0095-1137/06/$08.00
⫹
0
doi:10.1128/JCM.01712-06
Copyright © 2006, American Society for Microbiology. All Rights Reserved.
Array-Based Identification of Species of the Genera
Abiotrophia
,
Enterococcus
,
Granulicatella
, and
Streptococcus
䌤
†
Sheng Kai Tung,
1Lee Jene Teng,
2Mario Vaneechoutte,
3Hung Mo Chen,
4and Tsung Chain Chang
1*
Department of Medical Laboratory Science and Biotechnology, School of Medicine, National Cheng Kung University, Tainan,
Taiwan, Republic of China
1; School of Medical Technology, National Taiwan University College of Medicine, Taipei,
Taiwan, Republic of China
2; Department of Clinical Chemistry, Microbiology and Immunology,
Ghent University Hospital, Ghent, Belgium
3; and Division of Clinical Microbiology,
Department of Pathology, National Cheng Kung University Hospital, Tainan,
Taiwan, Republic of China
4Received 18 August 2006/Returned for modification 25 September 2006/Accepted 13 October 2006
Some species of enterococci and streptococci are difficult to differentiate by phenotypic traits. The feasibility
of using an oligonucleotide array for identification of 11 viridans group streptococci was previously established.
The aim of this study was to expand the array to identify species of
Abiotrophia
(1 species),
Enterococcus
(18
species),
Granulicatella
(3 species), and
Streptococcus
(31 species and 6 subspecies). The method consisted of
PCR amplification of the ribosomal DNA intergenic spacer (ITS) regions, followed by hybridization of the
digoxigenin-labeled PCR products to a panel of oligonucleotide probes (16- to 30-mers) immobilized on a nylon
membrane. Probes could be divided into three categories: species specific, group specific, and supplemental
probes. All probes were designed either from the ITS regions or from the 3
ⴕ
ends of the 16S rRNA genes. A
collection of 312 target strains (162 reference strains and 150 clinical isolates) and 73 nontarget strains was
identified by the array. Most clinical isolates were isolated from blood cultures or deep abscesses, and only
those strains having excellent species identification with the Rapid ID 32 STREP system (bioMe
´rieux Vitek,
Taipei, Taiwan) were used for array testing. The test sensitivity and specificity of the array were 100% (312/312)
and 98.6% (72/73), respectively. The whole procedure of array hybridization took about 8 h, starting from
isolated colonies, and the hybridization patterns could be read by the naked eye. The oligonucleotide array is
accurate for identification of the above microorganisms and could be used as a reliable alternative to
phenotypic identification methods.
Enterococci, nutritionally variant streptococci, most of
which have been allocated to the genera
Abiotrophia
and
Granulicatella
, and streptococci are gram-positive and
cata-lase-negative bacteria. Although many species of the above
genera are commensals of the human body, some of them
cause local or systemic infections, including subacute
endocar-ditis, bacteremia, meningitis, pneumonia, soft-tissue infections,
and eye infections (15, 22, 41, 49). Sherman (46) proposed a
scheme for dividing the streptococci into four categories: the
pyogenic division, the viridans division, the lactic division, and
the enterococci. No single system of phenotypic identification
suffices for the differentiation of this heterogeneous group of
organisms. Instead, classification depends on a combination of
features, including patterns of hemolysis on blood agar plates,
antigenic composition, growth characteristics, and biochemical
reactions (15, 41). Accurate identification of these bacteria
would be useful for understanding the pathogenesis of
infec-tions and the epidemiology of the increasing antibiotic
resis-tance among some of these microorganisms (50).
In clinical laboratories, phenotypic test kits, such as the
Rapid ID 32 STREP system (bioMe
´rieux Vitek, Taipei,
Tai-wan), the API 20 STREP system (bioMe
´rieux Vitek), or the
Vitek GPI card (bioMe
´rieux Vitek), are commonly used for
identification of enterococci, streptococci, and related bacteria
(16, 23, 28). The inherent problem of the culture-based
iden-tification of these microorganisms is the large number of
spe-cies relative to the limited number of biochemical traits, the
poor reproducibility of some tests (4, 41), the variability of
some traits within species (6, 24, 41), and the lack of sufficient
phenotypic data on more recently described species. The last
problem applies to species such as
Streptococcus australis
,
Streptococcus cristatus
,
Streptococcus infantarius
subspecies
in-fantarius
,
Streptococcus infantis
,
Streptococcus gallolyticus
, and
Streptococcus lutetiensis
(38, 44).
A variety of molecular methods have been developed for
identification of strains of enterococci, viridans group
strepto-cocci, and streptococci to the species level. The targets used for
molecular diagnosis include genes encoding rRNA (6, 10, 26,
27), the beta subunit of RNA polymerase (
rpoB
) (14), the
D
-alanine:
D-alanine ligase (18), the RNA subunit of RNase P
(
rnpB
) (25), the elongation factor (
tuf
) (36), the
manganese-dependent superoxide dismutase (
sodA
) (37, 38), the heat
shock proteins (
groESL
) (51), and the tRNA gene intergenic
spacer (2, 3, 12). Recently, nonhemolytic streptococci were
successfully identified by phylogenetic sequence analysis of
four housekeeping genes (
ddl
,
gdh
,
rpoB
, and
sodA
) (24).
Cor-rect identification of catalase-negative gram-positive cocci is to
some extent achievable by sequencing of the 16S rRNA gene
* Corresponding author. Mailing address: Department of Medical
Laboratory Science and Biotechnology, School of Medicine, National
Cheng Kung University, 1 University Road, Tainan 701, Taiwan,
Re-public of China. Phone: 886-6-2353535, ext. 5790. Fax: 886-6-2363956.
E-mail: [email protected].
† Supplemental material for this article may be found at http://jcm
.asm.org/.
䌤
Published ahead of print on 25 October 2006.
4414
on May 16, 2020 by guest
http://jcm.asm.org/
(6, 27). However, there are two problems with this approach.
First, the method does not allow differentiation of
Streptococ-cus mitis
,
Streptococcus oralis
, and
Streptococcus pneumoniae
because of significant sequence conservation of the 16S rRNA
genes in this group of bacteria (6, 24). Second, many sequences
in the public databases are mislabeled, either because of
in-correct identification on the source strain or because of
non-recorded revised classification of the strain subsequent to
de-position of the sequence (24).
