Copyright 8 1997 by the Genetics Society of America
Calmodulin
Point
Mutations
Affect
Drosophila
Development and
Behavior
Heidi B. Nelson,' Robert
G.
Heiman,
ClareBolduc, Gae
E.
Kovalick,p
Penn
W
h
e
l
t
y
i
:
Michael Stern and
Kathy Beckingham
Department of Biochemistty and Cell Biology, Rice University, Houston, Texas 77005-1892 Manuscript received May 28, 199'7
Accepted for publication August 27, 1997
ABSTRACT
Calmodulin (CAM) is recognized as a major intermediary in intracellular calcium signaling, but as
yet little is known of its role in developmental and behavioral processes. We have generated and studied mutations to the endogenous Cam gene of Drosophila melanogastm that change single amino acids within the protein coding region. One of these mutations produces a striking pupal lethal phenotype involving failure of head eversion. Various mutant combinations produce specific patterns of ectopic wing vein formation or melanotic scabs on the cuticle. Anaphase chromosome bridging is also seen as a maternal effect during the early embryonic nuclear divisions. In addition, specific behavioral defects such as poor climbing and flightlessness are detected among these mutants. Comparisons with other Drosophila mutant phenotypes suggests potential CAM targets that may mediate these developmental and behavioral effects, and analysis of the CAM crystal structure suggests the structural consequences of the individual mutations.
I
N eukaryotes, fluxes of intracellular calcium play a major role in modulating cellular activity. Calmodu- lin (CAM) has emerged as a central mediator of these calcium-induced signaling events. Binding of calcium to the four sites on CAM regulates its ability to interact with numerous target proteins. From the CAM targets identified to date, a number of key molecular mecha- nisms by which CAM can influence intracellular events have been recognized. These include regulation of pro- tein phosphorylation, modulation of cyclic nucleotide levels, control of cytoskeletal dynamics and feedback regulation of intracellular calcium levels themselves, via effects on calcium pumps and channels (see COHEN and KLEE 1988 for review).A general model for CAM binding to its targets has evolved from molecular studies of individual target en- zymes (see CRIVICI and IKURA 1995 for review). How- ever, the larger task of
identifying
all the responses and processes regulated by CAM in an intact organism is still in its early phases. To date, in vivo studies with cultured cells and simple unicellular organisms have been the major sources of insight into these processes. Constructs that change the levels of wild-type CAM have been studied in cultured mammalian cells (RASMUSSEN and MEANS 198'7,1989) and Dictyostelium discoid.eum (LIU et al. 1992), and the phenotypes of various Cam muta- tions have been examined in Saccharomyces cm&iaeCmaponding author: Kathy Beckingham, Department of Biochemis-
try and Cell Biology, Rice University, MS 140,6100 S. Main, Houston, TX 770051892. E-mail: [email protected]
'Present address: Lark Technologies, Inc., Houston, TX 77024. *Present address: Department of Zoology, Miami University, Oxford, 'Present addms: Department of Zoology, Univenity of Texas, Austin, OH 45056.
TX 78712.
Genetics 147: 1783-1798 (December, 1997)
(DAVIS et al. 1986; GEISER et al. 1991; SUN et al. 1991; DAVIS 1992a; OHYA and BOT~TEIN 1994a,b), Schizosa- charomyces Po& (TAKEDA and Y A " o T o 1987; TAKEDA et al. 1989), Aspf?rgallw niduhns (RUMUSSEN et al. 1990) and Paramecium tetraurelia (KINK
et
al. 1990).These studies have demonstrated that Cam is an es- sential gene, with roles in the cell cycle and cell division. Interestingly, findings for
S.
cerevisiae indicate that the essential mitotic functions of CAM involve exclusively the calcium-free form of the protein, since a Cam mu- tant with no functional calcium binding sites shows good viability (GEISER et al. 1991). A further important finding from these studies is that mutations to specific amino acids of CAM can produce distinct phenotypes, indicating that different regions of the protein are in- volved in activating unique targets. Thus, in P. tetraure lia, viable Cam mutants show either diminished or exag- gerated avoidance responses, dependent upon the ex- act amino acid mutated and reflecting effects on different plasma membrane ion channels (KINKet
al. 1990). In S. cerevisiae, OHYA and BOTSTEIN (1994a,b)identified four different phenotypes among mutations to the phenylalanines of CAM and have further demon- strated intraallelic complementation between these mu- tations.
1784 H. B. Nelson et al.
first larval instar, demonstrating that the maternal s u p ply of CAM maintains embryonic development. In strik- ing parallel to the findings for P. tetraurelia, first instar Drosophila larvae null for Cam expression show sponta- neous avoidance reactions and other aberrant behav- ioral responses.
As
one approach to examining the roles of CAM in Drosophila beyond the point of death for null muta- tions, we have generated and analyzed point mutations to the protein coding region of the gene. Given the findings in S. cermisiae and P. tetraurelia, we anticipated that this approach might yield viable mutations whose phenotypes could give information on CAM function in discrete cellular processes. As we show below, the mutations we have generated identify several cellular, developmental and behavioral processes regulated by CAM, and comparison with other Drosophila mutant phenotypes suggests targets that could mediate this reg- ulation. In addition, the advanced structural studies of CAM permit us to speculate on how the individual mu- tations affect CAM structure and function.MATERIALS AND METHODS
Isolation of mutant Cum alleles: Males from a stock car- rying an isogenized second chromosome marked with bwwere fed 0.25% EMS in a 1% sucrose solution (LEWIS and BACHER
1968) and mated to either Sco Bl/CyO pT cn or Sco Bl/CyO R o i cn2 bw virgin females. Curly-winged progeny were individually mated to flies carrying one of two Cam alleles ( Camd3’, an RNA null caused by a deletion described in HEIMAN et al.
