comment
reviews
reports
deposited research
interactions
information
refereed research
Review
Nuclear dynamics: where genes are and how they got there
Jason R Swedlow and Angus I Lamond
Address: Division of Gene Regulation and Expression, Wellcome Trust Biocentre, University of Dundee, Dow Street, Dundee DD1 5EH, UK.
Correspondence: Jason R Swedlow. E-mail: [email protected]
Abstract
DNA is highly organized spatially, both within domains of chromatin along each chromosome and within the nucleus as a whole. Recent studies suggest that chromatin localization can affect transcriptional and replicational activity. The similarity between the movements of chromatin nuclear bodies suggests a common mechanism that regulates nuclear dynamics.
Published: 9 Month 2001
GenomeBiology2001, 2(3):reviews0002.1–0002.7
The electronic version of this article is the complete one and can be found online at http://genomebiology.com/2001/2/3/reviews/0002 © BioMed Central Ltd (Print ISSN 1465-6906; Online ISSN 1465-6914)
We have grown used to the linear representation of the genome as a cluster of coloured lines representing different chromosomes or sections thereof. Unfortunately, for those of us studying its properties, the structure of the genome in vivo is far more complicated. Each chromosome is a complex of protein and nucleic acid, assembled into higher-order chromatin structures and packed into the cell nucleus. It is this chromatin complex that is the substrate for tran-scription, replication, damage repair and recombination. But chromatin can exist in at least two forms, termed open and closed, that differ in their accessibility to nuclear factors including transcriptional activators and the replication machinery. It now appears that processes governing the opening of chromatin may play key regulatory roles in the nucleus. Recent studies have shown that loci do indeed expand upon transcriptional activation [1,2]. Moreover, protein machines and post-translational modifications have been identified that are involved in mediating structural changes in chromatin [3-5].
It is likely that nuclear architecture also influences chro-matin structure, just as cytoplasmic organelles are affected by their localization. Efforts to understand the architecture of the nucleus have exposed a remarkable level of organiza-tion of chromatin. Individual chromosomes are seen to occupy separate, non-overlapping territories in the inter-phase nucleus [6-9]. Moreover, a number of nuclear bodies
are now known, usually identified by the presence of specific molecular markers and formed in the nucleoplasm without the use of membrane barriers [10,11]. These bodies may be sites where specific nuclear activities take place; for example, the production of ribosomal subunits takes place in the nucleolus.
The nucleus is a dynamic organelle. Its partitioning into dif-ferent compartments, each formed without membrane boundaries, suggests that the nucleus may use a different organizing principle than that of much of the cytoplasm. It also implies that there is a transport system to deliver factors to their appropriate targets and to move macromolecules between compartments, in addition to the tremendous flux of macromolecules entering and leaving the nucleus through the nuclear pores. So, how do things localize and move in the nucleus? In this review, we discuss the positioning and movement of nuclear components and how these processes may impinge on critical nuclear functions.
Chromatin positioning
chromatin loci that are separated by many kilobases can nev-ertheless make contact and functionally interact. Changes in higher-order chromatin structure are important for function, although we dont yet understand the mechanisms involved.
A good example of the relationship between function and nuclear location is provided by the inactivated human X chromosome, or Barr body. In human females, the inac-tive X chromosome is maintained in a condensed, hete-rochromatic state and predominantly positioned at the periphery of the nucleus, juxtaposed to the nuclear envelope. Transcriptionally inactive heterochromatin is also often observed clustered along the nuclear envelope. Does the location of inactive heterochromatin reflect a regulated localization of chromatin to a specific subnuclear domain? It is possible that parts of chromosomes (or as in the case of the Barr body, whole chromosomes) that are repressed are localized to the periphery of the nucleus because they are simply excluded from the more interior portion. Recent evi-dence suggests, however, that the localization of chromatin loci is a regulated process with profound consequences for their activity. The key to these observations has been the combination of techniques for fluorescence in situ hybridiza-tion (FISH) that preserve the structure of the nucleus and high-resolution, multi-colour fluorescence microscopy.