[image:2.585.48.538.83.619.2]The 16S-23S rRNA gene intergenic spacer (ITS) has been
TABLE 1. Reference strains and clinical isolates used in this study
Species Reference strain(s)a No. of
clinical isolates
Total no. of strains
Enterococcus
E. avium BCRC 10801T, CCUG 34661, 44888 4 7
E. casseliflavus ATCC 12755, 12817, CCUG 18657T 7 10
E. cecorum ATCC 43198T
, BAA150, CCUG 38939 0 3
E. columbae CCUG 27894T 0 1
E. dispar CCUG 33309T, 37857 0 2
E. durans ATCC 19432T, CCUG 46232, 37858, 44816, LMG 16886 1 6 E. faecalis ATCC 19433T
, 27332 13 15
E. faecium ATCC 19434T, CCUG 37851, 46070 11 14
E. flavescens CCUG 30567T, 30568, 30569 0 3
E. gallinarum ATCC 49573T, CCUG 29831, 34517 6 9
E. gilvus CCUG 45553T
, LMG 13600 0 2
E. hirae ATCC 8043T, BCRC 11547, 12496 3 6
E. mundtii ATCC 43186T, LMG 12308 0 2
E. pallens CCUG 45554T 0 1
E. pseudoavium CCUG 33310T
, 44888 0 2
E. raffinosus CCUG 29292T, 37864, 37865 0 3
E. saccharolyticus CCUG 33311T, NCIMB 702609 0 2
E. villorum CCUG 45025T, LMG 19177 0 2
Streptococcus
S. agalactiae ATCC 13813T 13 14
S. alactolyticus BCRC 14738T, CCUG 41502, 41503 0 3
S. anginosus ATCC 33397T, 700231 5 7
S. australis CCUG 45919T
, 45974, 45975 0 3
S. bovis ATCC 33317T, CCUG 4214 0 2
S. canis BCRC 14753T, CCUG 27660 0 2
S. constellatus ATCC 27513T, 27823 5 7
S. cristatus CCUG 33481T
, 30424, 35233 0 3
S. downei BCRC 14752T 0 1
S. dysgalactiaesubsp.equisimilis ATCC 9542, 35666, CCUG 27483, 27479 7 11 S. equisubsp. zooepidemicus BCRC 14756T, 15414, CCUG 43890 0 3 S. gallolyticussubsp. gallolyticus ATCC 43143T
, CCUG 35224, 46101, 46667 5b
9 S. gallolyticussubsp. macedonicus CCUG 39970T, 33369, 43003 0 3 S. gallolyticussubsp. pasteurianus ATCC 43144T, CCUG 19454, 35885, 46034, 46150 7b 12
S. gordonii ATCC 35105T, 49818 1 3
S. infantariussubsp.infantarius ATCC BAA102T
, CCUG 43820, 43821, 44960, 47548 0 5
S. infantis CCUG 39817T, 39818, 39821 0 3
S. iniae BCRC 14744T, CCUG 27623 0 2
S. intermedius ATCC 27335T 3 4
S. lutetiensis CCUG 43822T
, 38926, 43823 0 3
S. mitis ATCC 49456T, 15910, 15914 5 8
S. mutans BCRC 15255, 15256, CCUG 35254 5 8
S. oralis ATCC 35037T, 9811, 15914, 55229, 700233, 700234 6 12 S. parasanguinis ATCC 15912T
, 15909 4 6
S. parauberis LMG 12174 0 1
S. pneumoniae ATCC 33400T, 6301, 27336, 49619 9 13
S. porcinus BCRC 14751T, CCUG 7982, 41363 0 3
S. pyogenes ATCC 14289T
, 12344 8 10
S. ratti CCUG 27641T, 27642, BCRC 15255, 15256 0 4
S. salivarius ATCC 7073T, 13419, 25975, CCUG 32749B 6 10
S. sanguinis ATCC 10556T, 49295, 49296 3 6
S. sobrinus BCRC 14757T
, CCUG 21019, 35254 0 3
S. suis BCRC 14757T, CCUG 42755, 42756 0 3
S. thermophilus BCRC 13869T, ATCC 15910, CCUG 35458 0 3
S. uberis ATCC 19436T, 13386, 700407 1 4
S. urinalis CCUG 41590T
, 41825 0 2
S. vestibularis CCUG 24893T, 32749 0 2
Nutritionally variant streptococci
Abiotrophia defectiva ATCC 49176T, CCUG 27805 5 7
Granulicatella
G. adiacens ATCC 49175T, CCUG 27811, 44406, 44407, 44408 7 12
G. balaenopterae CCUG 37380T 0 1
G. elegans CCUG 38949T, 13462, 26024, 27554 0 4
Total 150 312
aATCC: American Type Culture Collection, Manassas, Va.; BCRC, Bioresources Collection and Research Center, Taiwan; CCUG, Culture Collection of the University of Go¨teborg, Sweden; LMG, Laboratorium voor Microbiologie, Belgium; NCIMB, The National Collection of Industrial, Marine and Food Bacteria, United Kingdom. A total of 162 reference strains were used.
bThree biotypes (I, II.1, and II.2) are recognized by the Rapid ID 32 STREP system.S. bovisbiotypes I and II.2 were renamedS. gallolyticussubsp.gallolyticusand S. gallolyticussubsp.pasteurianus, respectively (44).
on May 16, 2020 by guest
http://jcm.asm.org/
suggested as a good candidate for bacterial identification and
strain typing (8, 19, 21, 40). The ITS region is found to have a
high degree of sequence and length variation at both the genus
and species levels (21, 30, 55). Recently, DNA array
technol-ogy was found to be a useful tool to identify (or detect) a wide
variety of microorganisms (7, 17, 29, 34, 43, 53, 54). In our
previous study, the feasibility of using an oligonucleotide array
to identify 11 species of viridans group streptococci was
estab-lished (9). This study aimed to expand the results of the array
technique to cover a more comprehensive spectrum including
53 species and 6 subspecies of
Abiotrophia
,
Enterococcus
,
Granulicatella
, and
Streptococcus
.