1996 or Cam’, which is the tAP-4 insertion of GOLDBERG et al.
1983) and Cy0 R o i cn’ bw. Ultimately, lines from seven flies
(5/7154 initially tested with Cam’ and 2/1579 with Cam”””4 gave progeny that showed reduced viability (530%) when in combination with either Cam chromosome. Stocks were made from these seven mutants and other initial candidates by cross- ing brown-eyed, curly-winged males from each positive vial to phenotypically identical virgin siblings. The H47 mutant was isolated as a female sterile in a similar EMS screen (1/1579
tested with Cam“”39, 0/1566 with Cam’) by assaying the fertility of F2 is0 bw*/CamR3”9 virgin females, where the asterisk repre- sents the mutated chromosome. Stocks for H47 and other candidates were generated by crossing the is0 bw*/CyO R o i cn2 bw brothers to Sco Bl/CyO R o i m2 bw virgin females.
AUele nomenclature: To aid in distinguishing among the various mutations, the following nomenclature has been used. Mutations that clearly aEect Cam have been given Cam allele superscript numbers, with the lowest new allele number (Cam”) indicating the weakest allele and the highest (Cam’) indicating the strongest allele. For mutations that affect resi- dues in calcium binding sites, a sufiix “c” has been added to the allele number. The affected site is indicated by a number following this “c.” Thus, Cam3c’ affects calcium binding site 1. The suffix “t” in allele Cam“indicates that a protein trunca- tion is produced.
Sequencing of mutant alleles: Total RNA was prepared from animals heteroallelic for a mutated chromosome and the Cam”””’ chromosome using Trizol (Gibco BRL) as per instructions. Five flies, 25 third instar, 35 second instar or 40
first instar larvae were used for RNA preparations and were frozen at -80” in 100-200 pl Trizol before crushing. Double- stranded cDNA was prepared using Superscript I1 reverse transcriptase (Gibco
BRL)
as per instructions, except thatrandom hexanucleotides were used to prime the first strand at 42”. PCR fragments were generated using primers (Life Technologies) to the 5‘ and 3’ untranslated regions of Cam:
5’ CACCCTCGAGCAGTAGCCGCG and 3’ AACCTCTA GATTACTGGGGTCCATAG. Amplicons were generated us- ing Deep Vent polymerase (NEB) and the following condi- tions: 5 min at 95”, and then 35 cycles of 30 sec at 94”, 45 sec at 52“, 30 sec at 72”. PCR products were cleaned with Wizard PCR Preps (Promega) as per instructions and digested with XhoI and XbaI (Boehringer Mannheim). After heat inactiva- tion of the enzymes and DNA precipitation, fragments were cloned into Bluescript (Stratagene) for DNA sequencing. Manual dideoxy sequencing was performed using primers that span the Cam open reading frame. Genomic mutations were distinguished from PCR errors and other artifacts by sequencing clones from at least two independent PCR ampli- fications, except for the C43 mutation (one PCR reaction only). One PCR fragment for C43 and one for Cam7 were generated by creating single-stranded (SS) cDNA as above, except using poly(dT) in place of hexanucleotides. These SS cDNAs were amplified with Taq polymerase, cloned directly into the pCR2.1 vector (Invitrogen) and sequenced with the PCR primers, a T7 primer (Stratagene) and an M13 reverse sequencing primer (USB)
.
The molecular lesion in the Cum* coding region: The Cam“ mutation appears to have been created by a small deletion (underlined, below), followed by a small duplication (bold, below), At base position 109 (encoding amino acid 36), the sequence ATG CGC TCC CTG GGG CAG AAT CCC ACA is changed to ATG CGC TCC CTG GCG CT CCC ACA, generat- ing a frameshift.
Mapping of mutant Cum alleles: The adult lethali associ- ated with three of the mutations (Cam””, Cam’, Cam
9
when transheterozygous to Camn-”’ was mapped after recombina- tion with a chromosome dominantly marked with J L Pin. Individual recombinant chromosomes marked with L were tested for the presence of the lethal mutation by a test cross to a balanced stock of Camn3”, followed by quantitation of all classes of progeny. Approximately 90 recombinant chrome somes were analyzed for each mutation, except for Cam6“‘, where 28 were examined. For the Cam4cl mutation, the effect on viability was too small for reliable mapping (see Table 1)and recombinant chromosomes (>200 total) were therefore tested based on the poor performance of Cam4cl/Camn”39 individuals in the climb test (see Table 5). The map position of Cam was established by mapping Cam””3’ with the J L Pin chromosome; Cam“””’maps -14 units to the left of Lobe, plac- ing the gene at -58. All four mutations ( Cam3”, Cam’, Cam“ and Cam4“) mapped approximately to this position. The pu- pal lethal phenotypes associated with C43/Camn3”’ and Cam7/ Cam””’ were mapped using recombination with the Cam’ w+
P element, which is at -34 relative to the transcription start site of the gene. These experiments placed the C43 and Cam7 lesions 15 and 20 kilobases (kb) , respectively, from the Cam’
w+ P element. The nucleotide altered by the Cam7 mutation is in fact 13 kb from this position (DOYLE et al. 1990). Mapping of the lethality of H47seen in transheterozygous combination with C43 (see text) placed this mutation closer than 48 kb to the Cam”’”’ deletion.
Generation of a Cam heat shock expression construct: The construct prepared (hsCMTAcam) has the Cam protein cod- ing sequence upstream of the metallothionein (MT) 3’ UTR and downstream of the hp70 promoter. MT-C, a CaSpeR derivative (PIRROTTA 1988) containing the Drosophila MT polyA site, was first created by inserting into CaSpeR the BamHI/PstI (base pairs 169-555) polyAfragment of pMTneo.
