0095-1137/96/$04.00
1
0
Copyright
q
1996, American Society for Microbiology
Cloning and Sequencing of a Region of
Vibrio cholerae
O139
Bengal and Its Use in PCR-Based Detection
SUSANNA FALKLIND,
1,2MALIN STARK,
2M. JOHN ALBERT,
3MATHIAS UHLE
´ N,
2JOAKIM LUNDEBERG,
2*
ANDANDREJ WEINTRAUB
1Karolinska Institute, Department of Immunology, Microbiology, Pathology and Infectious Diseases, Division of Clinical
Bacteriology, Huddinge University Hospital, S-141 86 Huddinge,
1and Department of Biochemistry and Biotechnology,
KTH, Royal Institute of Technology, S-100 44 Stockholm,
2Sweden, and International Centre for Diarrhoeal Diseases
Research, Bangladesh, Dhaka, Bangladesh
3Received 7 June 1996/Returned for modification 12 August 1996/Accepted 13 September 1996
We isolated and characterized a
Vibrio cholerae
O139 Bengal-specific DNA region by arbitrary PCR. The
fragment contains open reading frames encoding two potential glycosyltransferases possibly involved in
capsular polysaccharide or lipopolysaccharide biosynthesis. In order to evaluate the possibility that this region
could be used for the specific detection of
V. cholerae
O139 Bengal, a PCR system was established. The
specificity and sensitivity of the PCR were investigated by analyzing 240 strains within the family
Vibrionaceae
and 178 strains of other gram-negative bacteria. All
V. cholerae
O139 Bengal strains tested were positive, and
none of the 384 control strains were amplified. The sensitivity of the assay was 10
2CFU/ml.
In October 1992, an epidemic of cholera-like disease due to
a
Vibrio cholerae
non-O1 serogroup broke out in the southern
Indian port city of Madras. In December of that year, there
was an outbreak of cholera-like disease in the southern coastal
area of Bangladesh, which during the subsequent months
spread to the entire country (1, 4, 5). The epidemic strain was
not related to the then known 138 serogroups of
V. cholerae
(serogroup O1 and 137 non-O1 serogroups); therefore, a new
serogroup, O139, was assigned to the strain along with the
synonym Bengal, to indicate its first isolation from the coastal
areas of the bay of Bengal (28).
V. cholerae
O139 Bengal is closely related to
V. cholerae
O1
El Tor strains associated with the seventh pandemic, and it
causes a disease which is virtually indistinguishable from
chol-era caused by
V. cholerae
O1.
V. cholerae
O139 Bengal and
V.
cholerae
O1 El Tor share several phenotypic and genotypic
properties (for a review, see reference 2). However,
V. cholerae
O139 Bengal does not produce the O1 lipopolysaccharide and
lacks all the
rfb
genes necessary for production of the O1
antigen (21).
Unlike
V. cholerae
O1, but similar to some non-O1
sero-groups,
V. cholerae
O139 Bengal possesses a polysaccharide
capsule which may confer increased virulence (14, 34). The
capsular polysaccharide is made up of hexasaccharide
repeat-ing units. The sugar composition includes colitose, galactose,
glucosamine, quinovosamine, galacturonic acid, and a cyclic
phosphate (16). The rare sugar colitose, which is a
stereoiso-mer of abequose, has only been identified in some serogroups
of
Salmonella
,
Escherichia coli
, and
Yersinia
(8, 9, 15, 17, 18).
The biochemical pathway of colitose is not completely known,
but another 3,6-dideoxyhexose, tyvelose, is present in certain
Salmonella
serogroups (10). The only structural difference
be-tween colitose and tyvelose is the orientation of the 6-deoxy
group. Theoretically, tyvelose could be transformed into
coli-tose by an epimerase. The biochemical pathways of abequose,
paratose, and tyvelose are described, and the sequences of the
rfbJ
,
rfbS
, and
rfbE
genes that are involved are known (33, 35).
To investigate whether
V. cholerae
O139 Bengal contains an
analogous
rfbS
gene, arbitrary PCR with primers
complemen-tary to the
Salmonella rfbS
gene was performed. In this study,
a
V. cholerae
O139 Bengal-specific genomic region was
iso-lated, characterized, and evaluated for PCR-based detection.