MATERIALS AND METHODS
Bacterial strains.A collection of 312 target strains (162 reference strains and 150 clinical isolates) was analyzed (Table 1). Reference strains were obtained from the American Type Culture Collection (ATCC) (Manassas, Va.), the Biore-sources Collection and Research Center (BCRC, Hsinchu, Taiwan), and the Culture Collection of the University of Go¨teborg (CCUG, Go¨teborg, Sweden). Clinical isolates were obtained from the National Taiwan University Hospital (Taipei, Taiwan), the National Cheng Kung University Medical Center (Tainan, Taiwan), and the Ghent University Hospital (Ghent, Belgium). Most clinical isolates were isolated from blood cultures or deep abscesses and identified to the species level with the Rapid ID 32 STREP system. To avoid ambiguous identities of clinical isolates, only those strains having excellent species identification ac-cording to the criteria of the Rapid ID 32 STREP system were used for array testing. In addition, species names of clinical isolates of viridans group strepto-cocci were verified by ITS sequence analysis as previously described (8). In addition, a total of 73 nontarget strains (belonging to 51 species other than the species covered by the DNA array) were used for a specificity test of the array (see Table S1 in the supplemental material). All strains were cultured on sheep blood agar, incubated at 35°C for 24 to 48 h, and then used for DNA extraction. Strains ofAbiotrophiaandGranulicatellawere cultured on chocolate agar.
DNA preparation.The boiling method was used to extract DNA from bacteria (31). Briefly, one to several colonies of pure cultures were suspended in 50l of sterilized water, heated at 100°C for 15 min in a heating block, and centrifuged in a microcentrofuge (6,000⫻g; 10 min). The supernatant containing bacterial DNA was stored at⫺20°C for further use.
Database of oligonucleotide probes.A total of 88 oligonucleotide probes (16-to 30-mers) (Table 2) were used (16-to construct an oligonucleotide database (16-to identify the bacteria listed in Table 1. Most probes were designed from the ITS regions, except for five that were based on the sequences of the 3⬘ends of 16S rRNA genes. Reference sequences extracted from GenBank (Table 1) were confirmed by at least one sequence of another reference strain of the same species in the database. If an ITS sequence was determined in this study and used for probe design, the sequence was also confirmed by using the ITS sequence of a second reference strain of the same species, except for a few species that had only one strain in our collection (Table 1).
Alignments of the ITS sequences of different species were performed by using the PrettyBox algorithm of the Wisconsin Genetics Computer Group package (version 10.3; Accelrys Inc., San Diego, Calif.), and areas displaying sequence divergence among species were used for probe synthesis. The designed probes were checked for internal repeats, self-biding, secondary structure, and GC content by using the software Vector NTI (Invitrogen Corporation, Carlsbad, Calif.) and screened against the databases of the National Center for Biotech-nology Information for homology with other bacterial sequences using BLAST (Basic Local Alignment Search Tool). Fifteen, ten, or five additional bases of thymine were added to the 3⬘(or 5⬘) ends of probes that hadTmvalues less than 37°C, 45°C, or 50°C, respectively (7) (Table 2). Some probes used for identifi-cation of viridans group streptococci were reported previously (9); however, many probes were newly designed or modified to identify several additional species in this group and to meet the change of hybridization stringency used in this study (Table 2).
One probe or multiple probes were designed to identify a single species, depending on the availability of divergent sequences in the ITS regions (Table 2). The 88 probes could be divided into 3 categories: species specific, group specific (i.e., a probe shared by several species), and supplemental probe (i.e., a probe used to differentiate between genetically related streptococci). Supplemental probes were either designed from the ITS regions or from the 3⬘ends of the 16S
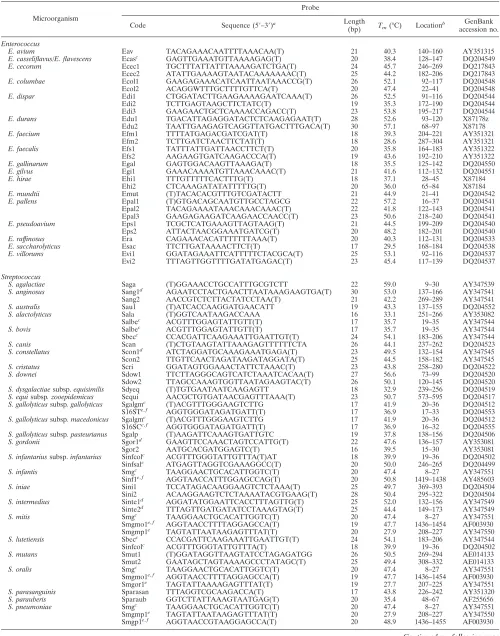
rRNA genes. A probe based on a conserved sequence (5⬘-GTCGTAACAAGG TAGCCGTA-3⬘) at the 3⬘end of the 16S rRNA gene (GenBank accession no. AB023575) was used as a positive control probe. In addition, the digoxigenin-labeled reverse primer 6R (5⬘-dig-GGGTTYCCCCRTTCRGAAAT-3⬘; Y⫽C or T, and R⫽A or G), used to amplify the ITS region, was spotted on the array and used as a position marker after hybridization (Fig. 1 and 2).
Fabrication of oligonucleotide arrays.The array (0.9 by 1.1 cm) contained 117 dots (9⫻13 dots), including 88 dots for species (or subspecies) identification, 1 dot for a positive control (probe code PC), 8 dots for negative controls (probe code NC, tracking dye only), and 20 dots (probe code M) for position markers (Fig. 1). The oligonucleotide probes were diluted 1:1 (final concentration, 10
M) with a tracking dye solution, drawn into wells of 96-well microtiter plates, and spotted onto positively charged nylon membranes (Roche, Mannheim, Ger-many) as described previously (9). The arrays were fabricated with an automatic arrayer (SR-A300; Ezspot, Taipei, Taiwan) by using a solid pin (diameter, 400
m). Probes on the array were divided by position markers into four areas. Probes in the upper left, upper right, and lower left corner were used for identification of species of nutritionally variant streptococci, enterococci, and streptococci, respectively. The lower right corner contained supplemental probes that helped group-specific probes to differentiate some genetically related strep-tococci (Fig. 1).
Amplification of the ITS regions for hybridization.The bacterium-specific universal primers 13 BF (5⬘-GTGAATACGTTCCCGGGCCT-3⬘) and 6R (5⬘ dig-GGGTTYCCCCRTTCRGAAAT3⬘) (Y⫽C or T, and R⫽A or G) (39) were used to amplify a DNA fragment that encompassed a small portion of the 16S rRNA gene, the ITS, and a small portion of the 23S rRNA gene. The reverse primer 6R was labeled with a digoxigenin molecule at its 5⬘end. PCR was carried out as described previously (9), except that digoxigen11-dUTP was not in-cluded in the PCR mixture.