[pMTneo, a generous gift of R. DAVIS, Baylor College of Medi- cine, was constructed by inserting base pairs 122-785 of the
CAM Affects Drosophila Development 1785
Drosophila MT gene (LASTOWSKI-PERRY et al. 1985) into CosPneo (STELLER and PIRRO~TA 1985) .] Next, the hsp70 pro- moter was added to MT-C (to create MT-Chs) by inserting the appropriate BamHI/EcoRI fragment of pUChsneo ( S ~ L
LER and PIRROTTA 1985). Finally, the Cum protein coding sequence was added by inserting the BgfiI/BumHI Cam cDNA fragment of pJFM19 (MAUNE et aL 1992) into BamHIdigested MT-Chs, to generate construct hsCMTAcam. DNA was in- jected with helper plasmid pr25.7wc (KARESS and RUBIN
1984) as described (SPRADLING 1986). A tra'nsformant line
(y"""23p[w+ hsCMTAcam/66) that contains a viable insertion of this vector on the X chromosome was used in the heat shock experiments.
Heat shock rescue with ectopic
CAM:
yw6ic23p[w' hsCMTA cam]66; Cam"""'"/CyOqy+] flies were mated in combinations to produce mutant animals homo- or hemizygous for the hsCMTAcam vector. Mutant first instar larvae were selected( 1 12 for ~am~/Cam""' , 85 for C 4 3 / C ~ m " ~ ~ ~ and 15 for Cam8'/
and placed on yeasted grape-juice agar caps. The caps were stored at 25" and were placed at 37" for 1 hour each day until adulthood or death. Control animals remained at 25" throughout their lives.
viability amlysis: Five virgin females and five males of the appropriate genotypes (yw; Cam&/ or i ~ o b ~ / C y ~ p [ y + ] ) were stored slweek at 18" and then mated in yeasted vials at 25". Flies were transferred to fresh, yeasted vials every 3 days. Progeny from each vial were scored twice daily for 10 days total. The y- marker allowed for identification of both yellow- bodied mutant animals and siblings carrying the balancer and
y+. Each cross was set up in duplicate and reciprocally. Two control stocks were created for these studies: yw; Cam"339/
CyOPLy'] and yw; is0 bw/CyOP[y+J.
Lethal phase and deveIopmentaI delay analyses: To collect larvae, 10-15 mating pairs of various genotypes were placed in inverted, empty bottles with grapejuice agar caps and stored at 25". First instar larvae were handpicked from the plates and transferred to 3% agarose plates to facilitate scor- ing larval mouth-part color. Thirty mutant animals were is+ lated on two separate grape-juice plates and stored at 25". Animals were counted twice daily and staged based on mouth- part development and body size, until adults emerged or there were no longer any viable animals.
Adult morphology and fertility screens: In an initial screen, three y- males and three y- females of all allelic combinations that gave adult progeny were scored for thoracic bristle pat- tern and shape, wing size and shape, wing vein pattern, anten- nae and eye abnormalities and abdominal segmentation pat- tern. For a selected set of mutant combinations, much larger numbers of flies were analyzed to determine penetrance of mutant traits (see Table 3).
To screen quickly for fertility problems, adults from all mutant combinations were individually mated to three @eR flies and vials were checked for progeny after 10 days at 25". Further fertility tests on those genotypes that proved interest- ing involved testing 10 flies of each sex mated individually to
two OreR flies in a vial at 25". These vials were scored after 5 days for the presence of progeny. In one case, Cam3c'/Camn339
males were tested in a y+ background by mating stocks bal- anced with Cy0 Roi cn2 bw. For these experiments, an is0 bw/
Cy0 Roi cn2 bw control was generated.
All hemizygous combinations were also tested for male and female fertility by placing 15-20 mating pairs (crossed to flies) in inverted bottles with grape-juice agar caps. The caps were removed after 24 hours, the eggs were counted and, after at least 36 hours at 25", the number of unhatched eggs was determined. Certain female transheterozygous combina- tions were also tested in this fashion.
Dep-tion of embryonic terminal phenotypes: The un- hatched eggs from the experiments described above were analyzed for cuticle deposition, as described by WIESCHAUS and NOSSLEIN-VOLHARD (1986). In addition, fresh eggs were collected, representing 0-8 hour and 0-16 hour ages. These eggs were dechorionated, fixed and stored in 95% ethanol (WIESCHAUS and NOSSLEIN-VOLHARD 1986) prior to staining with 1.0 pg/ml DAPI (Polysciences) in Vectashield (Vector Labs).
Determination of OVBljan phenotypes Ovaries were dissected from 2 3 day old females, fixed in 3.7% formaldehyde for 20-
60 minutes and rinsed and stored in PES until stained with DAPI (as above). At least three ovaries of each genotype were analyzed for defects in nurse cell/oocyte/follicle cell nuclei numbers, positions, sizes and shapes and organization of DNA. In addition, follicle cell patterns surrounding the egg chambers were analyzed, as well as egg chamber development within ovari- oles and the average number of ovarioles per ovary.
Larval movement recordings: Larval locomotion was ana- lyzed for a small number of larvae (two to eight) of each mutant genotype in the hemizygous condition (videore- corded as described in HEIMAN et al. 1996). Larvae from recip rocal crosses were examined in all three stages, except for the early lethal Cam?