MATERIALS AND METHODS
Bacterial strains.V. choleraeO139 Bengal, strain AI-1838, was isolated at the Microbiology Laboratory of the International Centre for Diarrhoeal Disease Research, Bangladesh (ICDDR,B), Dhaka, and was used as a reference strain. The various bacterial strains used to test the specificity of the PCR are listed in Table 1. All strains were clinical isolates.V. choleraeserotypes O2 to O138 and O140 to O155 were kindly supplied by T. Shimada, National Institute of Health, Tokyo, Japan.Salmonella enteritidisSH1262,Salmonella typhimuriumSH4809,
Salmonella greensideIS842, andE. coliO55 were from the strain collection at the Division of Clinical Bacteriology, Karolinska Institute, Huddinge University Hospital, and were used as controls in the PCR.
Preparation of DNA for PCR.Fresh bacteria were suspended in phosphate-buffered saline buffer (10 mM phosphate buffer [pH 7.2], 140 mM NaCl) to an optical density (595 nm) of 0.6, corresponding to approximately 109bacteria per ml, and were pelleted by centrifugation at 10,0003gfor 5 min. The supernatant was discarded and the pellet was resuspended in 500ml of sterile distilled water. The bacteria were lysed by heating for 10 min at 1008C. After centrifugation, the supernatant containing the bacterial lysate was stored at2208C until it was used. For sensitivity determination, the experimental template was serially diluted (1:10) in sterile distilled water. Quantification ofV. choleraeO139 Bengal bac-teria was performed by determining the viable counts.
In vitro amplification.Arbitrary PCR with the DO-1 and DO-2 (Table 2) primers complementary to theSalmonella rfbSgene was performed as described by Luk et al. (19). The PCR amplification withV. choleraeO139 Bengal-specific primers was performed with 10ml of experimental template and 5 pmol of each primer, O139-1 and O139-2 (Table 2), to a total volume of 50ml of reaction buffer containing 10 mM Tris-HCl (pH 8.3), 50 mM KCl, 0.2 mM (each) de-oxynucleoside triphosphate, 2.5 mM MgCl2, and 1 U of AmpliTaq DNA poly-merase (Perkin-Elmer, Norwalk, Conn.). The PCR program used was an initial denaturation at 948C for 5 min and then a cycling procedure comprising dena-turation at 948C for 30 s, annealing at 508C for 30 s, and extension at 728C for 1 min, which was repeated for 30 cycles, and a final extension at 728C for 10 min. Nested PCR amplification was performed with O139-1 and O139-2 as the inner primer pair and O139-3 and O139-4 (Table 2) as the outer primer pair. Five microliters from the resulting PCR with the outer primer pair was used as the template in the PCR with the inner primer pair. The reaction mixture and the PCR program used were the same as those used for the PCR with the inner primers, but the number of cycles was 35 in the amplifications with both the outer and inner primers. The amplifications were carried out on a 2400 or 9600 GeneAmp PCR System (Perkin-Elmer), and the amplified products of all PCRs were analyzed by 1% agarose gel electrophoresis.
Cloning.The PCR products obtained with theSalmonella rfbSprimers were
* Corresponding author. Mailing address: Department of
Biochem-istry and Biotechnology, KTH, Royal Institute of Technology, 100 44
Stockholm, Sweden. Phone: 46-8-790 87 58. Fax: 46-8-24 54 52.
Elec-tronic mail address: [email protected].
2904
on May 15, 2020 by guest
http://jcm.asm.org/
purified by using Geneclean (Bio 101, Inc., La Jolla, Calif.), and subsequent cloning was performed with the TA cloning kit from Invitrogen (San Diego, Calif.). Plasmids were transformed intoE. coliRR1DM15 (26). Colonies were
screened by PCR, and purification of plasmids containing an insert was per-formed as described by Maniatis et al. (20).