Hybridization procedures.Unless otherwise indicated, the hybridization pro-cedures were carried out at room temperature in an oven with a shaking speed of 60 rpm. All reagents except buffers were included in the DIG nucleic acid detection kit (Roche). Each array was prehybridized for 2 h with 1 ml of hybrid-ization solution (5⫻SSC [1⫻SSC is 0.15 M NaCl plus 0.015 M sodium citrate], 1% [wt/vol] blocking reagent, 0.1%N-laurylsarcosine, 0.02% sodium dodecyl sulfate [SDS]) in an individual well of a 12-well cell culture plate. The digoxi-genin-labeled PCR product amplified from an isolate was heated on a 100°C heating block for 5 min and immediately cooled in an ice bath. Ten microliters of the denatured PCR product of the test organism was diluted with 0.5 ml of hybridization solution and added to each well. Hybridization was carried out at 45°C for 90 min. The array was then given three washes (5 min each) in 1 ml of washing buffer (2⫻SSC, 0.1% SDS) and one wash (1 min) in 1 ml of a second washing buffer (0.5⫻SSC, 0.1% SDS). The array was then blocked with 1% blocking solution supplied in the DIG nucleic acid detection kit (Roche), incu-bated with alkaline phosphatase-conjugated sheep antidigoxigenin antibodies and then with the substrates of alkaline phosphatase as described previously (9). The hybridized chip was air-dried, and the image of the hybridization pattern was processed by a high-resolution scanner (Powerlook 3000; Umax, Taipei, Taiwan). The hybridized spot (diameter, 400m), displaying a blue color on a white nylon membrane, could be easily recognized by the naked eye.
Identification of strains by array hybridization.A strain was identified as one of the species (or subspecies) listed in Table 1 when the probe (or all probes) specified for that species was hybridized (Table 2). Unless otherwise specified, hybridization signals produced by supplemental probes located in the lower right corner of the array were ignored. Some species were identified by their unique hybridization patterns, produced by group-specific or group-specific and supple-mental probes, as indicated in Table 2. Supplesupple-mental probes were used to differentiate several genetically related streptococci; they had no use for identi-fication of enterococci and nutritionally variant streptococci.
Discrepant analysis.When a strain produced discrepant identification by the array, the near-complete-length 16S rRNA gene of the strain was amplified by PCR and sequenced for species clarification. To amplify the near-complete-length 16S rRNA gene by PCR, the primer pair of 8FPL (5⬘-GTTTGATCCTG GCTCAG-3⬘) and 1492RPL (5⬘-GGTTACCTTGTTACGACTT-3⬘) was used (39). PCR products were purified by a PCR-M Clean Up kit (Viogene, Taipei, Taiwan) and sequenced in both directions by using the above two primers and an additional primer, 1055r (5⬘-CACGAGCTGACGACAGCCAT-3⬘) with the BigDye Terminator cycle sequencing kit (Applied Biosystems, Taipei, Taiwan) and the ABI 377 sequencing system (Applied Biosystems). The determined sequences were compared to known sequences of 16S rRNA genes in the data-bases of the National Center for Biotechnology Information using the BLASTN algorithm. The following criteria were used for identification of a strain to the genus or species level by 16S rRNA gene sequencing: (i) when the comparison of
on May 16, 2020 by guest
http://jcm.asm.org/
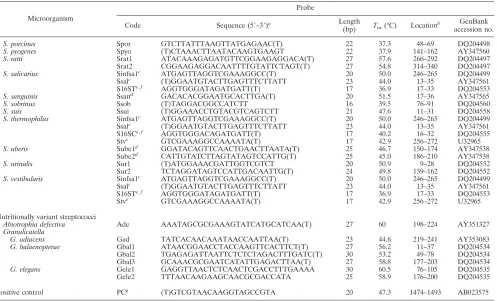
TABLE 2. Oligonucleotide probes used in this study
Microorganism
Probe
Code Sequence (5⬘–3⬘)a Length
(bp) Tm(°C) Location
b GenBank
accession no.
Enterococcus
E. avium Eav TACAGAAACAATTTTAAACAA(T) 21 40.3 140–160 AY351315 E. casseliflavus/E. flavescens Ecasc GAGTTGAAATGTTAAAAGAG(T) 20 38.4 128–147 DQ204549 E. cecorum Ecec1 TGCTTTATTATTTAAAAGATCTGA(T) 24 45.7 246–269 DQ217843
Ecec2 ATATTGAAAAGTAATACAAAAAAAC(T) 25 44.2 182–206 DQ217843
E. columbae Ecol1 GAAGAGAAACATCAATTAATAAACCG(T) 26 52.1 92–117 DQ204548
Ecol2 ACAGGWTTTGCTTTTGTTCA(T) 20 47.4 22–41 DQ204548
E. dispar Edi1 CTGGATACTTGAAGAAAAGAATCAAA(T) 26 52.5 91–116 DQ204544
Edi2 TCTTGAGTAAGCTTCTATC(T) 19 35.3 172–190 DQ204544
Edi3 GAAGAACTGCTCAAAACCAGACC(T) 23 53.8 195–217 DQ204544
E. durans Edu1 TGACATTAGAGGATACTCTCAAGAGAAT(T) 28 52.6 93–120 X87178z
Edu2 TAATTGAAGAGTCAGGTTATGACTTTGACA(T) 30 57.1 68–97 X87178
E. faecium Efm1 TTTTATGAGACGATCGAT(T) 18 39.3 204–221 AY351321
Efm2 TCTTGATCTAACTTCTAT(T) 18 28.6 287–304 AY351321