H e a t - i n d d padyak test: A slight modification of the p m cedure devised by WU d d. (1978) was used. Flies were in- duced into a standard glass culture vial and a cotton stopper was inserted to a depth of 6 cm from the bottom. Vials were immersed in a 37" water bath to a depth of 7 cm and incubated there for 5 min. The location and stance of the flies was in- spected immediately upon withdrawal of vials from the water. Wildtype flies typically cling to the cotton plug or to high posi- tions on the vial walls. Flies that show temperaturesensitive paral- ysis drop from these positions and are found lying immobile at the bottom of the vial. At least four individual flies (two of each sex) were tested for each genotype.
Bang-sensitivity (vortex) test: Sensitivity to mechanical shock was measured essentially as described by GANETZKY and WU (1982), except that vortex agitation of the flies was per- formed for 15 sec. Four individuals (two of each sex) were tested in the initial survey screening of all mutant combina- tions. Fifty young adults (<7 days old and 25 of each sex) were used for detailed studies of mutant combinations selected by this initial screening.
Climb test: Flies used for the detailed vortex testing (see above) were first tested for their ability to climb vertically as described by RICHARDS et al. (1996). Since some flies failed to climb at all, tests were terminated after 60 sec and these flies were arbitrarily assigned a score of 60 sec.
Flight test: Flies were tested for their ability to fly using a modification of the Sparrow box (DRUMMOND et al. 1991). In brief, a Coranaesthetized fly was placed in the top half of a sliced Eppendorf tube, covered and allowed to awaken. The open end of the tube was then placed through a hole in a piece of hard plastic covering a 1 liter graduated cylinder, and the fly was gently tapped out of the tube and scored for where it first landed. Flight ability was assayed by scoring landing at or above the 1 liter mark as a 10.0, at the bottom as a 0.0, etc. Each individual was tested four times (transferred by mouth pipet the final three times), and the scores were averaged. For each genotype, at least four individuals (two of each sex) were initially tested. Where abnormalities or dis- crepancies between individuals were found, an additional five or more flies of each sex were tested.
RESULTS
1786 H. B. Nelson et al.
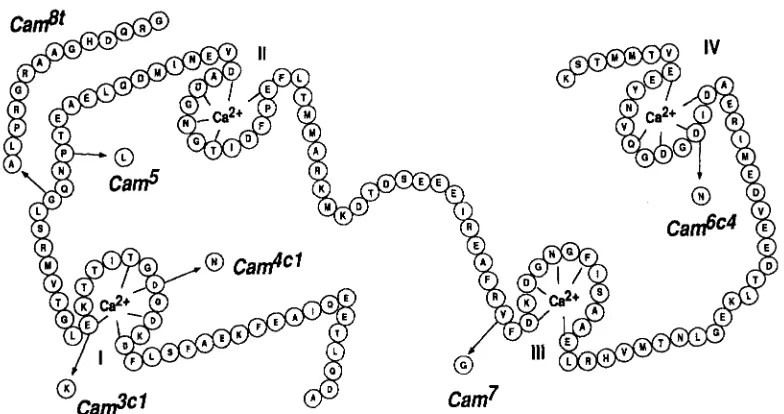
FIGURE 1.-Amino acid sequence changes in the EMSgenerated Cam alleles. The amino acid sequence of wild-type Drosophila CAM is shown (SMITH et al. 1987) together with the residue changes generated by EMS mutagenesis. Camk' changes aspartate
24 to an asparagine; Cam3'* changes glutamine 31 to a lysine; Cam8t generates a frameshift at glycine 40, creating a truncated protein of 53 amino acids; Cam' changes proline 43 to a leucine; Cam' changes valine 91 to a glycine; and changes
aspartate 131 to an asparagine. Diagram is derived from KLEE and VANAMAN (1982), with cy-helices shown as straight columnar arrays (nine residues N-terminal to binding sites and eight residues Gterminal), based on crystal data of Drosophila CAM
(TAYLOR et al. 1991). I-IV, the four calcium binding sites of CAM.
RIALS AND METHODS. In brief, we generated candidate mutations using random chemical mutagenesis of an isogenized chromosome marked with brown (is0 bw), followed by selection for reduced viability when hetero- zygous to either an RNA null ( Cum"339 or Cumnu"; HEI- MAN et ul. 1996) or a strong mutation (Cum') of the Cum gene. Ultimately seven mutations, showing <30% survival in combination with either of these two muta- tions, were selected for further study. Given that a large maternal supply of Cum mRNA is deposited in the ma- ture egg, we also screened for female sterility (described in MATERIALS AND METHODS). A single mutation (H47) was selected for analysis.
The Cum protein coding region was sequenced for all of the mutations. Six of the mutations change the CAM amino acid sequence (Figure 1 ) . Three mutations
( Cum3", Cum4" and Cum6c4) alter single amino acid resi- dues known to be involved in calcium binding (Figure 1). Surprisingly, one of these mutations ( Cum3") proved to correspond to a sitedirected mutation that was made previously in vitro in our laboratory ( MAUNE et ul. 1992). Cum' alters a proline residue present in the linker re- gion between sites 1 and 2 and Cum7 changes a residue in the N-terminal helix of binding site 3. The Cumsi mutation is not a single base alteration but a more extensive change (see MATERIALS AND METHODS) pre- dicted to produce a severely truncated version of CAM, consisting of the first calcium binding site (up to resi- due 39) and 14 new residues generated from a
frameshift. The CAM protein coding sequence is un- changed in mutations C43and H47, but RNA blot analy- sis established that Cum transcript levels are severely
reduced in larvae hemizygous for the C43 mutation and relatively unchanged in adults hemizygous for the H47 mutation (data not shown).