DNA sequencing.A solid-phase DNA sequencing strategy (12) was used to determine the DNA sequence of the cloned inserts as described previously (11). Briefly, PCR was performed with 5 to 10 ng of purified plasmid and 5 pmol of each primer, RIT28 and RIT29 (biotinylated) (Table 2). A two-step PCR pro-gram was performed for 30 cycles. After PCR, the biotinylated DNA fragments were immobilized onto paramagnetic beads with covalently coupled streptavidin (Dynabeads M280-streptavidin; Dynal AS, Oslo, Norway) and were sequenced by using primers USP and RSP (Table 2) (11, 12). The sequencing products were loaded onto a 6% polyacrylamide gel, and the separation was monitored on-line with an automated laser fluorescent DNA sequencer (A.L.F.; Pharmacia Bio-tech, Uppsala, Sweden).
Restriction fragment length polymorphism.Ten micrograms of genomic DNA fromV. choleraeO139 Bengal,V. choleraeO1,E. coliO55,S. enteritidis, andS.
typhimuriumwas digested with two restriction enzymes,TaqI andHindII, and the resulting fragments were separated by electrophoresis through a 0.7% agarose gel at 20 V for 10 h. Southern blot analysis was performed as described previously (20, 29). To generate a probe,Salmonella rfbSprimers were used for PCR amplification ofS. enteritidisSH1262. The PCR product was purified with the Magic PCR Preps DNA Purification System (Promega, Madison, Wis.) and was labeled with 50 mCi of [32
P]dCTP by using an oligolabeling kit (Pharmacia Biotech).
RESULTS
PCR amplification with
Salmonella rfbS
primers.
The
cap-sular polysaccharide in
V. cholerae
O139 Bengal contains
coli-tose, a sugar not present in
V. cholerae
O1. The biochemical
pathway of colitose is not known, but theoretically, it could be
formed from tyvelose by an epimerase. The
rfbS
gene product
has been described to be involved in the pathway of tyvelose in
S. enteritidis
. We therefore investigated, by using
rfbS
-specific
primers and low-stringency PCR conditions, the presence of an
rfbS
-related gene in
V. cholerae
O139 Bengal that could be
linked to colitose. Several controls were included,
S. enteritidis
SH1262 as an
rfbS
gene-positive control,
E. coli
O55 as
coli-tose-containing control, and
V. cholerae
O1 as a negative
trol. After PCR, 1% agarose gel electrophoresis was
con-ducted. For
S. enteritidis
SH1262,
E. coli
O55, and
V. cholerae
O139 Bengal, a fragment of the expected length (
;
720 bp) was
obtained, but for
V. cholerae
O1 a fragment of
;
1,700 bp was
present (Fig. 1).
Cloning and DNA sequencing.
In order to examine if the
Salmonella rfbS
primers amplified the correct gene, the PCR
products were cloned, sequenced, and compared with the
pub-lished sequence (33). Five clones of each bacterial strain were
sequenced. The sequence of
S. enteritidis
SH1262 that was
obtained was identical to the published one. However, the
FIG. 1. Results from agarose gel electrophoresis after PCR amplification withSalmonella rfbSprimers. Lanes: 1,S. enteritidis; 2,E. coliO55; 3,V. choleraeO139 Bengal; 4,V. choleraeO1. BacteriophagelDNA restricted withPstI was used as a marker (lane M). The bottom arrow indicates the expectedS. enteritidis
fragment length of 720 bp. The top arrow indicates the approximately 1,700-bp
V. choleraeO1 fragment.
Vibrio mimicus
2
0
Aeromonas trota
1
0
Aeromonas sobria
1
0
Aeromonas hydrophila
2
0
Aeromonas caviae
2
0
Other gram-negative
organisms
Salmonella
spp.
b40
0
Shigella dysenteriae
c27
0
Shigella flexneri
d30
0
Shigella sonnei
2
0
Shigella boydii
5
0
Haemophilus influenzae
22
0
Escherichia coli
20
0
Neisseria gonorrhoeae
10
0
Yersinia enterocolitica
10
0
Klebsiella
spp.
5
0
Pseudomonas aeruginosa
3
0
Enterobacter
spp.