E. faecalis Efs1 TATTTATTGATTAACCTTCT(T) 20 35.8 164–183 AY351322
Efs2 AAGAAGTGATCAAGACCCA(T) 19 43.6 192–210 AY351322
E. gallinarum Egal GAGTGGACAAGTTAAAGA(T) 18 35.5 125–142 DQ204550 E. gilvus Egi1 GAAACAAAATGTTAAACAAAC(T) 21 41.6 112–132 DQ204551 E. hirae Ehi1 TTTGTTTTTCACTTTG(T) 18 37.1 28–45 X87184
Ehi2 CTCAAAGATATATTTTTG(T) 20 36.0 65–84 X87184
E. mundtii Emut (T)TACACACGTTTGTCGATACTT 21 44.9 21–41 DQ204542 E. pallens Epal1 (T)GTGACAGCAATGTTGCCTAGCG 22 57.2 16–37 DQ204541
Epal2 TACAGAAAATAAACAAACAAAC(T) 22 41.8 122–143 DQ204541
Epal3 GAAGAGAAGATCAAGAACCAACC(T) 23 50.6 218–240 DQ204541
E. pseudoavium Eps1 TCGCTCATGAAAGTTAGTAAG(T) 21 44.5 199–209 DQ204540
Eps2 ATTACTAACGGAAATGATCG(T) 20 48.2 182–201 DQ204540
E. raffinosus Era CAGAAACACATTTTTTTAAA(T) 20 40.3 112–131 DQ204533 E. saccharolyticus Esac TTCTTGATAAAACTTCT(T) 17 29.5 168–184 DQ204538 E. villorums Evi1 GGATAGAAATTCATTTTTCTACGCA(T) 25 53.1 92–116 DQ204537
Evi2 TTTAGTTGGTTTTGATATGAGAC(T) 23 45.4 117–139 DQ204537
Streptococcus
S. agalactiae Saga (T)GGAAACCTGCCATTTGCGTCTT 22 59.0 9–30 AY347539 S. anginosus Sang1d
AGAATCCTACTGAACTTAATAAAGAAGTGA(T) 30 53.0 137–166 AY347541
Sang2 AACCGTCTCTTACTATCCTAA(T) 21 42.2 269–289 AY347541
S. australis Sau1 (T)ATCACCAAGGATGAACATT 19 43.3 137–155 DQ204552 S. alactolyticus Sala (T)GGTCAATAAGACCAAA 16 33.1 251–266 AY353082
Salbec
ACGTTTGGAGTATTGTT(T) 17 35.7 19–35 AY347544
S. bovis Salbec
ACGTTTGGAGTATTGTT(T) 17 35.7 19–35 AY347544
Sbecc CCACGATTCAAGAAATTGAATTGT(T) 24 54.1 183–206 AY347544
S. canis Scan (T)CTGTAAGTATTAAAGAGTTTTTTCTA 26 44.1 237–262 DQ204523 S. constellatus Scon1d
ATCTAGGATGCAAAGAAATGAGA(T) 23 49.5 132–154 AY347545
Scon2 TTGTTCAACTAGATAAGATAGGATA(T) 25 44.5 158–182 AY347545
S. cristatus Scri GGATAGTGGAAACTATTCTAAAC(T) 23 43.8 258–280 DQ204522 S. downei Sdow1 TTCTTAGGGCAGTCATCTAAATCACAA(T) 27 56.6 73–99 DQ204520
Sdow2 TTAGCCAAAGTGGTTAATAGAAGTAC(T) 26 50.1 120–145 DQ204520
S. dysgalactiaesubsp.equisimilis Sdyeq (T)TGTGAATAATCAAGAGTT 18 32.9 239–256 DQ204519 S. equisubsp. zooepidemicus Sequi AACGCTGTGATAACGAGTTTAAA(T) 23 50.7 573–595 DQ204517 S. gallolyticussubsp. gallolyticus Sgalgmc (T)ACGTTTGGGAAGTCTTG 17 41.9 20–36 DQ204512
S16STe,f
AGGTGGGATAGATGATT(T) 17 36.9 17–33 DQ204553
S. gallolyticussubsp. macedonicus Sgalgmc
(T)ACGTTTGGGAAGTCTTG 17 41.9 20–36 DQ204512
S16SCe,f AGGTGGGATAGATGATT(T) 17 36.9 16–32 DQ204555
S. gallolyticussubsp. pasteurianus Sgalp (T)AAGATTCAAAGTGATTGTC 19 37.8 138–156 DQ204506 S. gordonii Sgor1d
GAAGTTCCAAACTAGTCCATTG(T) 22 47.6 136–157 AY353081
Sgor2 AATGCACGATGGAGTC(T) 16 39.5 15–30 AY353081
S. infantariussubsp.infantarius Sinfcolc ACGTTTGGGTATTGTTTA(T)AT 18 39.9 19–36 DQ204502
Sinfsalc ATGAGTTAGGTCGAAAGGCC(T) 20 50.0 246–265 DQ204499
S. infantis Smgc
TAAGGAACTGCACATTGGTC(T) 20 47.4 8–27 AY347551
Sinf1e,f
AGGTAACCATTTGGAGCCAG(T) 20 50.8 1419–1438 AY485603
S. iniae Sini1 TCCATAGACAAGGAAGTCTCTAAA(T) 25 49.7 369–393 DQ204504
Sini2 ACAAGGAAGTCTCTAAAATACGTGAAG(T) 28 50.4 295–322 DQ204504
S. intermedius Sinte1d
AGGATATGGAATTCACCTTTAGTTG(T) 25 52.0 132–156 AY347549
Sinte2d
TTTAGTTGATGATATCCTAAAGTAG(T) 25 44.4 149–173 AY347549
S. mitis Smgc TAAGGAACTGCACATTGGTC(T) 20 47.4 8–27 AY347551
Smgmo1e,f AGGTAACCTTTTAGGAGCCA(T) 19 47.7 1436–1454 AF003930
Smgmp1e
TAGTATTAATAAGAGTTTAT(T) 20 27.9 208–227 AY347550
S. lutetiensis Sbecc
CCACGATTCAAGAAATTGAATTGT(T) 24 54.1 183–206 AY347544
Sinfcolc ACGTTTGGGTATTGTTTA(T) 18 39.9 19–36 DQ204502
S. mutans Smut1 (T)GGATAGGTTAAGTATCCTAGAGATGG 26 50.5 269–294 AE014133
Smut2 GAATAGCTAGTAAAAGCCCTATAGC(T) 25 49.4 308–332 AE014133
S. oralis Smgc
TAAGGAACTGCACATTGGTC(T) 20 47.4 8–27 AY347551
Smgmo1e,f AGGTAACCTTTTAGGAGCCA(T) 19 47.7 1436–1454 AF003930
Smgor1e TAGTATTAAAAGAGTTTAT(T) 19 27.7 207–225 AY347551
S. parasanguinis Sparasan TTTAGGTCGCAAGACCA(T) 17 43.8 226–242 AY351320 S. parauberis Sparaub GGTCTTATTAAAGTAATGAG(T) 20 35.4 48–67 AF255656 S. pneumoniae Smgc TAAGGAACTGCACATTGGTC(T) 20 47.4 8–27 AY347551
Smgmp1e TAGTATTAATAAGAGTTTAT(T) 20 27.9 208–227 AY347550
Smgp1e,f
AGGTAACCGTAAGGAGCCA(T) 20 48.9 1436–1455 AF003930
Continued on following page
on May 16, 2020 by guest
http://jcm.asm.org/
the determined sequence with a best-scoring reference sequence of a classified species yielded an identity ofⱖ99%, the unknown isolate was assigned to that species; and (ii) when the identity was⬍99% andⱖ95%, the unknown isolate was assigned to the corresponding genus (6).