Mutant phenotypes map to the Cam locus: Although six of the mutations clearly alter the Cum protein coding sequence, it seemed formally possible that the mutant phenotypes could involve effects from additional muta- tions generated on the chromosomes by the EMS treat- ment. Recombination mapping, however, established that all of the mutant phenotypes seen in the hemizy- gous condition (that is, when transheterozygous to the null mutation), map to the region of the Cum locus
(see MATERIALS AND METHODS). In particular the H47 and C43 mutations, which do not alter the Cum amino acid sequence, were mapped to within 48 and 15 kb of the Cum transcription start site, respectively. To avoid completely the potential for problems from additional mutations present on mutant chromosomes, all analysis of mutations was therefore performed in the hemizy- gous condition (that is, heterozygous to the null muta- tion) or in transheterozygous combinations. Thus, when phenotypes of individual mutations are discussed below, unless otherwise stated, it is their effects in the hemizygous condition that are being described.
CAM Affects Drosophila Development
TABLE 1
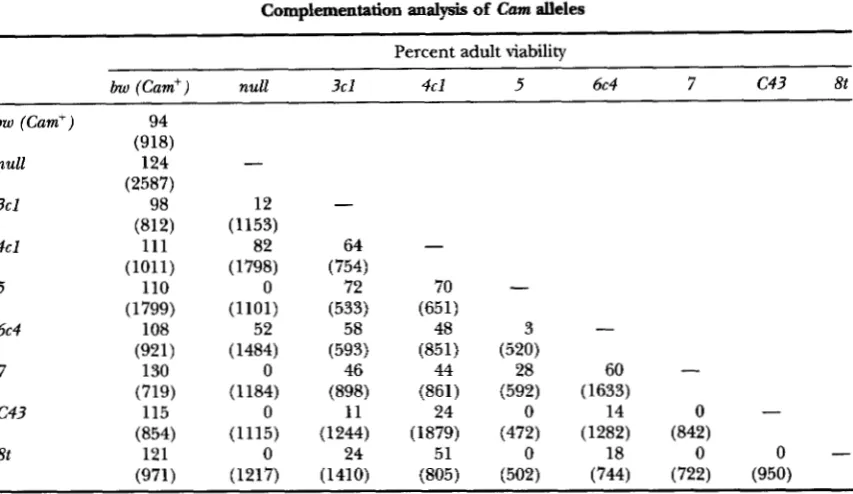
Complementation analysis of Cam alleles
1787
Percent adult viability
bw (Cam') null 3 c l 4cl 5 6c4 7 c43 8t
bw (Cam')
null
3 c l
4c1
5
6c4
7
c43
8t
94 (918)
124 (2587)
98 (812)
111 (1011)
110
(1 799) 108
130 (719)
115 (854)
121 (921)
-
12 (1153)
82 (1798)
0 (1101)
52 (1484)
0
(1 184) 0 (1115)
0
-
64 (754)
72 (533)
58 (593)
46 (898)
11
(1244) 24 (971) (1217) (1410) (805) (502)
60 (1633)
14 0
-
(1282) (842)
18 0 0
--
(744) (722) (950)
The percent viability of various mutant combinations was determined by counting both mutant adults (y-)
and adults carrying the balancer chromosome (y+) from the appropriate crosses (MATERMS AND METHODS).
bw (Cum') is the control (Cum') is0 bw chromosome in this and all following tables. The Cam mutations are
ordered with the weakest, Cum3cJ, listed first, and the strongest, Cum*', listed last, as determined from lethal phase analysis (see Table 2). All data have been corrected for the percent mutant class expected to emerge as follows: (#y-/[#y' X 0.51) X 100. In addition, each heterozygous combination has been divided by 0.94
(to correct for the observed 94% viability of bw (Cum'); see Table) and each hemizygous combination has been divided by 1.24 (to correct for the observed 124% viability of bw (Cum')/Cum""'; see Table). All crosses were performed reciprocally, and data for at least four repetitions of each cross were collected (duplicates of the cross in both parental directions). The data for all repetitions was combined and averaged since, for all but two allelic combinations, the averages of the reciprocal crosses were within 10% of each other. The exceptions were (1) the cross of Cum4" X Cum""" for which maternally derived Cum4"produced 75% viability and paternally derived Cum4" produced 88% viability and (2) the cross of C43 X Cam for which maternally derived C43 produced 10% viability and paternally derived C43 produced 38% viability. Values in parentheses are the total number of adults scored. The dash indicates that the homozygotes were not tested.
to the three fully penetrant lethal mutations isolated here: Cam7, CamBt and C43 (see below). Further, more detailed studies of the electrophysiological and physio- logical defects associated with the Cam3"' mutant (see below) have shown that these can be rescued com- pletely by ectopic expression of CAM from this heat shock construct
(L.
ARREDONDO, H. B. NELSON, K. BECKINGHAM and M. STERN, unpublished data).Effects on overall viability and a determination of the lethal phases; an allelic series of Cum mutati- ons: Experiments to quantitate effects on viability and to identify the point of death for mutant animals were part of our initial characterization. Viability was deter- mined under standardized conditions (see MATERIALS AND METHODS) for all mutations (1) in the hemizygous condition, (2) in all possible transheterozygous combi- nations and (3) in trans to the Cam+ is0 bw chromosome (Table 1). The point in the life cycle at which mutant individuals died was determined for all alleles in the hemizygous condition and for combinations of the more lethal alleles (Table
2).
Given that, until very latein embryogenesis, maternally derived CAM is the only CAM present in most embryonic tissues
(B.
ANDRUSSand K. BECKINGHAM, unpublished observations), we were interested in whether the effects of a particular mutant CAM might depend upon its being present in the newly laid
egg.