2
0
Campylobacter jejuni
2
0
a
Serotypes O2 to O138 and O140 to O155 (one strain each).
b
Serotypes A, B, C1, C2-3, D, E1, and E4.
c
Serotypes 1, 2, 3, 4, 7, and 9.
d
[image:2.612.58.298.91.364.2]Serotypes 1a, 1b, 1c, 2a, 2b, 3a, 3b, 4a, 4b, 5, 6, X, and Y.
TABLE 2. PCR primers used in the study
Primer Target Sequence
DO-1
Salmonella rfbS
5
9
-
TCA CGA CTT ACA TCC TAC
-3
9
DO-2
Salmonella rfbS
5
9
-
CTG CTA TAT CAG CAC ACC
-3
9
O139-1
V. cholerae
O139
5
9
-
GCG TTA TAG GTA TCA TCA AGA GA
-3
9
O139-2
V. cholerae
O139
5
9
-
GTC ATT ATT AAA ACT GCT CCA TT
-3
9
O139-3
V. cholerae
O139
5
9
-
TCG AAT TTT CAA AAT ATA CAC TT
-3
9
O139-4
V. cholerae
O139
5
9
-
CAA ACA TCT TAC AAT AGA GTA GT
-3
9
RIT-28
PCR for sequencing
5
9
-
AAA GGG GGA TGT GCT GCA AGG CG
-3
9
RIT-29
PCR for sequencing
5
9
-
Biotin-GCT TCC GGC TCG TAT GTT GTG TG
-3
9
USP
Sequencing
5
9
-
FITC
a-CGT TGT AAA ACG ACG GCC AG
-3
9
RSP
Sequencing
5
9
-
FITC-TTC ACA CAG GAA ACA GCT ATG ACC
-3
9
aFITC, fluorescein isothiocyanate.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:2.612.62.554.605.717.2]sequences of
V. cholerae
O139 Bengal,
V. cholerae
O1, and
E.
coli
O55 did not show any similarity to that of the
rfbS
gene or
to each other (data not shown). Detailed analysis of the
V.
cholerae
O139 Bengal DNA sequence revealed segments of
two open reading frames of 547 and 144 bp, respectively (Fig.
2). An alignment corresponding to the amino acid sequence
showed a significant similarity to glycosyltransferases from
other bacterial species (Fig. 3).
Restriction fragment length polymorphism.
In order to
re-solve these ambiguous observations, a restriction fragment
length polymorphism analysis was performed to investigate
whether the
rfbS
gene is present in
V. cholerae
O139 Bengal.
Previous biochemical analyses have shown that
V. cholerae
O139 Bengal,
E. coli
O55,
S. greenside
, and
S. enteritidis
all
contain colitose, while
V. cholerae
O1 and
S. typhimurium
do
not. Therefore, DNA from these bacteria was prepared and
restricted with
Taq
I and
Hin
dII and was used in the Southern
blot analysis. The probe was generated by PCR of
S. enteritidis
with the
rfbS
primers. The results suggested that the
Salmonella
rfbS
gene was present in
S. enteritidis
SH1262 only and not in
any of the other strains tested (data not shown).
PCR amplification with
V. cholerae
O139 Bengal primers.
Since the sequence of the apparent specific
V. cholerae
O139
Bengal fragment did not show any similarity to that of
V.
cholerae
O1, it could be unique for the O139 serogroup and
could therefore be used for diagnostic purposes. In order to
investigate this further, a pair of oligonucleotide primers
com-plementary to the new
V. cholerae
O139 Bengal sequence were
synthesized: O139-1 at positions 136 to 158 and O139-2 at
positions 532 to 554 in Fig. 2. After amplification of
V. cholerae
O139 Bengal DNA by PCR, 1% agarose gel electrophoresis
showed that the fragment was of the expected length, 419 bp
(Fig. 4).