The gene density in human chromosomes is known to vary widely. Bickmore and colleagues [13] have explored the loca-tion of gene-rich and gene-poor chromosomes in human cells. Using FISH techniques that paint whole somes, they have visualized the relative position of chromo-somes in the nucleus and correlated these with transcriptional activity. A statistical analysis of the positions of the relatively gene-poor human chromosome 18 and the gene-rich chromosome 19 showed that chromosome 18 was preferentially located near the nuclear periphery. After inhibiting transcription, the preferential localization of chro-mosome 18 to the nuclear periphery was no longer observed. These data suggest that chromosome location can be depen-dent upon active transcription. Most recently, the same workers have examined the disposition of all human chro-mosomes in the interphase nucleus [14], and the results reinforce the inverse correlation between proximity to the nuclear periphery and gene density.
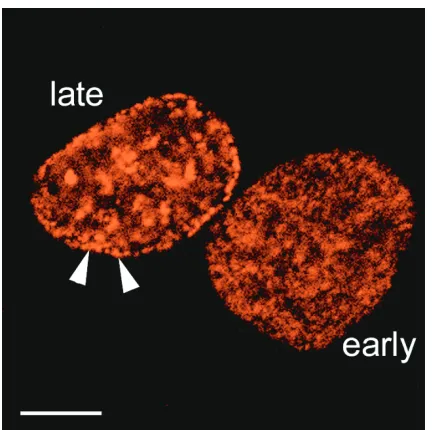
It is interesting to compare these results with previous observations showing that late-replicating chromatin is pref-erentially located at the nuclear periphery (Figure 1) and that high levels of transcription and of acetylated histone H4 are found in the nuclear interior [15,16]. These domains are stably maintained through a number of cell cycles, showing that they are a fundamental aspect of nuclear organization [9,16]. Collectively, these data suggest that in the crowded milieu of the nucleus, chromosome position may be finely tuned and dependent on transcription, replication, and possibly other nuclear activities.
If domains of heterochromatin are specifically localized to defined regions within the nucleus, this implies that both
cis-and trans-acting factors must mediate these events. It has been suggested that the nuclear periphery may be repressive by nature and may contain some factors that promote hetero-chromatin formation. Indeed, in yeast, localization of telo-meres to the nuclear periphery requires the presence of the Sir proteins, factors that mediate transcriptional silencing at the telomere [8,17]. The requirement for specific factors that mediate heterochromatin formation and localization suggests that localization is unlikely to be an entirely passive event.
Position-effect variegation
[image:2.609.328.544.84.299.2]The phenomenon of position-effect variegation (PEV) pro-vides further evidence that the localization of a chromatin locus has strong consequences for its activity, and that prox-imity to heterochromatin directly represses transcription. PEV was first observed in Drosophila, where chromosome rearrangements caused repression of a number of genes by moving them close to a large block of centric heterochro-matin [18]. A classic example involves an inversion of the X chromosome that moves the white gene to a location near a large block of centric heterochromatin. Expression from the relocated whitegene was usually repressed, but the level of repression differed between groups of clonally related
Figure 1
comment
reviews
reports
deposited research
interactions
information
refereed research
cells, generating a variegated phenotype. The overall level of repression also depended on the proximity of whiteto hete-rochromatin: inversions that placed white close to hete-rochromatin repress more than those that put whitefarther away. This cis-inactivation might occur by the spreading of transcription-silencing factors from heterochromatin into the white gene [19]. Alternatively, the proximity to hete-rochromatin of whiteon the inverted X might expose white
to a repressive environment, perhaps mediated by a cluster of heterochromatin [20].