All crosses for these experiments and many others presented here were therefore performed reciprocally, so that this issue could be addressed. This aspect is discussed in the final section of the RESULTS.The relative strengths of the seven recessive alleles
was
evaluated from the lethal phase data in Table 2 and was used to order them loosely into an allelic series. This series, from strongest to weakest mutation, is as follows: CamBt>
~ 4 3>
cam7>
>
cam5>
cam4"1788 H. B. Nelson et ul.
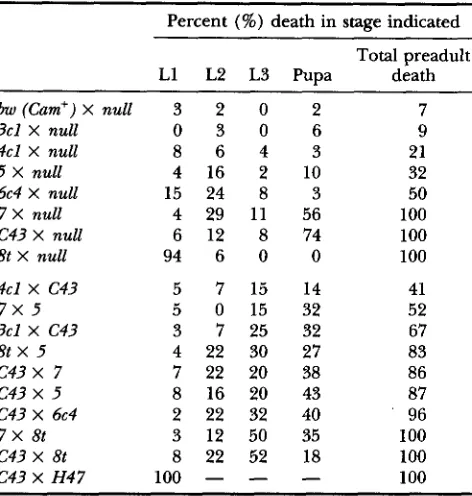
TABLE 2
balysis of the lethal phases of Cam mutations
Percent (%) death in stage indicated
Total preadult
L1 L2 L3 Pupa death
bw (Cam') X null 3 2 0 2 7
3cl X null 0 3 0 6 9
4cl X null 8 6 4 3 21 5 X null 4 16 2 10 32
6c4 X null 15 24 8 3 50
7 X null 4 29 11 56 100
C43 X null 6 12 8 74 100
8t X null 9 4 6 0 0 100
4cl X C43 5 7 15 14 41
7 x 5 5 0 15 32 52
3cl X C43 3 7 25 32 67
8t
x
5 4 22 30 27 83c 4 3
x
7 7 22 20 38 86c 4 3
x
5 8 16 20 43 87C43 X 6c4 2 22 32 40 96
7X 8t 3 12 50 35 100
C43 X 8t 8 22 52 18 100
C43 X H 4 7 100
-
- -
100 The lethal phases for each Cam allele in the hemizygous condition and for transheterozygous combinations of thestronger alleles were determined. The y- mutant larvae from the appropriate crosses were separated from their y+ siblings (see MATERIALS AND METHODS) and grown in noncompetitive conditions. The percent of y- animals that died in each stage was determined and is shown, together with the total percent preadult death (TPD). As for Table 1, data for reciprocal versions of the crosses were collected but then combined since for all but two crosses the TPD averages were within 10% of
each other. The exceptional crosses were (1) Cum4"' X C43
where maternally derived Cum4" produced 23% TPD and a ternally derived Cum4"' produced 57% TPD and (2) Cam3", C43 where maternally derived Cum3c' produced 80% TPD and paternally derived Cum3" produced 53% TPD.
individuals are raised alone (4 Tables 1 and 2), In fact, Cam' appears to be a fully penetrant lethal mutation when assayed in our adult viability studies.
As
will be shown below, Cam3c' and Cam5 mutant animals show pronounced adult behavioral defects and it seems likely that behavioral abnormalities contribute to their failure to survive in the presence of more vigorous animals. Certain Drosophila mechanosensory mutants also ex- hibit decreased survival in competition with nonmutant siblings(KERNAN
et al. 1994). It is also noticeable that Cam4" and Cam- mutant animals show greater adult viability under the standardized conditions used to gen- erate the data in Table 1 than the <30% survival rate used as the original selection criterion (see above). It seems likely that these differences in viability also reflect behavioral defects in newly emerged adults that were aggravated by crowded conditions in the original screens.Only the strongest allele, the completely penetrant lethal mutation Cam", behaves like Camnuu and pro-
duces death in the first larval instar (Table 2). In fact, this mutation has a more deleterious effect than Cam""" on developmental progression when analyzed in combi- nation with several other alleles (Table 2). The other
two strong lethal alleles, Cam7 and C43, proved to be pupal lethal mutations: all animals die before adult- hood, with most deaths occurring during the pupal stage (Table 2). Both mutations also confer profound developmental delay, increasing the time spent in the larval stages by twofold (data not shown).
A strong genetic interaction between C43 and
H47:
The viability studies provided insight into the na- ture of the two mutations (C43 and H47) that do not affect the protein coding sequenceCAM.
C43 clearly interacts strongly with the Cam locus, producing com- pletely penetrant lethality or markedly reduced viability with all proven Cum alleles (Tables 1 and2).
In contrast, the H47 mutation showed no strong effect on viability in combination with any of the established Cam alleles (data not shown). Further, the female sterility associ- ated with the H47 mutation (see earlier) proved to be a dominant trait unrelated to the mutational status of the Cam locus. Nevertheless, H47 proved to interact very strongly with the C43 mutation.As
shown in Table2,
although C43/Cumn""animals survive to pupation and H47/CamnUu animals survive to adulthood, all C43/H47 animals die during the first larval instar. This interac- tion is independent of the parental origin of the H47 mutation. The implications of this strong genetic inter- action are examined in the DISCUSSION.Developmental phenotypes associated with Cum mu-
tations: The strong, pupal-lethal mutation Cam7 con- fers unusual and distinctive developmental abnormali- ties. Most animals carrying Cam7 in the hemizygous con- dition survive to pupate but never emerge from their pupal cases (Table
2).