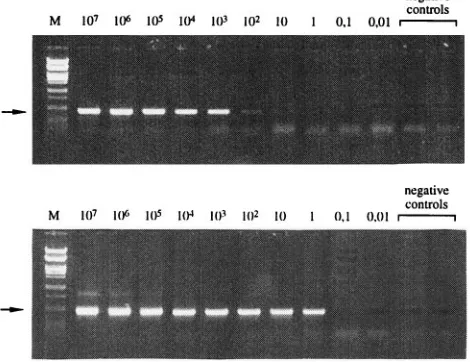
Sensitivity of nested PCR for
V. cholerae
O139 Bengal.
To
determine the detection limit of the PCR, a dilution
experi-ment was performed. With the single primer pair O139-1 and
O139-2, the detection limit was at least 10
3CFU per assay,
corresponding to 10
5CFU/ml of lysate. To increase the
sensi-tivity, a nested PCR was designed with an additional outer
primer pair: O139-3 at positions 87 to 109 and O139-4 at
positions 593 to 615 (Fig. 2). After nested PCR, the last
dilu-tion step amplified corresponded to approximately 1 CFU per
assay (10
2CFU/ml of lysate). The results of gel electrophoresis
analysis after PCR with O139-1 and O139-2 and after nested
PCR are presented in Fig. 4.
Specificity of the PCR amplification.
To examine the
spec-ificity of the oligonucleotide primer pair O139-1 and O139-2, a
total of 240 strains from the family
Vibrionaceae
were tested:
34 strains of serotype O139, 34 strains of serotype O1, 153
strains of the non-O1, non-O139 serotype, and 19 strains of
other species in the family
Vibrionaceae
(
Vibrio
parahemolyti-cus
,
Vibrio fluvialis
,
Vibrio mimicus
,
Aeromonas trota
,
Aeromo-FIG. 2. DNA sequence of theV. choleraeO139 Bengal PCR fragment
ob-tained withSalmonella rfbSprimers. The two open reading frames (open reading frames 1 and 2) are marked, and the fourV. choleraeO139 Bengal primers are indicated by arrows.
[image:3.612.318.553.77.258.2]FIG. 3. Deduced amino acid sequences of the two open reading frames (only a part of open reading frame 1) are aligned with some of the similar protein sequences found in databases by using BLAST software.1, a similar amino acid as defined by the software. The first amino acids correspond to nucleotides 2 to 4 (valine in open reading frame 1) and 550 to 552 (methionine in open reading frame 2) in Fig. 2. Sequences B, C, D, E, and F are identified as glycosyltransferases, sequence G is identified as a 40.5-kDa hypothetical protein, and sequence I is identified as a galactosyltransferase. The accession numbers of the sequences in GenBank are as follows: A, U72485; B, L46209; C, U14554; D, U15992; E, U14554; F, U25839; H, U46859; I, S73325. The accession number for sequence G in the PIR database is S47836. FIG. 4. Sensitivity of PCR assay for the detection ofV. choleraeO139 Bengal by a single PCR (A) and with nested primers (B) evaluated with a serially diluted bacterial lysate, in numbers of CFU per assay. The arrow indicates the expected length of 419 bp. Lane M, aPstI-restricted bacteriophagelDNA marker.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:3.612.141.472.572.684.2]The occurrence of epidemics of cholera due to a
V. cholerae
non-O1 isolate (
V. cholerae
O139 Bengal) marked a turning
point in the history of cholera. Until then, agglutination of
vibrios in O1 antiserum served as a serological marker for the
recognition of cholera. The disease caused by
V. cholerae
O139
Bengal is indistinguishable from traditional cholera, but
immu-nity to
V. cholerae
O1 does not protect against
V. cholerae
O139
Bengal infection (2, 3).
V. cholerae
O139 Bengal has shown a
great ability to survive in the environment and to spread
rap-idly. It is of epidemiological interest to be able to diagnose
V.
cholerae
O139 Bengal as the etiological agent of cholera.
