan 1987; Llinas 1988; Rudy 1988). The large number of K1channel subunits discovered in the last 10 years unexpectedly exceeds the diversity predicted from elec-trophysiological studies of native K1 currents. Over 100 dif-ferent pore-forming subunits of mammalian K1channels have been discovered (Coetzee et al. 1999). Interactions among different subunits and other factors suggest the existence of hundreds if not thousands of different types of K1channels. A significant challenge lies in integrating this molecular diversity into a physiological context. Kv3 channels represent a case in point. Before the isolation of Kv3 cDNAs and the character-ization of Kv3 channels in heterologous expression systems, the existence of these channels in neurons as a separate channel type was apparently undetected. Kv3 channels had not been separated from other components of the K1current, and some of the initial papers on the cloning of Kv3 cDNAs suggested that their properties in neurons might be different (McCormack et al. 1990; Rudy et al. 1991; Vega-Saenz de Miera et al. 1992). The characterization of the electrophysiological and pharma-cological properties of Kv3 currents in heterologous expression systems and the delineation of their expression patterns in the CNS provided clues that have allowed the isolation of native Kv3 currents in neurons and the generation of hypotheses as to their functional roles in the CNS. We now provide strong evidence in favor of the hypothesis that Kv3.1-Kv3.2 channels play specific roles in the generation of sustained high-fre-quency firing in cortical interneurons. It is expected that similar strategies with other cloned K1 channel proteins whose neu-ronal roles are unknown will result in the discovery of addi-tional previously unknown native K1 channels and novel mechanisms to regulate neuronal function.
We thank Dr. S. Hestrin for help and advice about recording from neocor-tical interneurons.
This research was supported by National Institute of Neurological Disorders and Stroke Grant NS-27881 and National Science Foundation Grant IBN989606 to C. S. Leonard and National Institute of Neurological Disorders and Stroke Grants NS-30989 and NS-35215 and National Science Foundation Grant IBN9630832 to B. Rudy.
Address for reprint requests: C. S. Leonard, Dept. of Physiology, New York Medical College, Valhalla, NY 10595.
Received 21 June 1999; accepted in final form 26 August 1999.
REFERENCES
ADAMS, P. R. AND GALVAN, M. Voltage-dependent currents of vertebrate neurons and their role in membrane excitability. Adv. Neurol. 44: 137–170, 1986.
AZOUZ, R., GRAY, C. M., NOWAK, L. G.,ANDMCCORMICK, D. A. Physiological properties of inhibitory interneurons in cat striate cortex. Cereb. Cortex 7: 534 –545, 1997.
BARANYI, A., SZENTE, M. B.,ANDWOODY, C. D. Electrophysiological char-acterization of different types of neurons recorded in vivo in the motor cortex of the cat. II. Membrane parameters, action potentials, current-induced voltage responses and electrotonic structures. J. Neurophysiol. 69: 1865–1879, 1993.
BAXTER, D. A.ANDBYRNE, J. H. Ionic conductance mechanisms contributing to the electrophysiological properties of neurons. Curr. Opin. Neurobiol. 1: 105–112, 1991.
BREW, H. M.ANDFORSYTHE, I. D. Two voltage-dependent K1conductances with complementary functions in postsynaptic integration at a central audi-tory synapse. J. Neurosci. 15: 8011– 8022, 1995.
CAHALAN, M. D., CHANDY, K. G., DECOURSEY, T. E., AND GUPTA, S. A voltage-gated potassium channel in human T lymphocytes. J. Physiol.
(Lond.) 358: 197–237, 1985.
CAULI, B., AUDINAT, E., LAMBOLEZ, B., ANGULO, M. C., ROPERT, N., TSUZUKI, K., HESTRIN, S.,ANDROSSIER, J. Molecular and physiological diversity of cortical nonpyramidal cells. J. Neurosci. 17: 3894 –3906, 1997.
CHAGNAC-AMITAI, Y.ANDCONNORS, B. W. Horizontal spread of synchronized activity in neocortex and its control by GABA-mediated inhibition. J.