A clue to the mechanism of PEV came from studies on the
Drosophila browngene and a trans-inactivation phenome-non. In Drosophila, homologous chromosomes pair in somatic cells, allowing the localization of one homolog to affect the other. The brown-Dominantallele (bwD) contains
a 1-2 Mb insertion of heterochromatin into the browngene [21]. In a heterozygous animal bearing one wild-type and one bwD allele (bwD/+), transcription from the wild-type
allele is repressed. In cells bearing two copies of the wild-type allele, FISH analysis showed that both loci were local-ized away from the nuclear periphery. By contrast, in bwD/+
flies, both alleles were clustered together on the nuclear periphery in the vicinity of heterochromatin [22,23]. These studies demonstrated that the location of a locus could move. Moreover, they suggest that one part of heterochro-matin-mediated repression includes the recruitment of a locus to a heterochromatin zone of the nucleus. In this model, a gene proximal to heterochromatin in linear sequence is guided to a heterochromatic repressive zone, possibly by interactions between separate blocks of hete-rochromatin [24]. Furthermore, interactions of specific loci with heterochromatin can be transient.
The studies on bwD suggest that the insertion of DNA
sequences that form heterochromatin near to a gene can influence the genes activity and location. What might these
cis-acting sequences be? The recent development of syn-thetic compounds, such as oligomers of pyrrole and imida-zole amino acids that bind to specific repetitive tracts of DNA with high affinity, has provided a biochemical tool for probing the effects of heterochromatin structure [25]. These compounds make heterochromatin more accessible to nucle-ases, presumably by making it more open, and they also show interesting effects in vivo. For example, Drosophilafed these compounds are viable, but show limited homeotic transformations. Moreover, the compounds also reverse the effects of heterochromatin silencing of both the whitegene and the brownlocus in bwD/+ flies [26]. It will be
interest-ing to determine the effects of drugs affectinterest-ing heterochro-matin on the localization of specific gene loci. It is also possible that drugs targeted to other heterochromatic regions (such as telomeres) might be developed in future.
How universal is the regulated localization of chromatin loci in interphase? Since the initial observations in Drosophila,
targeted localization of heterochromatic sequences has been observed in fission yeast [27], mice, and humans, suggesting that it is does not depend on the somatic pairing of homologs that occurs in insects but is, in fact, a general phe-nomenon. In humans, the Ikaros DNA-binding protein is required for gene regulation during the development of B and T cells in the immune system [28]. Interestingly, Ikaros localizes to pericentric heterochromatin [29]. It has been proposed that it regulates gene activity by recruiting the loci it binds to the repressive environment of pericentric hete-rochromatin [30]. Ikaros localization depends on two Zn-finger motifs, suggesting that it associates with pericentric heterochromatin by directly binding DNA [31]. Recent studies have identified a homolog of Ikaros, named Helios, that forms a heterodimer with Ikaros and also localizes to centric heterochromatin [32]. In addition, at least part of the cellular pool of Ikaros is complexed to NURD, a chromatin-remodelling complex, suggesting that Ikaros function may also involve the remodelling of chromatin [33,34].
In summary, there is suggestive evidence that directed local-ization of chromatin loci is one mechanism for regulating gene expression within the nucleus. Targeted localization can be either permanent or transient. Although it is still unclear how general this process might be, it raises a number of important questions concerning the movement of chromatin domains in the nucleus. For example, do specific chromatin loci differ in their dynamic behaviour? And how rapidly can a chromatin locus move, and what factors mediate the movement of loci?
Chromatin dynamics
Direct methods have recently been used in living cells to analyse chromatin movements. The technique of marking a chromosome with a tandemly repeated array of lacO
(transcription-factor-binding) sites and expression of a fusion protein made up of green fluorescent protein (GFP) and lacI (the cognate transcription factor) provides a way of detecting a specific chromatin locus in a live cell [35]. Once integrated into the Saccharomyces cerevisiaegenome, the position of the marked locus can be tracked over time [36,37]. A plot of the distance a locus travels as a function of time provides a measure of the type of motion, whether it is passive or active, and its characteristics.Results of this type of study showed that yeast chromatin diffuses passively with a diffusion constant D = 5 x 10-12cm2/sec within a
con-fined nuclear subvolume with a radius R = 0.3mm [36]. Treatment of the cells with sodium azide to inhibit ATP syn-thesis via respiratory electron transport caused little change in these measurements, suggesting that chromatin diffusion does not require ATP hydrolysis. A similar analysis in
R = 0.9mm) [36]. An analysis of chromatin movement in living human cells, using Cy3-dUTP-labeled DNA, found similar characteristics [38]. Taken together, these results suggest that bulk chromatin passively diffuses within the nucleus and that, in general, the volume within which it moves is quite limited. We note that these observations do not necessarily rule out the presence of specific paths that nuclear components could move along [39]; but a track-like mode of movement does not appear to be the dominant mechanism used by chromatin or other nuclear bodies so far examined (see below).