V.
cholerae
O139 Bengal is closely related to
V. cholerae
O1, and
it is believed that this new serogroup is a mutant of
V. cholerae
O1 El Tor (22). It is of great interest to elucidate the genetic
and biological differences between the O139 and O1
groups (7, 31). It is not possible to distinguish the O139
sero-group from the O1 serosero-group by standard biochemical
meth-ods. For the detection of
V. cholerae
O139 Bengal, specific
monoclonal antibodies have been produced (24), and a new
diagnostic kit (Bengal SMART) for the rapid detection of this
species was recently described (25). When evaluated, this kit
was able to detect 5
3
10
5CFU/ml, and the specificity was 97%
in comparison with the results of culture. A DNA-based
de-tection method has also been described (23). Those
investiga-tors used two probes in a colony hybridization approach. The
specificity of this assay was very good. However, the colony
hybridization method is both laborious and time-consuming. In
this study we have investigated the possibility of detecting
V.
cholerae
O139 Bengal using specific PCR primers. The original
hypothesis was that colitose, a sugar present in the capsular
polysaccharide of
V. cholerae
O139 Bengal but absent from
V.
cholerae
O1, is synthesized by the same pathway as paratose
and tyvelose in
Salmonella
spp. Primers complementary to the
rfbS
gene encoding an enzyme involved in the biosynthesis of
tyvelose in
Salmonella
spp. were therefore tested with
V.
chol-erae
O139 Bengal and resulted in a PCR fragment of the
expected length (
;
720 bp). The fragment was isolated and
cloned. Interestingly, the nucleic acid sequence of this
frag-ment showed no similarity to the published
rfbS
gene sequence
in
Salmonella
spp. (33). It is also different from the 13-kb
V.
cholerae
O139 Bengal sequence recently published by
Com-stock et al. (6). In addition, we could confirm by restriction
fragment length polymorphism analysis that the
Salmonella
rfbS
gene was not present in
V. cholerae
O139 Bengal. The
sequence obtained from PCR with
V. cholerae
O139 Bengal
was compared with the sequences in the DNA and protein
sequence databases by using the BLAST software. Open
read-ing frames 1 and 2 showed similarities to the
glycosyltrans-ferases present in different bacterial species. In addition, open
reading frame 2 was similar to a hypothetical protein in
E. coli
(Fig. 3). It is possible that the sequence described here encodes
glycosyltransferases which might be involved in the synthesis of
the capsular polysaccharide or the lipopolysaccharide.
Although the original hypothesis has been proven wrong, the
initial experiments indicated that the isolated region was
spe-cific for
V. cholerae
O139 Bengal and could therefore be used
shown that a single
V. cholerae
O139 Bengal bacterium can be
detected by nested PCR. In addition, this step can easily be
integrated with an initial immunomagnetic separation step,
since this approach has been successful in the enrichment of
target bacteria from different types of crude samples (13, 27,
30). We are investigating the use of such methods for the
identification of
V. cholerae
O139 Bengal in clinical and
envi-ronmental specimens.
ACKNOWLEDGMENTS
This work was supported by a grant from the Swedish Medical
Research Council (grant B95-16X-11227-01A), the Go
¨ran Gustafsson
Foundation for Research in Natural Sciences and Medicine, and
IC-DDR,B. ICDDR,B is supported by countries and agencies which share
its concern for the health problems in developing countries.
REFERENCES
1.Albert, M. J.1993. Personal reflections on the discovery ofVibrio cholerae
O139 synonym Bengal: a tribute to team work and international collabora-tion. J. Diarrhoeal Dis. Res.11:207–210.
2.Albert, M. J.1994.Vibrio choleraeO139 Bengal. J. Clin. Microbiol.32:2345– 2349.
3.Albert, M. J., K. Alam, M. Ansaruzzaman, F. Qadri, and R. B. Sack.1994. Lack of cross-protection against diarrhea due toVibrio choleraeO139 (Ben-gal strain) after oral immunization of rabbits withV. choleraeO1 vaccine strain CVD103-HgR. J. Infect. Dis.169:230–231.
4.Albert, M. J., M. Ansaruzzaman, P. K. Bardhan, A. S. G. Faruque, S. M. Faruque, M. S. Islam, D. Mahalanabis, R. B. Sack, M. A. Salam, A. K. Siddique, M. D. Yunus, and K. Zaman.1993. Large epidemic of cholera-like disease in Bangladesh caused byVibrio choleraeO139 synonym Bengal. Lancet342:387–390.