Neu-rophysiol. 61: 747–758, 1989.
CHANDY, K. G.ANDGUTMAN, G. A. Voltage gated channels. In: Handbook of
Receptors and Channels: Ligand-Gated and Voltage-Gated Ion Channels,
edited by R. A. North. Boca Raton, FL: CRC, 1995, p. 1–71.
CHOW, A., ERISIR, A., FARB, C., LAU, D.H.P.,ANDRUDY, B. Kv3.1 and Kv3.2 proteins distingish three subpopulations of GABA-ergic interneurons in the mouse cortex. Soc. Neurosci. Abstr. 24: 1579, 1998.
CHOW, A., ERISIR, A., FARB, C., WELKER, E., LAU, D.,AND RUDY, B. K1 channel expression distinguishes subpopulations of parvalbumin- and so-matostatin-containing cortical interneurons. J. Neurosci. In press. COETZEE, W. A., AMARILLI, Y., CHIU, J., CHOW, A., LAU, D., MCCORMACK, T.,
MORENO, H., NADAL, M. S., OZAITA, A., POUNTNEY, D., SAGANICH, M., VEGA-SAENZ DE MIERA, E., AND RUDY, B. Molecular diversity of K1 channels. Ann. NY Acad. Sci. 868: 233–286, 1999.
CONNORS, B. W. AND GUTNICK, M. J. Intrinsic firing patterns of diverse neocortical neurons. Trends Neurosci. 13: 99 –104, 1990.
DU, J., ZHANG, L., WEISER, M., RUDY, B.,ANDMCBAIN, C. J. Developmental expression and functional characterization of the potassium-channel subunit Kv3.1b in parvalbumin-containing interneurons of the rat hippocampus.
J Neurosci. 16: 506 –518, 1996.
FREUND, T. F.ANDBUZSAKI, G. Interneurons of the hippocampus.
Hippocam-pus 6: 347– 470, 1996.
GILBERT, C. D. Circuitry, architecture, and functional dynamics of visual cortex. Cereb. Cortex. 3: 373–386, 1993.
GRAY, C. M. Synchronous oscillations in neuronal systems: mechanisms and functions. J. Comput. Neurosci. 1: 11–38, 1994.
GRISSMER, S., NGUYEN, A. N., AIYAR, J., HANSON, D. C., MATHER, R. J., GUTMAN, G. A., KARMILOWICZ, M. J., AUPERIN, D. D.,ANDCHANDY, K. G. Pharmacological characterization of five cloned voltage-gated K1channels, types Kv1.1, 1.2, 1.3, 1.5, and 3.1, stably expressed in mammalian cell lines.
Mol. Pharmacol. 45: 1227–1234, 1994.
HAMILL, O. P., HUGUENARD, J. R.,ANDPRINCE, D. A. Patch-clamp studies of voltage-gated currents in identified neurons of the rat cerebral cortex. Cereb.
Cortex. 1: 48 – 61, 1991.
HENDERSON, G., PEPPER, C.,ANDSHEFNER, S. Electrophysiological properties of neurones contained in the locus coeruleus and mesencephalic nucleus of the trigeminal nerve in vitro. Exp. Brain Res. 45: 29 –37, 1982.
HERNANDEZ´ -PINEDA, R., CHOW, A., AMARILLO, Y., MORENO, H., SAGANICH, M., VEGA-SAENZ DE MIERA, E., HERNANDEZ´ -CRUZ, A., AND RUDY, B. Kv3.1–Kv3.2 channels underlie a high voltage–activating component of the delayed rectifier K1current in projecting neurons from the globus pallidus.
J. Neurophysiol. 82: 1512–1528, 1999.
HILLE, B. Ion Channels of Excitable Membranes. Sunderland, MA: Sinauer, 1992.
HOSFORD, D. A. Models of primary generalized epilepsy. Curr. Opin. Neurol. 8: 121–125, 1995.