It is instructive to consider the scale of these motions rela-tive to free molecules and other nuclear components. Recent studies have used fluorescence recovery after photo-bleaching (FRAP), a technique in which fluorescence is ini-tially bleached in a spot using a focused laser beam and the subsequent recovery of fluorescence is measured as a reporter of the diffusion of unbleached molecules into the bleached zone. The nuclear diffusion constants of GFP fused with the pre-mRNA splicing factor SF2/ASF, the chromatin-binding protein HMG-17, and the nucleolar protein fibrillarin, were measured in living cells using FRAP [40]. Diffusion constants for all three proteins were found to be approximately 10-9cm2/sec, significantly faster
than that measured for chromatin. A diffusion constant of 10-9cm2/sec was also obtained for polyadenylated RNA in
living cells [41] and again, no evidence was found for energy-dependent mobility.
We have discussed (above) results indicating that repression of transcriptional activity can be correlated with the location of a chromatin locus. This highlights the importance of analysing the localization of a single locus during transcrip-tional activation. Recently, Tumbar and Belmont [42] have accomplished this in living cells using a stable cell line bearing 10-20 tandemly repeated copies of the dihydrofolate reductase gene DHFR fused to lacO and also expressing GFP-lacIor GFP-lacIfused to the acidic activation domain of VP16 (GFP-VP16-AAD). In cells expressing GFP-lacI, the
DHFR-lacOrepeat was seen localized to the nuclear periph-ery in around 50% of cells examined. In the presence of GFP-VP16-AAD, only about 30% of cells showed peripheral localization. These data suggest that transcriptional activa-tion of a peripheral locus can change its posiactiva-tion in the nucleus, at least in a proportion of cells.
Taken together, these results provide a picture of a nucleus in which no components are immobile, but where proteins and other macromolecular complexes rapidly diffuse around, and perhaps within, slower moving, yet still mobile, chromatin. It should be borne in mind, however, that methods such as FRAP assay bulk populations of molecules and so may not identify small subpopulations of molecules with significantly different properties. Thus, even although passive diffusion may be the predominant mode of movement in the nucleus,
it cannot be excluded that some factors can move by energy-dependent mechanisms.
If bulk chromatin movement is largely random and well confined, then two events are required to localize a chro-matin locus specifically into a regulatory nuclear subcom-partment. First, the locus must be liberated from a confined volume the restriction to its mobility must be removed -and second, a specificity mechanism must be available to target the locus to a specific subcompartment. In view of the large amount of data suggesting passive diffusion mecha-nisms, specificity probably comes from the binding of chro-matin-associated factors, either to one another or to other nuclear components or receptors. This may be similar to the known mechanism of nuclear protein export that involves the selective transport of protein export cargoes to the nuclear pore complex, mediated by the nuclear export receptor Crm1 [43].