5.Albert, M. J., A. K. Siddique, M. S. Islam, A. S. Faruque, M. Ansaruzzaman, S. M. Faruque, and R. B. Sack.1993. Large outbreak of clinical cholera due toVibrio choleraenon-O1 in Bangladesh. Lancet341:704.
6.Comstock, L. E., J. A. Johnson, J. M. Michalski, J. G. Morris, Jr., and J. B. Kaper.1996. Cloning and sequence of a region encoding a surface polysac-charide ofVibrio choleraeO139 and characterization of the insertion site in the chromosome ofVibrio choleraeO1. Mol. Microbiol.19:815–826. 7.Comstock, L. E., D. Maneval, Jr., P. Panigrahi, A. Joseph, M. M. Levine,
J. B. Kaper, J. G. Morris, Jr., and J. A. Johnson.1995. The capsule and O antigen inVibrio choleraeO139 Bengal are associated with a genetic region not present inVibrio choleraeO1. Infect. Immun.63:317–323.
8.Eklind, K., P. J. Garegg, L. Kenne, A. A. Lindberg, and B. Lindberg.1978. Structural studies on theEscherichia coliO111 lipopolysaccharide, p. 493.In
Proceedings of the 9th International Symposium on Carbohydrate Chemis-try.
9.Gorshkova, R. P., V. A. Zubkov, V. V. Isakov, and Y. S. Ovodov.1983. Structural features of O-specific polysaccharide from lipopolysaccharide of
Yersinia pseudotuberculosisVI serovar. Bioorg. Khim.9:1068–1073. 10. Hellerqvist, C. G., B. Lindberg, S. Svensson, T. Holme, and A. A. Lindberg.
1968. Structural studies on the O-specific side-chains of the cell-wall lipo-polysaccharide fromSalmonella typhimurium395 MS. Carbohydr. Res.8:43– 55.
11. Hultman, T., S. Bergh, T. Moks, and M. Uhle´n.1991. Bidirectional solid-phase sequencing ofin vitroamplified plasmid DNA. BioTechniques10:84– 93.
12. Hultman, T., S. Ståhl, E. Hornes, and M. Uhle´n.1989. Direct solid phase sequencing of genomic and plasmid DNA using magnetic beads as support. Nucleic Acids Res.17:4937–4946.
13. Islam, D., and A. A. Lindberg.1992. Detection ofShigella dysenteriaetype 1 andShigella flexneriin feces by immunomagnetic isolation and polymerase chain reaction. J. Clin. Microbiol.30:2801–2806.
14. Johnson, J. A., C. A. Salles, P. Panigrahi, M. J. Albert, A. C. Wright, R. J. Johnson, and J. G. Morris, Jr.1994.Vibrio choleraeO139 synonym Bengal is closely related toVibrio choleraeEl Tor but has important differences. Infect. Immun.62:2108–2110.
on May 15, 2020 by guest
http://jcm.asm.org/
15. Kenne, L., B. Lindberg, and E. So¨derholm.1983. Structural studies of the O-antigens fromSalmonella greensideandSalmonella adelaide. Carbohydr. Res.111:289–296.
16. Knirel, Y. A., L. Paredes, P. Jansson, A. Weintraub, G. Widmalm, and M. J. Albert.1995. Structure of the capsular polysaccharide ofVibrio cholerae
O139 synonym Bengal containing D-galactose 4,6-cyclophosphate. Eur. J. Biochem.232:391–396.
17. Komandrova, N. A., R. P. Gorshkova, V. A. Zubkov, and Y. S. Ovodov.1989. Structure of the O-specific polysaccharide chain of lipopolysaccharide of
Yersinia pseudotuberculosisserovar VII. Bioorg. Khim.15:104–110. 18. Lindberg, B., F. Lindh, J. Lo¨nngren, A. A. Lindberg, and S. B. Svenson.1981.
Structural studies of the O-specific side-chain of the lipopolysaccharide from
Escherichia coliO55. Carbohydr. Res.97:105–112.