Their pupal cases are smaller than normal, with a distinctive "Michelin-man" pheno- type of deeply indented rings corresponding to the junctions of the larval body segments (Figure 2C). Strik-ingly, most Cam7 individuals (54% of 88 examined) that develop within the pupal cases are headless or have partial head structures. This headlessness is the result of a failure in head eversion, as opposed to a failure of head development. The imaginal discs that give rise to the head are normal in Cam7 late third instar larvae. Indeed, complete but inverted head structures, includ- ing fully formed eyes, could be dissected out of the thoraces of Cam7 pupae (Figure 2F). A small fraction of Cam7 pupae have fully everted heads, suggesting that Cam7 produces other defects that result in pupal death. In combination with Camst, which encodes a truncated CAM, Cam' produces a similar effect on pupal case mor- phology, but fewer pupae develop far enough to show failed head eversion. The phenotype of failed head eversion was first identified for the mutation l(2)
q p t o -
CAM Affects Drosophila Development 1789
1
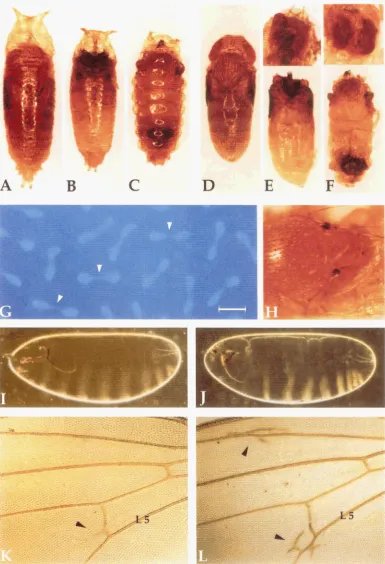
FIGURE 2.-Morphological defects of Drosophila Cum mutants. (A-C) Pupae of wild-type OreR (A), crc (B) and C~rn~/Cum'"'~~
(C) animals. Note that both crc and Cum7/CumnuN pupae are smaller than wild-type. The C ~ m ~ / C u r n " " ~ pupal case has deeply indented concentric rings, giving it a "Michelin-man" appearance. (D-F) Pharate adults dissected from the pupal cases shown in A-C: OreR (D) c-rc (E) and Cum7/Cum""" (F). The crc and C ~ m ~ / C u m " " ~ animals have failed to evert their heads. Insets above E and F eyes from the dissected heads that developed inside the thoraces of the pu ae shown below. (G) Chromosomes showing anaphase bridges (arrowheads) in a syncytial blastoderm embryo laid by a yw; Cum /Cum3" mother. Bar, 13.5 pm. (H) Adult yw;
C ~ m ~ ~ ' / C u m ~ fly displaying bilateral cuticular scabs on the notum. (I and J) Embryo cuticle preparations. (I) A wild-type embryo laid by a control is0 bw mother.
u)
A lateralized embryo (cuticle shows defects in the dorsal-ventral axis) laid by an H47/Camndmother. (K) Wing from a yw; iso bw fly showing the worst vein defect seen in the control flies: a faint posterior spur (arrowhead) growing from the posterior crossvein. (L) Wing from a yw; C ~ r n ~ ~ ~ / C u m ~ ' fly showing ectopic veins located on the longitudinal veins L2 and L5 (arrowheads). L5, longitudinal vein L5.
1790 H. B. Nelson et al.
Cam. Although crc individuals make normal pupal cases (Figure
2B),
the range of defects in head eversion is very similar to theCam7
phenotype (Figure2E).
The phenotype of C43, the other pupal-lethal muta- tion, is distinct from that of Cam7. Most C43pupae (55%
of 82 examined) do not develop past the white prepupal stage. Because of the unusual biology of Drosophila, death at the pupal stage can reflect a defect in a ubiqui- tous component of mitosis (GIRDHAM and GLOVER 1991). C43 does not appear to affect mitotically active tissues, however. The imaginal discs in C43/CamnUu white prepupae are normal in size, and no obviously defective mitotic figures have been identified in larval neuroblasts. In addition, many C43/CamnUupupae (45%
of 82 examined) die as phenotypically normal pharate adults. Eversion of the anterior spiracles, an early event in pupation, fails to occur in most C43/CamnUN pupae
(66% of 80 examined).
A
broad survey of external adult morphology was per- formed for all mutants and mutant combinations that gave survival to the adult stage (see M A ~ W AND METHODS). From this analysis, three types of adult mor- phological defects were identified that were produced by many allelic combinations. None of these pheno- types proved completely penetrant, however, and de- tailed quantitation of penetrance was only performed for a subset of the mutant combinations (see Table 3). The most penetrant morphological defect identified was ectopic wing veins. At a very low frequency (3%), the control isobw
flies exhibit very short extra vein spurs growing away from the posterior crossvein in a posterior direction (Figure2K).
In contrast, many Cam mutant combinations show, at high penetrance (up to68%
in the mutant combinations examined) much longer and more pigmented posterior crossvein spurs (Table 3). Certain mutant combinations also show an additional anterior-growing spur, and, less frequently, ectopic veins appear along the second or fifth longitudinal veins (Figure2L).
The second specific, but incompletely penetrant de- fect seen in a number of Cam mutant combinations was the appearance of either lateral or bilateral cuticular "scabs" on the notum (Figure
2H).