When might such movements be detectable? During S phase, all chromatin is disassembled and DNA is replicated and then reassembled into chromatin. This period of disas-sembly might be associated with increased chromatin dynamics and therefore might afford a chance for chro-matin to redistribute within the nucleus. Indeed, in
Drosophilathe association of heterochromatic clusters has been shown to change during S phase [44]. In mammalian cells, analysis of the localization of an inserted late-replicat-ing heterochromatic region showed movement from the nuclear periphery to the nuclear interior that correlated with late S phase [45]. A recent careful study in budding yeast revealed that late-firing replication origins were clus-tered on the nuclear periphery in G1 [46]. As cells advanced into S phase, late-firing origins lost this constraint and moved into the nuclear interior, apparently before the actual firing of the origin [46]. These data all show that large movements of specific chromatin loci can occur during S phase. It will be interesting therefore to extend the analysis of the dynamic properties of chromatin to see whether large-scale chromatin movements play an important role at other stages of the cell cycle.
Nuclear body dynamics
treatment of cells with a-amanitin, an inhibitor of tran-scription by RNA polymerase II. This suggests that dynamic changes in speckled domains were correlated with RNA polymerase II activity, and it was proposed that splicing factors can shuttle between speckles and sites of transcrip-tion [47,48].
More recently, time-lapse movies of a stable HeLa cell line expressing GFP fused to the CB marker protein p80coilin
have revealed that these can be remarkably dynamic struc-tures. CBs traverse the nucleus and undergo both joining and splitting reactions [49]. CBs could also be detected moving to and from the periphery of nucleoli and rates of movement up to about 1mm/min were recorded. CB move-ment has also been detected in HeLa cells expressing GFP fused to the nucleolar protein fibrillarin [49,50] and in the nuclei of plant cells expressing a GFP-fused snRNP protein [51]. The rate of CB movement changes markedly over time. CBs show transitions between making small movements that are apparently confined in a local subvolume and larger movements that involve their transiting more rapidly to more distant sites elsewhere in the nucleus. This behaviour of CBs is consistent with a two-step system for nuclear mobility, where constrained mobility is first relaxed, allow-ing the bodies to diffuse until they reach a new target or confined location. CBs have been seen to be transiently associated with many different transcriptional loci, suggest-ing that one mechanism for confinsuggest-ing CB mobility may be
association with chromatin [52-56]. It is interesting to compare this putative mechanism with the proposed mode of chromatin targeting discussed above, and it will be important to test in future whether a two-step release and target model may provide a general way of organizing large-scale nuclear structures.
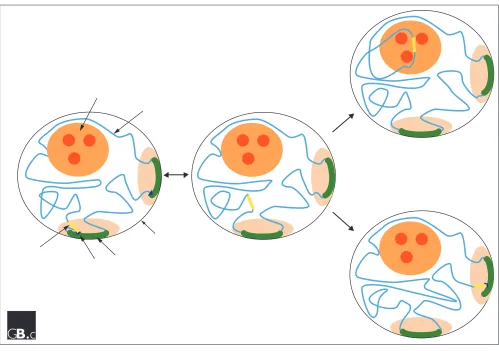
In summary, recent studies have underlined the probable importance of regulated localization of specific chromatin loci and nuclear bodies as a way of modulating their func-tion. In this review we have considered the recent data describing the dynamic behaviour of chromatin loci and nuclear bodies. We have discussed a general, two-step model that attempts to account for the movements of the large-scale nuclear structures that have been observed to date (Figure 3). A prediction of this model is that future analysis of the movements of chromatin loci and nuclear bodies will reveal marked changes either in diffusion rates, or in constrained volumes, as they undergo transitions from freely diffusing to tethered or constrained states, and vice versa. A major goal for future studies will be to understand the mechanisms involved in both moving and targeting large nuclear structures. It will be equally important to understand what mechanisms restrict their movement and tether them to specific locations. This will involve the estab-lishment of better assay systems for detecting movement and the identification and characterization of nuclear factors that mediate targeted localization.
comment
reviews
reports
deposited research
interactions
information
[image:5.609.57.554.86.337.2]refereed research
Figure 2
Chromatin and nuclear bodies. The micrograph shows the separate detection in the same HeLa cell nucleus of bulk
The further development of new and sensitive microscopy techniques to analyse the properties of fluorescently tagged fusion proteins will continue to provide insights into the dynamic properties of the nucleus in living cells. In our opinion, it will be of particular importance to focus such studies on the dynamics of single particles and specific chro-mosomal loci in living cells. Systems are now available to allow visualization of the events associated with the activa-tion of a specific locus in a living cell [1,42]. While the focus of these studies so far has been on the events associated with transcriptional activation, it will be important to look in future also at specific repression events. We anticipate that quantitative analysis of nuclear dynamics will provide a much clearer picture of how structures are localized and moved around within the nucleus, and should improve our understanding of the effects of such movement on gene expression and replication.