Nucleotide sequence accession numbers.Sequences of accession numbers with a prefix of “DQ” in Table 2 were determined in this study and have been submitted to GenBank.
RESULTS
Construction of oligonucleotide probe database.
A total of
88 probes (Table 2) were used to construct an oligonucleotide
database, which was then used for fabrication of the array
shown in Fig. 1. Among the 88 probes, 70 were species specific,
8 were group specific, and 10 were supplemental probes. A
group-specific probe (such as the probes Ecas and Salbe) could
hybridize with the PCR products of two or more species due to
high sequence similarities among these species (Table 2).
Sup-plemental probes were designed from the ITS or the 16S
rRNA gene regions that were coamplified by PCR during ITS
amplification and were used to differentiate genetically related
species of streptococci.
An individual species was identified by one to four probes,
depending on the availability of divergent sequences in the ITS
region. For example,
Enterococcus avium
was identified by a
single probe (Eav), while a strain was identified as
Enterococ-cus dispar
if all three probes (Edi1, Edi2, and Edi3) were
simultaneously hybridized (Table 2). Some species were
iden-tified by a combination of group-specific probes. For example,
Streptococcus bovis
was identified by its hybridization to two
group-specific probes, Salbe and Sbec (Table 2; Fig. 2).
Al-though
Streptococcus alactolyticus
also hybridized to the probe
Salbe and
S. lutetiensis
hybridized to the probe Sbec, neither
species could simultaneously hybridize with both probes.
Fur-thermore, some species were identified by their unique
pat-terns of hybridization to group-specific and supplemental
probes. For example, both
S. gallolyticus
subsp.
gallolyticus
and
S. gallolyticus
subsp.
macedonicus
hybridized with the
group-specific probe Sgalgm and were differentiated by hybridization
to two supplemental probes, S16SC and S16ST.
S. gallolyticus
subsp.
gallolyticus
produced positive hybridization with the
probe S16ST, whereas
S. gallolyticus
subsp.
macedonicus
was
positive with the probe S16SC (Table 2; Fig. 2). Another
ex-ample was
S. miti
s and
S. oralis
; both species hybridized to
the group-specific probe Smg and the supplemental probe
Smgmo1. The two species were distinguished by hybridization
of
S. mitis
to an additional supplemental probe, Smgmp1, and
[image:5.585.47.544.80.381.2]S. oralis
to another supplemental probe, Smgor1 (Table 2; Fig.
TABLE 2—
Continued
Microorganism
Probe
Code Sequence (5⬘–3⬘)a Length
(bp) Tm(°C) Location
b GenBank
accession no.
S. porcinus Spor GTCTTATTTAAGTTATGAGAAC(T) 22 37.3 48–69 DQ204498 S. pyogenes Spyo (T)CTAAACTTAATACAAGTGAAGT 22 37.9 141–162 AY347560 S. ratti Srat1 ATACAAAGAGATGTTCGGAAGAGGACA(T) 27 57.6 266–292 DQ204497
Srat2 CGGAAGAGGACAATTTTGTATTCTAGT(T) 27 54.8 314–340 DQ204497
S. salivarius Sinfsa1c
ATGAGTTAGGTCGAAAGGCC(T) 20 50.0 246–265 DQ204499
Ssalc
(T)GGAATGTACTTGAGTTTCTTATT 23 44.0 13–35 AY347561
S16STe,f AGGTGGGATAGATGATT(T) 17 36.9 17–33 DQ204553
S. sanguinis Ssand GACACACGGAATGCACTTGA(T) 20 51.5 17–36 AY347565 S. sobrinus Ssob (T)TAGGACGGCCATCTT 16 39.5 76–91 DQ204560 S. suis Ssui (T)GGAAACCTGTACGTCAGTCTT 21 47.6 11–31 DQ204558 S. thermophilus Sinfsa1c ATGAGTTAGGTCGAAAGGCC(T) 20 50.0 246–265 DQ204499
Ssalc (T)GGAATGTACTTGAGTTTCTTATT 23 44.0 13–35 AY347561
S16SCe,f
AGGTGGGACAGATGATT(T) 17 40.2 16–32 DQ204555
Stve
GTCGAAAGGCCAAAATA(T) 17 42.9 256–272 U32965
S. uberis Sube1d GGATACAGTTCAACTGAACTTAATA(T) 25 46.7 150–174 AY347538
Sube2d CATTGTATCTTAGTATAGTCCATTG(T) 25 45.0 186–210 AY347538
S. urinalis Sur1 (T)ATGGAAACGATTGGTCGTCT 20 50.9 9–28 DQ204552
Sur2 TCTAGGATAGTCCATTGACAATTG(T) 24 49.8 139–162 DQ204552
S. vestibularis Sinfsa1c ATGAGTTAGGTCGAAAGGCC(T) 20 50.0 246–265 DQ204499
Ssalc (T)GGAATGTACTTGAGTTTCTTATT 23 44.0 13–35 AY347561
S16STe,f
AGGTGGGATAGATGATT(T) 17 36.9 17–33 DQ204553
Stve
GTCGAAAGGCCAAAATA(T) 17 42.9 256–272 U32965
Nutritionally variant streptococci
Abiotrophia defectiva Ade AAATAGCGCGAAAGTATCATGCATCAA(T) 27 60 198–224 AY351327 Granulicatella
G. adiacens Gad TATCACAACAAATAACCAATTAA(T) 23 44.6 219–241 AY353083 G. balaenopterae Gbal1 ATAACGGAACCTACCAAGTTCACTTCT(T) 27 56.2 11–37 DQ204534
Gbal2 TGAGAGATTAATTCTCTCTAGACTTTGATC(T) 30 53.2 49–78 DQ204534
Gbal3 GCAAACGCGAATCATATTGAGACTTAA(T) 27 58.8 177–203 DQ204534
G. elegans Gele1 GAGGTTAACTCTCAACTCGACCTTTGAAAA 30 60.5 76–105 DQ204535
Gele2 TTTAACAAGAAGCAACGCGACCATA 25 58.9 176–200 DQ204535
Positive control PCg (T)GTCGTAACAAGGTAGCCGTA 20 47.3 1474–1493 AB023575
a
(T), additional bases of thymine were added to the 5⬘or 3⬘end of the probe. IfTmwasⱕ37°C, 15 thymine bases were added; ifTmwasⱕ45°C, 10 thymine bases were added; and ifTmwasⱖ45°C, 5 thymine bases were added to the probe.
b
The location of the probe is indicated by the nucleotide number of the ITS sequence, except where otherwise indicated. c
Group-specifics probes. d
Probe sequences already published (9). e
Supplemental probes. f
Probes designed from the 16S rRNA genes. g
The positive control probe was designed from a conserved region at the 3⬘end of the bacterial 16S rRNA gene.
on May 16, 2020 by guest
http://jcm.asm.org/
2). Species that required group-specific and supplemental
probes for identification are indicated in Table 2.