HUETTNER, J. E.ANDBAUGHMAN, R. W. The pharmacology of synapses formed by identified corticocollicular neurons in primary cultures of rat visual cortex. J. Neurosci. 8: 160 –175, 1988.
HUGUENARD, J. R., HAMILL, O. P.,ANDPRINCE, D. A. Developmental changes in Na1conductances in rat neocortical neurons: appearance of a slowly inactivating component. J. Neurophysiol. 59: 778 –795, 1988.
JACOBS, K. M.ANDDONOGHUE, J. P. Reshaping the cortical motor map by unmasking latent intracortical connections. Science 251: 944 –947, 1991. JEFFERYS, J. G.ANDWHITTINGTON, M. A. Review of the role of inhibitory
neurons in chronic epileptic foci induced by intracerebral tetanus toxin.
Epilepsy Res. 26: 59 – 66, 1996.
JONES, E. G. GABAergic neurons and their role in cortical plasticity in primates. Cereb. Cortex 3: 361–372, 1993.
KACZMAREK, L. K.ANDLEVITAN, I. B. Neuromodulation. New York: Oxford, 1987.
KALMAN, K., NGUYEN, A., TSENG-CRANK, J., DUKES, I. D., CHANDY, G., HUSTAD, C. M., COPELAND, N. G., JENKINS, N. A., MOHRENWEISER, H., BRANDRIFF, B., CAHALAN, M., GUTMAN, G. A.,ANDCHANDY, K. G. Genomic organization, chromosomal localization, tissue distribution, and biophysical characterization of a novel mammalian Shaker-related voltage-gated potas-sium channel, Kv1.7. J. Biol. Chem. 273: 5851–5857, 1998.
KANEMASA, T., GAN, L., PERNEY, T. M., WANG, L. Y.,ANDKACZMAREK, L. K. Electrophysiological and pharmacological characterization of a mammalian Shaw channel expressed in NIH 3T3 fibroblasts. J Neurophysiol. 74: 207– 217, 1995.
KAWAGUCHI, Y. Physiological subgroups of nonpyramidal cells with specific morphological characteristics in layer II/III of rat frontal cortex. J. Neurosci. 15: 2638 –2655, 1995.
KAWAGUCHI, Y.ANDKUBOTA, Y. GABAergic cell subtypes and their synaptic connections in rat frontal cortex. Cereb. Cortex 7: 476 – 486, 1997. KAWAGUCHI, Y.ANDKUBOTA, Y. Neurochemical features and synaptic
con-nections of large physiologically-identified GABAergic cells in the rat frontal cortex. Neuroscience 85: 677–701, 1998.
KELLER, A. Synaptic organization of the barrel cortex. In: Cerbral Cortex, edited by E. G. Jones and I. P. Diamond. New York: Plenum, 1995, p. 221–262.
KEROS, S.ANDMCBAIN, C. J. Arachidonic acid inhibits transient potassium currents and broadens action potentials during electrographic seizures in hippocampal pyramidal and inhibitory interneurons. J. Neurosci. 17: 3476 – 3487, 1997.
KUBOTA, Y.ANDKAWAGUCHI, Y. Three classes of GABAergic interneurons in neocortex and neostriatum. Jpn. J. Physiol. 44: S145–S148, 1994. LENZ, S., PERNEY, T. M., QIN, Y., ROBBINS, E., AND CHESSELET, M. F.
GABA-ergic interneurons of the striatum express the Shaw-like potassium channel Kv3.1. Synapse 18: 55– 66, 1994.
LEONARD, C. S.ANDLLINAS´ , R. Electrophysiology of mammalian pedunculo-pontine and laterodorsal tegmental neurons in vitro: implications for the control of REM sleep. In: Brain Cholinergic Systems, edited by M. Steriade and D. Biesold. New York: Oxford, p. 205–223, 1990.
LLINAS, R. The intrinsic electrophysiological properties of mammalian neu-rons: insights into central nervous system function. Science 242: 1654 – 1664, 1988.