19. Luk, J. M., U. Kongmuang, P. R. Reeves, and A. A. Lindberg.1993. Selective amplification of abequose and paratose synthase genes (rfb) by polymerase chain reaction for identification ofSalmonellamajor serogroups (A, B, C2, and D). J. Clin. Microbiol.31:2118–2123.
20. Maniatis, T., E. F. Fritsch, and J. Sambrook.1982. Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
21. Manning, P. A., U. H. Stroeher, and R. Morona.1994. Molecular basis for O-antigen biosynthesis inVibrio choleraeO1: Ogawa-Inaba switching, p. 77–94.InI. K. Wachsmuth, P. Blake, and O. Olsvik (ed.),Vibrio choleraeand cholera: molecular to global perspectives. ASM Press, Washington, D.C. 22. Morris, J. G., Jr.1995.Vibrio choleraeO139 Bengal—emergence of a new
epidemic strain of cholera. Infect. Agents Dis.4:41–46.
23. Nair, G. B., P. K. Bag, T. Shimada, T. Ramamurthy, T. Takeda, S. Yamamoto, H. Kurazono, and Y. Takeda.1995. Evaluation of DNA probes for specific detection ofVibrio choleraeO139 Bengal. J. Clin. Microbiol.
33:2186–2187.
24. Qadri, F., T. Azim, A. Chowdhury, J. Hossain, R. B. Sack, and M. J. Albert.
1994. Production, characterization, and application of monoclonal antibod-ies toVibrio choleraeO139 synonym Bengal. Clin. Diagn. Lab. Immunol.
1:51–54.
25. Qadri, F., J. A. Hasan, J. Hossain, A. Chowdhury, Y. A. Begum, T. Azim, L. Loomis, R. B. Sack, and M. J. Albert.1995. Evaluation of the monoclonal antibody-based kit Bengal SMART for rapid detection ofVibrio cholerae
O139 synonym Bengal in stool samples. J. Clin. Microbiol.33:732–734. 26. Ru¨ther, U. 1982. pUR allows rapid chemical sequencing of both DNA
strands of inserts. Nucleic Acids Res.10:5765–5772.
27. Seesod, N., J. Lundeberg, A. Hedrum, L. Åslund, A. Holder, S. Thaithong, and M. Uhle´n.1993. Immunomagnetic purification to facilitate DNA diag-nosis ofPlasmodium falciparum. J. Clin. Microbiol.31:2715–2719. 28. Shimada, T., G. B. Nair, B. C. Deb, M. J. Albert, R. B. Sack, and Y. Takeda.
1993. Outbreak ofVibrio choleraenon-O1 in India and Bangladesh. Lancet
341:1347.
29. Southern, E. M.1975. Detection of specific sequences among DNA frag-ments separated by gel electrophoresis. J. Mol. Biol.98:503–517. 30. Stark, M., E. Reizenstein, M. Uhle´n, and J. Lundeberg.1996.
Immunomag-netic separation and solid-phase detection ofBordetella pertussis. J. Clin. Microbiol.34:778–784.
31. Stroeher, U. H., K. E. Jedani, B. K. Dredge, R. Morona, M. H. Brown, L. E. Karageorgos, M. J. Albert, and P. A. Manning.1995. Genetic rearrange-ments in therfbregions ofVibrio choleraeO1 and O139. Proc. Natl. Acad. Sci. USA92:10374–10378.
32. Swerdlow, D. L., and A. A. Ries.1993.Vibrio choleraenon-O1—the eighth pandemic? Lancet342:382–383.
33. Verma, N., and P. Reeves.1989. Identification and sequence ofrfbSandrfbE, which determine antigenic specificity of group A and group D salmonellae. J. Bacteriol.171:5694–5701.
34. Weintraub, A., G. Widmalm, P. E. Jansson, M. Jansson, K. Hultenby, and M. J. Albert.1994.Vibrio choleraeO139 Bengal possesses a capsular poly-saccharide which may confer increased virulence. Microb. Pathog.16:235– 241.
35. Wyk, P. J., and P. R. Reeves.1989. Identification and sequence of the gene for abequose synthase, which confers antigenic specificity on group B sal-monellae: homology with galactose epimerase. J. Bacteriol.171:5687–5693.