Acknowledgements
We thank Judith Sleeman and Barry Wong for providing Figures 1 and 2. We are grateful to Wendy Bickmore for helpful discussions. J.R.S. is a Career Development Fellow of the Wellcome Trust. A.I.L. is a Principal Fellow of the Wellcome Trust.
References
1. Tsukamoto T, Hashiguchi N, Janicki SM, Tumbar T, Belmont AS, Spector DL: Visualization of gene activity in living cells.Nat Cell Biol 2000, 2:871-878.
2. Tumbar T, Sudlow G, Belmont AS: Large-scale chromatin unfolding and remodeling induced by VP16 acidic activation domain.J Cell Biol 1999, 145:1341-1354.
3. Cheung P, Allis CD, Sassone-Corsi P: Signaling to chromatin through histone modifications.Cell 2000, 103:263-271. 4. Sudarsanam P, Winston F: The Swi/Snf family
nucleosome-remodeling complexes and transcriptional control. Trends Genet 2000, 16:345-351.
[image:6.609.57.556.83.434.2]5. Pirrotta V: Polycombing the genome: PcG, trxG, and chro-matin silencing.Cell 1998, 93:333-336.
Figure 3
6. Lichter P, Cremer T, Borden J, Manuelidis L, Ward DC: Delin-eation of individual human chromosomes in metaphase and interphase cells by in situ suppression hybridization using recombinant DNA libraries.Hum Genet 1988, 80:224-234. 7. Schardin M, Cremer T, Hager HD, Lang M: Specific staining of
human chromosomes in Chinese hamster x man hybrid cell lines demonstrates interphase chromosome territories.
Hum Genet 1985, 71:281-287.
8. Cockell M, Gasser SM: Nuclear compartments and gene regu-lation.Curr Opin Genet Dev 1999, 9:199-205.
9. Zink D, Cremer T, Saffrich R, Fischer R, Trendelenburg MF, Ansorge W, Stelzer EH: Structure and dynamics of human interphase chromosome territoriesin vivo.Hum Genet 1998, 102:241-251. 10. Lamond AI, Earnshaw WC: Structure and function in the
nucleus.Science 1998, 280:547-553.
11. Matera AG: Nuclear bodies: multifaceted subdomains of the interchromatin space.Trends Cell Biol 1999, 9:302-309.
12. Felsenfeld G: Chromatin unfolds.Cell 1996, 86:13-19.
13. Croft JA, Bridger JM, Boyle S, Perry P, Teague P, Bickmore WA: Dif-ferences in the localization and morphology of chromo-somes in the human nucleus.J Cell Biol 1999, 145:1119-1131. 14. Boyle S, Gilchrist S, Bridger JM, Mahy NL, Ellis JA, Bickmore WA:
The spatial organization of human chromosomes within the nuclei of normal and emerin-mutant cells. Hum Mol Genet 2001, 10:211-219.
15. Ferreira J, Paolella G, Ramos C, Lamond AI: Spatial organization of large-scale chromatin domains in the nucleus: a magni-fied view of single chromosome territories. J Cell Biol 1997,
139:1597-1610.
16. Sadoni N, Langer S, Fauth C, Bernardi G, Cremer T, Turner BM, Zink D: Nuclear organization of mammalian genomes. Polar chromosome territories build up functionally distinct higher order compartments.J Cell Biol 1999, 146:1211-1226.
17. Palladino F, Laroche T, Gilson E, Axelrod A, Pillus L, Gasser SM:
SIR3 and SIR4 proteins are required for the positioning and integrity of yeast telomeres.Cell 1993, 75:543-555.