Identification of reference strains by the oligonucleotide
array.
Of 162 target reference strains tested, 154 hybridized to
their respective oligonucleotide probes and were correctly
identified. The hybridization patterns of 53 species and 6
sub-species, a majority of them being type strains, on the arrays are
alphabetically shown in Fig. 2. Reference strains of the
differ-ent taxa in the
S. bovis
complex, i.e.,
S. bovis
,
S. gallolyticus
subsp.
gallolyticus
,
S. gallolyticus
subsp.
macedonicus
, and
S.
gallolyticus
subsp.
pasteurianus
, were successfully differentiated
by the array. Species that have high sequence similarities in
their 16S rRNA genes were also accurately identified by the
present method. These species were
S. mitis
and
S.
pneu-moniae
,
Streptococcus gordonii
and
S. mitis
,
Enterococcus
du-rans
and
Enterococcus faecium
, and
Enterococcus gallinarum
and
Enterococcus casseliflavus
(27, 35).
Eight reference strains (4.9%), including one enterococcal
and seven streptococcal species, produced discrepant
identifi-cation by the array. Determination of the 16S rRNA gene
sequences of these strains showed that six of the eight strains
had been given wrong species names (Table 3).
Enterococcus
pseudoavium
CCUG 44888 was identified as
E. avium
by array
hybridization. A BLAST search revealed that the 16S rRNA
gene sequence of the strain CCUG 44888 had identities of 99.1
and 98.8% with GenBank reference sequences of
E. avium
and
E. pseudoavium
, respectively (Table 3). Therefore,
E.
pseudoa-vium
CCUG 44888 should be a strain of
E. avium
, as identified
by the array and confirmed by 16S rRNA gene sequencing.
Similarly,
Streptococcus mutans
BCRC 15255,
S. mutans
BCRC
15256,
Streptococcus sobrinus
CCUG 35254,
Streptococcus
uberis
ATCC 13386, and
Streptococcus vestibularis
CCUG
32749 were found to be misidentifications of
Streptococcus ratti
,
S. ratti
,
Streptococcus mutans
,
Streptococcus parauberis
, and
Streptococcus salivarius
, respectively, as revealed by array
hy-bridization and further confirmed by their 16S rRNA gene
sequences (Table 3).
S. cristatus
CCUG 35233 and
S. mitis
ATCC 15914 were identified only to the genus level (
Strepto-coccus
) by sequencing of the 16S rRNA genes (Table 3), since
the identities between the query sequences and the
best-scor-ing reference sequences in the public databases were only 98%.
However, a BLAST search of the ITS sequences of
S. cristatus
CCUG 35233 and
S. mitis
ATCC 15914 against GenBank
re-vealed that the both best-scoring sequences were from
S. oralis
,
with sequence identities of 99.6% and 99.1%, respectively. To
further clarify the identities of strains CCUG 35233 and ATCC
15914, the manganese-dependent superoxide dismutase genes
(
sodA
) of both strains were amplified (38) and sequenced. A
BLAST search of the
sodA
sequences of strains CCUG 35233
and ATCC 15914 revealed that the best-scoring reference
se-quences were
S. oralis
(GenBank accession no. Z99195;
se-quence identity, 96%) and
S. oralis
(GenBank accession
AB200066; sequence identity, 96%), respectively. Since the
hybridization results for strains CCUG 35233 and ATCC
15914 were supported by sequence analysis of the ITS region
and
sodA
gene, the two strains were considered to be correctly
identified as
S. oralis
by the array. Since all eight discordant
reference strains were proved to be correctly identified by
array hybridization, the test sensitivity of the array for
refer-ence strains was 100% (162/162).
Identification of clinical isolates by the oligonucleotide
ar-ray.
Of 150 target clinical isolates tested,
E. avium
2510 and
E.
durans
790 yielded discrepant identifications by array
hybrid-ization (Table 3).
E. avium
2510 and
E. durans
790 were
iden-tified as, respectively,
Enterococcus raffinosus
and
Enterococcus
hirae
by the array, and this identification was confirmed by
sequence analysis of the 16S rRNA genes. Since these two
discordant isolates were proved to be correctly identified by
hybridization, the test sensitivity of the array for clinical
lates was 100% (150/150). If reference strains and clinical
iso-lates were considered together, an overall sensitivity of 100%
(312/312) was obtained by the present method.
Hybridization of nontarget strains to the oligonucleotide
array.
Of 73 nontarget strains (51 species) tested by the array
(see Table S1 in the supplemental material), one strain (
Vibrio
alginolyticus
BCRC 12829) hybridized to the probe Saga and
was misidentified as
Streptococcus agalactiae
(Fig. 2). The
re-maining 72 strains did not produce any hybridization signals
with probes on the array, except for the positive control probe.
Therefore, the test specificity of the array was 98.6% (72/73).
DISCUSSION
In this study, an oligonucleotide array was developed to
identify 53 species and six subspecies of the genera
Abiotro-phia
,
Enterococcus
,
Granulicatella
, and
Streptococcus
. A
[image:6.585.43.286.67.298.2]sensi-tivity of 100% (312/312) and a specificity of 98.6% (72/73) were
obtained by the array. The present method used a standardized
FIG. 1. Layout of oligonucleotide probes on the array (0.9 by 1.1
cm). The probe “PC” (A9) (a positive control) was designed from a
conserved region at the 3
⬘
end of the 16S rRNA gene. Probes coded
“NC” were negative controls (tracking dye only). Probes coded “M”
were digoxigenin-labeled primer 6R and were used as position
mark-ers. Probes in the upper left, upper right, and lower left corners were
used to identify species of nutritionally variant streptococci,
entero-cocci, and streptoentero-cocci, respectively. Probes in the lower right corner
were supplemental probes. The corresponding sequences of all probes
are listed in Table 2.
on May 16, 2020 by guest
http://jcm.asm.org/
protocol encompassing DNA extraction, PCR amplification of
the ITS regions, and hybridization of the PCR products to the
array. The hybridized spot (400
m in diameter), displaying a
blue color on a white nylon membrane, could be easily
recog-nized by the naked eye. The whole procedure can be finished
within approximately 8 h, starting from isolated colonies. The
present method might be useful for conditions that necessitate
identification of enterococci, streptococci, and related bacteria
to the species (and subspecies) level.