MAINEN, Z. F.ANDSEJNOWSKI, T. J. Influence of dendritic structure on firing pattern in model neocortical neurons. Nature 382: 363–366, 1996. MARTIN, K. A. The Wellcome Prize lecture. From single cells to simple circuits
in the cerebral cortex. Q. J. Exp. Physiol. 73: 637–702, 1988.
MARTINA, M.AND JONAS, P. Functional differences in Na1channel gating between fast-spiking interneurones and principal neurones of rat hippocam-pus. J. Physiol. (Lond.) 505: 593– 603, 1997.
MARTINA, M., SCHULTZ, J. H., EHMKE, H., MONYER, H., AND JONAS, P. Functional and molecular differences between voltage-gated K1channels of fast-spiking interneurons and pyramidal neurons of rat hippocampus. J.
Neu-rosci. 18: 8111– 8125, 1998.
MASSENGILL, J. L., SMITH, M. A., SON, D. I.,ANDO’DOWD, D. K. Differential expression of K4-AP currents and Kv3.1 potassium channel transcripts in cortical neurons that develop distinct firing phenotypes. J. Neurosci. 17: 3136 –3147, 1997.
MCCORMACK, K., LIN, J. W., IVERSON, L. E.,ANDRUDY, B. Shaker K1channel subunits from heteromultimeric channels with novel functional properties.
Biochem. Biophys. Res. Commun. 171: 1361–1371, 1990.
MCCORMICK, D. A., CONNORS, B. W., LIGHTHALL, J. W.,ANDPRINCE, D. A. Comparative electrophysiology of pyramidal and sparsely spiny stellate neurons of the neocortex. J. Neurophysiol. 54: 782– 806, 1985.
MORENO, H., KENTROS, C., BUENO, E., WEISER, M., HERNANDEZ, A., VEGA -SAENZ DEMIERA, E., PONCE, A., THORNHILL, W.,ANDRUDY, B. Thalamo-cortical projections have a K1channel that is phosphorylated and modulated by cAMP-dependent protein kinase. J. Neurosci. 15: 5486 –5501, 1995. MOUNTCASTLE, V. B., TALBOT, W. H., SAKATA, H.,ANDHYVARINEN, J. Cortical
mon-keys. Neuronal periodicity and frequency discrimination. J. Neurophysiol. 32: 452– 484, 1969.
PERNEY, T. M. AND KACZMAREK, L. K. Localization of a high threshold potassium channel in the rat cochlear nucleus. J. Comp. Neurol. 386: 178 –202, 1997.
PERNEY, T. M., MARSHALL, J., MARTIN, K. A., HOCKFIELD, S.,ANDKACZMAREK, L. K. Expression of the mRNAs for the Kv3.1 potassium channel gene in the adult and developing rat brain. J. Neurophysiol. 68: 756 –766, 1992. ROBERTSON, B., OWEN, D., STOW, J., BUTLER, C.,ANDNEWLAND, C. Novel
effects of dendrotoxin homologues on subtypes of mammalian Kv1 potas-sium channels expressed in Xenopus oocytes. FEBS Lett. 383: 26 –30, 1996. RUDY, B. Diversity and ubiquity of K channels. Neuroscience 25: 729 –749,
1988.
RUDY, B., CHOW, A., LAU, D., AMARILLO, Y.A.O., SAGANICH, M., MORENO, H., HERNANDEZ-PINEDA, R., HERNANDEZ-CRUZ, A., MACINTOSH, P., ERISIR, A., LEONARD, C. S.,AND VEGA-SAENZ DE MIERA, E. Contributions of Kv3 channels to neuronal excitability. Ann. NY Acad. Sci. 868: 304 –343, 1999. RUDY, B., KENTROS, C., WEISER, M., FRUHLING, D., SERODIO, P., VEGA-SAENZ DEMIERA, E., ELLISMAN, M. H., POLLOCK, J. A.,ANDBAKER, H. Region-specific expression of a K1channel gene in brain. Proc. Natl. Acad. Sci.
USA 89: 4603– 4607, 1992.