18. Schultz J: Variegation in Drosophila and the inert chromo-some regions.Proc Natl Acad Sci USA 1936, 22:27-33.
19. Tartof KD, Hobbs C, Jones M: A structural basis for variegating position effects.Cell 1984, 37:869-878.
20. Henikoff S: A reconsideration of the mechanism of position effect.Genetics 1994, 138:1-5.
21. Henikoff S, Jackson JM , Talbert PB: Distance and pairing effects on the brownDominantheterochromatic element in Drosophila. Genetics 1995, 140:1007-1017.
22. Csink AK, Henikoff S: Genetic modification of heterochro-matic association and nuclear organization in Drosophila.
Nature 1996, 381:529-531.
23. Dernburg AF, Broman KW, Fung JC, Marshall WF, Philips J, Agard DA and Sedat JW: Perturbation of nuclear architecture by long-distance chromosome interactions.Cell 1996, 85:745-759. 24. Talbert PB, Henikoff S: A reexamination of spreading of posi-tion-effect variegation in the white-roughest region of
Drosophila melanogaster.Genetics 2000, 154:259-272.
25. Janssen S, Durussel T, Laemmli UK: Chromatin opening of DNA satellites by targeted sequence-specific drugs.Mol Cell 2000,
6:999-1011.
26. Janssen S, Cuvier O, Muller M, Laemmli UK: Specific gain- and loss-of-function phenotypes induced by satellite-specific DNA-binding drugs fed to Drosophila melanogaster.Mol Cell 2000, 6:1013-1024.
27. Grunstein M: Yeast heterochromatin: regulation of its assem-bly and inheritance by histones.Cell 1998, 93:325-328. 28. O’Riordan M, Grosschedl R: Transcriptional regulation of early
B-lymphocyte differentiation.Immunol Rev 2000, 175:94-103. 29. Brown KE, Guest SS, Smale ST, Hahm K, Merkenschlager M, Fisher
AG: Association of transcriptionally silent genes with Ikaros complexes at centromeric heterochromatin. Cell 1997,
91:845-854.
30. Brown KE, Baxter J, Graf D, Merkenschlager M, Fisher AG:
Dynamic repositioning of genes in the nucleus of lympho-cytes preparing for cell division.Mol Cell 1999, 3:207-217. 31. Cobb BS, Morales-Alcelay S, Kleiger G, Brown KE, Fisher AG, Smale
ST: Targeting of Ikaros to pericentromeric heterochromatin by direct DNA binding.Genes Dev 2000, 14:2146-2160.
32. Hahm K, Cobb BS, McCarty AS, Brown KE, Klug CA, Lee R, Akashi K, Weissman IL, Fisher AG, Smale ST: Helios, a T cell-restricted
Ikaros family member that quantitatively associates with Ikaros at centromeric heterochromatin. Genes Dev 1998,
12:782-796.
33. O’Neill DW, Schoetz SS, Lopez RA, Castle M, Rabinowitz L, Shor E, Krawchuk D, Goll MG, Renz M, Seelig HP, et al: An ikaros-con-taining chromatin-remodeling complex in adult-type ery-throid cells.Mol Cell Biol 2000, 20:7572-7582.
34. Kim J, Sif S, Jones B, Jackson A, Koipally J, Heller E, Winandy S, Viel A, Sawyer A, Ikeda T, Kingston R, Georgopoulos K: Ikaros DNA-binding proteins direct formation of chromatin remodeling complexes in lymphocytes.Immunity 1999, 10:345-355. 35. Belmont AS, Straight AF: In vivovisualization of chromosomes
using lac operator-repressor binding. Trends Cell Biol 1998,
8:121-124.
36. Marshall WF, Straight A, Marko JF, Swedlow J, Dernburg A, Belmont A, Murray AW, Agard DA, Sedat JW: Interphase chromosomes undergo constrained diffusional motion in living cells. Curr Biol 1997, 7:930-939.