In this study, an individual species was identified by either
one or multiple probes, depending on the availability of
diver-gent sequences in the ITS region. The advantage of using
multiple probes is the increase in specificity, since the chance
for a cross-reacting strain to hybridize to all probes designed
for a species is very low. However, the use of multiple probes
to identify a species may potentially decrease sensitivity, due to
the possibility of mutations that occur at the regions used for
probe design. The successful design of different probes,
includ-ing group-specific and supplemental probes, was based on the
known sequences in regions of the ITS and the 3
⬘
ends of 16S
rRNA genes. Multiple sequence alignment (interspecies and
intraspecies) plays an essential role in finding out the regions
that could be used for probe design.
Routine procedures based on phenotypic tests do not allow
unequivocal identification of some streptococci. Hoshino et al.
(24) examined a collection of 115 strains of nonhemolytic
streptococci isolated from bacteremic patients and 33
refer-ence strains by using 2 commercial kits (rapid ID 32 STREP
and STREPTOGRAM [Wako Pure Chemicals, Osaka,
Ja-pan]). The correct identification rates for strains by both
com-mercial kits were below 50% but varied significantly between
species. The most significant problems were observed with
S.
mitis
and
S. oralis
and 11
Streptococcus
species described since
1991. They concluded that phenotypic characterization is of
limited value for identification of many species of
nonhemo-lytic streptococci and is not a valid approach at the present
time. Recently, Bosshard et al. (6) tested 171 strains of aerobic
catalase-negative gram-positive cocci with the API 20 STREP
system and found that less than 60% of isolates could be
identified to the species or genus level. To solve the problems
of phenotypic identification, the feasibility of using an
oligo-nucleotide array for species identification was investigated
here and favorable results were obtained. The array technology
has been used to identify a spectrum of microorganisms,
in-cluding
Mycobacterium
(17, 34, 43), bacteria in positive blood
cultures (7), bacteria from cervical swab specimens (32),
on May 16, 2020 by guest
http://jcm.asm.org/
Campylobacter
(53),
Listeria
(54), and food-borne bacterial
pathogens (29).
Clinical isolates used in this study were identified by the
Rapid ID 32 STREP system. To avoid as much as possible
discrepant identification results, thus avoiding sequence
anal-ysis of the 16S rRNA gene as much as possible, only those
[image:8.585.45.283.69.425.2]strains having excellent species identification (i.e., percent
identification of
ⱖ
99.9%) according to the criteria of the ID 32
STREP system were used for array testing. Strains with good
identification (percent identification of
ⱖ
90%) or acceptable
identification (percent identification of
ⱖ
80%) were not
in-cluded in this study, since Bosshard et al. (6) observed that
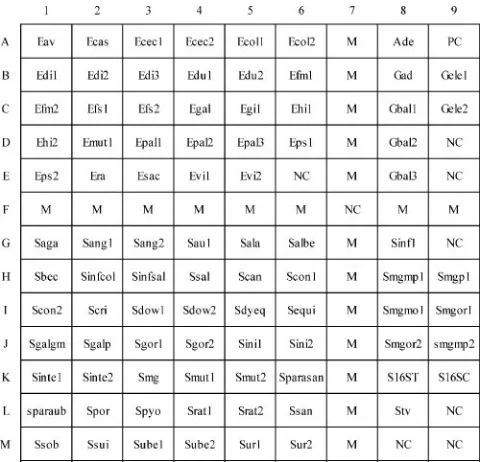
[image:8.585.42.546.557.692.2]FIG. 2. Hybridization results for species of enterococci (18
spe-cies), streptococci (31 species and 6 subspespe-cies), and nutritionally
variant streptococci (4 species). All strains, except two, were type
strains and were alphabetically arranged according to their species
names. The corresponding probes hybridized on the arrays are
indi-cated in Fig. 1, and the corresponding sequences of the hybridized
probes are shown in Table 2. The hybridized probe on the uppermost
right corner on each array was the positive control. Hybridization
signals produced by supplemental probes (located at the lower right
corner) were used to differentiate genetically related species of
strep-tococci and had no use in identification of enterococci and nutritionally
variant streptococci.
TABLE 3. Strains that produced discrepant identification by array hybridization and results of discrepant analysis
Strain Species name
received
Species identification by:
Result of discrepant analysis Array
hybridization 16S rRNA gene sequence (%) a
CCUG 44888
E. pseudoavium
E. avium
E. avium
(99.1),
E. pseudoavium
(98.8)
E. avium
CCUG 35233
S. cristatus
S. oralis
S. oralis
(98),
S. mitis
(98)
S. oralis
bATCC 15914
S. mitis
S. oralis
S. oralis
(98),
S. mitis
(98),
S. pneumoniae
(98)
S. oralis
bBCRC 15255
S. mutans
S. ratti
S. ratti
(99),
S. mutansc
(92)
S. ratti
BCRC 15256
S. mutans
S. ratti
S. ratti
(99),
S. mutans
(92)
S. ratti
CCUG 35254
S. sobrinus
S. mutans
S. mutans
(99),
S. sobrinus
(90)
S. mutans
ATCC 13386
S. uberis
S. parauberis
S. parauberis
(99),
S. uberis
(92)
S. parauberis
CCUG 32749
S. vestibularis
S. salivarius
S. salivarius
(99.7),
S. vestibularis
(99.1)
S. salivarius
2510
cE. avium
E. raffinosus
E. raffinosus
(100),
E. avium
(99)
E. raffinosus
790
cE. durans
E. hirae
E. hirae
(99),
E. durans
(98)
E. hirae
aValues in parentheses are percentages of 16S rRNA gene sequence identities of the test strains with sequences in GenBank.
bCCUG 35233 and ATCC 15914 were found to beS. oralisby sequence analysis of the ITS region (8) and the manganese-dependent superoxide dismutase gene (sodA) (38).
cClinical isolates.