RUDY, B., SEN, K., VEGA-SAENZ DEMIERA, E., LAU, D., RIED, T.,ANDWARD, D. C. Cloning of a human cDNA expressing a high voltage-activating, TEA-sensitive, type-A K1 channel which maps to chromosome 1 band p21.
J. Neurosci. Res. 29: 401– 412, 1991.
SEKIRNJAK, C., MARTONE, M. E., WEISER, M., DEERINCK, T., BUENO, E., RUDY, B.,ANDELLISMAN, M. Subcellular localization of the K1channel subunit Kv3.1b in selected rat CNS neurons. Brain Res. 766: 173–187, 1997. SILLITO, A. Functional considerations on the operation of GABAergic
inhibi-tory processes in the visual cortex. In: Cerebral Cortex, edited by E. G. Jones and A. Peters. New York: Plenum, 1984, p. 91–114.
SINGER, W.AND GRAY, C. M. Visual feature integration and the temporal correlation hypothesis. Annu. Rev. Neurosci. 18: 555–586, 1995. STERIADE, M. Synchronized activities of coupled oscillators in the cerebral
cortex and thalamus at different levels of vigilance [published erratum
appears in Cereb Cortex 1997 Dec;7(8): 779]. Cereb. Cortex 7: 583– 604, 1997.
TRAUB, R. D., WHITTINGTON, M. A., STANFORD, I. M.,ANDJEFFERYS, J. G. A mechanism for generation of long-range synchronous fast oscillations in the cortex. Nature 383: 621– 624, 1996.
VEGA-SAENZ DEMIERA, E., MORENO, H., FRUHLING, D., KENTROS, C., AND
RUDY, B. Cloning of ShIII (Shaw-like) cDNAs encoding a novel high-voltage– activating, TEA-sensitive, type-A K1channel. Proc. R. Soc. Lond.
B Biol. Sci. 248: 9 –18, 1992.
VEGA-SAENZ DEMIERA, E., WEISER, M., KENTROS, C., LAU, D., MORENO, H., SERODIO, P.,AND RUDY, B. Shaw-related K1 channels in mammals. In:
Handbook of Membrane Channels, edited by C. Peracchia. New York:
Academic, 1994, p. 41–78.
VIDYASAGAR, T. R., PEI, X.,ANDVOLGUSHEV, M. Multiple mechanisms un-derlying the orientation selectivity of visual cortical neurones. Trends
Neu-rosci. 19: 272–277, 1996.
WANG, L. Y., GAN, L., FORSYTHE, I. D.,ANDKACZMAREK, L. K. Contribution of the Kv3.1 potassium channel to high-frequency firing in mouse auditory neurones. J. Physiol. (Lond.) 509: 183–194, 1998.
WEISER, M., BUENO, E., SEKIRNJAK, C., MARTONE, M. E., BAKER, H., HILLMAN, D., CHEN, S., THORNHILL, W., ELLISMAN, M.,ANDRUDY, B. The potassium channel subunit KV3.1b is localized to somatic and axonal membranes of specific populations of CNS neurons. J. Neurosci. 15: 4298 – 4314, 1995. WEISER, M., VEGA-SAENZ DEMIERA, E., KENTROS, C., MORENO, H., FRANZEN,
L., HILLMAN, D., BAKER, H., AND RUDY, B. Differential expression of Shaw-related K1channels in the rat central nervous system. J. Neurosci. 14: 949 –972, 1994.
YANG, W. P., LEVESQUE, P. C., LITTLE, W. A., CONDER, M. L., RAMAKRISHNAN, P., NEUBAUER, M. G.,ANDBLANAR, M. A. Functional expression of two KvLQT1-related potassium channels responsible for an inherited idiopathic epilepsy. J. Biol. Chem. 273: 19419 –19423, 1998.
YAROM, Y., SUGIMORI, M.,ANDLLINAS´ , R. Ionic currents and firing patterns of mammalian vagal motoneurons in vitro. Neuroscience 16: 719 –737, 1985.