37. Zink D, Cremer T: Cell nucleus: chromosome dynamics in nuclei of living cells.Curr Biol 1998, 8:R321-324.
38. Bornfleth H, Edelmann P, Zink D, Cremer T, Cremer C: Quantita-tive motion analysis of subchromosomal foci in living cells using four-dimensional microscopy. Biophys J 1999, 77 :2871-2886.
39. Meier UT, Blobel G: Nopp140 shuttles on tracks between nucleolus and cytoplasm.Cell 1992, 70:127-138.
40. Phair RD, Misteli T: High mobility of proteins in the mam-malian cell nucleus.Nature 2000, 404:604-609.
41. Politz JC, Tuft RA, Pederson T, Singer RH: Movement of nuclear poly(A) RNA throughout the interchromatin space in living cells.Curr Biol 1999, 9:285-291.
42. Tumbar T, Belmont AS: Interphase movements of a DNA chromosome region modulated by VP16 transcriptional activator.Nat Cell Biol 2001, 3:134-139.
43. Gorlich D, Kutay U: Transport between the cell nucleus and the cytoplasm. Annu Rev Cell Dev Biol 1999, 15:607-660.
44. Csink AK, Henikoff S: Large-scale chromosomal movements during interphase progression in Drosophila. J Cell Biol 1998,
143:13-22.
45. Li G, Sudlow G, Belmont AS: Interphase cell cycle dynamics of a late-replicating, heterochromatic homogeneously staining region: precise choreography of condensation/decondensa-tion and nuclear posicondensation/decondensa-tioning.J Cell Biol 1998, 140:975-989. 46. Heun P, Laroche T, Raghuraman MK, Gasser SM: The positioning
and dynamics of origins of replication in the budding yeast nucleus.J Cell Biol 2001, 152:385-400.
47. Misteli T, Caceres JF, Spector DL: The dynamics of a pre-mRNA splicing factor in living cells.Nature 1997, 387:523-527. 48. Eils R, Gerlich D, Tvarusko W, Spector DL, Misteli T: Quantitative
imaging of pre-mRNA splicing factors in living cells.Mol Biol Cell 2000, 11:413-418.
49. Platani M, Goldberg I, Swedlow JR, Lamond AI: In vivoanalysis of Cajal body movement, separation and joining in live human cells.J Cell Biol 2000, 151:1561-1574.
50. Snaar S, Wiesmeijer K, Jochemsen AG, Tanke HJ, Dirks RW: Muta-tional analysis of fibrillarin and its mobility in living human cells.J Cell Biol 2000, 151:653-662.
51. Boudonck K, Dolan L, Shaw PJ: The movement of coiled bodies visualized in living plant cells by the green fluorescent protein.Mol Biol Cell 1999, 10:2297-2307.
52. Smith KP, Lawrence JB: Interactions of U2 gene loci and their nuclear transcripts with Cajal (coiled) bodies: evidence for PreU2 within cajal bodies.Mol Biol Cell 2000, 11:2987-2998. 53. Smith KP, Carter KC, Johnson CV, Lawrence JB: U2 and U1
snRNA gene loci associate with coiled bodies.J Cell Biochem 1995, 59:473-485.
54. Jacobs EY, Frey MR, Wu W, Ingledue TC, Gebuhr TC, Gao L, Mar-zluff WF, Matera AG: Coiled bodies preferentially associate with U4, U11, and U12 small nuclear RNA genes in inter-phase HeLa cells but not with U6 and U7 genes. Mol Biol Cell 1999, 10:1653-1663.
55. Frey MR, Bailey AD, Weiner AM, Matera AG: Association of snRNA genes with coiled bodies is mediated by nascent snRNA transcripts.Curr Biol 1999, 9:126-135.
56. Frey MR, Matera AG: Coiled bodies contain U7 small nuclear RNA and associate with specific DNA sequences in inter-phase human cells.Proc Natl Acad Sci USA 1995, 92:5915-5919.