These black, mela- notic scabs were most often found near the anterior scutellar bristles and frequently had a hardened, "tar- like" appearance. This phenotype was never observed in the control flies and was seen most often in Cam combinations carrying Cam3" (Table 3). A third pheno- type that was observed in several transheterozygous combinations involving the C43mutation was an altered wing shape, with the entire blade curved downward like a spoon (data not shown).Maternal effects
of
Cammutations:
Female fertility was examined (see MATERIALS AND METHODS) in all mu- tant combinations that gave viable adult progeny and only H47 emerged as a female sterile mutation of high penetrance.As
described above, H47was the only muta-TABLE 3
Adult morphological phenotypes of Cam mutant combinations
No. of Percent animals
Phenotype Genotype mutant scored
Wing vein
defects 4 c 1 / 5 68 204
4cl/ 8t 59 117
6c4/ 7" 57 494
C43/ 6c4' 57 268
3c1/4cI 46 153 C43/ 4cl 40 187
3 c 1 / 5 38 184
4 c l / 7b 33 403
3c1/8t 30 103
7/ 3cl 25 445
6c4/ null 39 253
4cl/nullb 24 543
6c4/ bw (Cam' ) 31 203
8 t / h (Cam') 27 202
4 c l / b w (Cam') 27 224
C43/bw (Cam') 25 114
bw (Cam+)/bw (Cam+) 4 234
nuU/bw (Cam+? 6 278
Scabs 3 c 1 / 5 23 66
3 c l / 4 c l b 8 229
5 / 4 c l 7 113
3c1/8t 4 103
bw (Cam+)/bw (Cam+) 0 234
Mutant animals were scored for morphological defects (see text). Shown here are those genotypes that show a significant percent of defects compared to the yw; iso bw control. The vein list contains those combinations scored that produce >20% animals with vein defects and for which 100+ animals were scored; the scab list contains those where scabs were noted and 65+ animals were scored. Genotype is fint muta- tion donated by the mother and the second by the father, except for those combinations noted as a or b.
63% of progeny for this allelic combination showed vein defects when Cam6c4 was donated maternally and 41 % when Cam6c4 was donated paternally.
*Weighted average scores from reciprocal crosses where only minor differences in the two data sets were observed.
tion initially isolated as a female sterile mutation. Fe- males carrying H47 lay eggs, but most of these fail to hatch. We have studied these embryos extensively and determined that they show defects in dorsal-ventral pat- terning (Figure
25)
associated with reduced levels of nuclear DORSAL protein(H.
B.
NELSON and K BECK- INGHAM, unpublished observations). However, given that this maternal effect of 1147 is dominant and unin- fluenced by Cam mutations and that H47 produces no alteration to the Cam protein coding sequence, this fe-male sterility may not involve a role for
C A M .
CAM AfTects Drosophila Development
TABLE
4Maternal effects of Cam mutants
1791
Percent No. Percent no cuticle
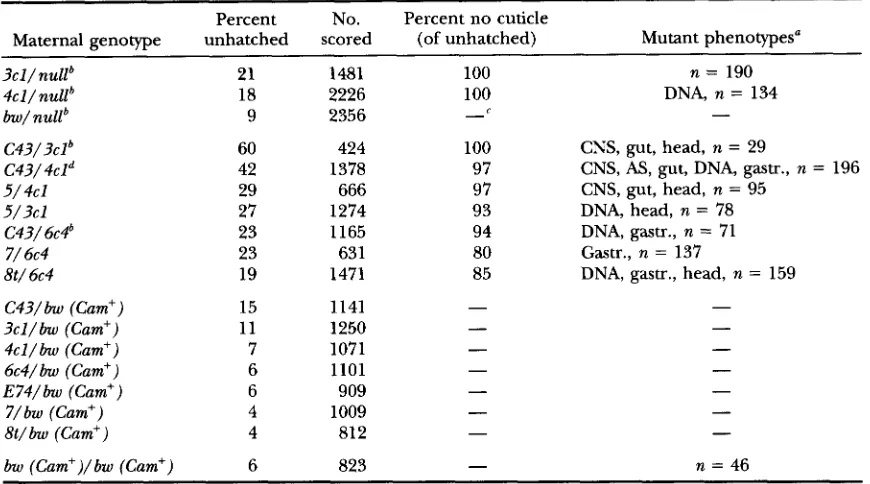
Maternal genotype unhatched scored (of unhatched) Mutant phenotypes"
3cl/nuUb 21 1481 100 n = 190
4cl/nuUb 18 2226 100 DNA, n = 134
bw/ nullb 9 2356
-
-
C43/3clb C43/4cld
5/ 4cl 5 / 3 c l
C43/ 6 ~ 4 ~
7/ 6c4
8t/ 6c4
60 424 100 CNS, gut, head, n = 29
42 1378 97 CNS, A S , gut, DNA, gastr., n = 196
29 666 97 CNS, gut, head, n = 95
27 1274 93 DNA, head, n = 78
23 1165 94 DNA, gastr., n = 71
23 631 80 Gastr., n = 137
19 1471 85 DNA, gastr., head, n = 159
C43/bw (Cam+) 15 1141
3 c l / b w (Cam') 11 1250
4 c l / b w (Cam') 7 1071
6c4/bw (Cam') 6 1101
E 74/ bzu (Cam") 6 909
-
7 / b w (Cam") 4 1009
-
8 t / h (Cam') 4 812 -
-
-
-
-bw (Cam")/bw (Cam') 6 823
-
n = 46Hatch rates for mothers of the genotypes listed were determined after mating to wildtype males; -100 unhatched eggs were analyzed for cuticle deposition and another -100 were stained for DNA (MATERIALS AND METHODS). Embryonic defects are listed as follows: DNA, chromosomal bridges seen at syncytial blasto- derm; CNS, gut, head, defects in CNS condensation, gut development and head shape, respectively; gastr., gastrulation defects such as ectopic folds; AS, defects in amnioserosa localization.
n is the number of embryos scored by staining for DNA.
Data for mothers obtained from the two reciprocal versions of the cross were comparable and are shown
Not tested.
For this allelic combination, 62% of embryos failed to hatch when Cam4" was maternally derived and 27% when Cam4" was paternally derived. For other genotypes, data shown are for mothers from a single direction of the cross and the maternally derived allele is listed first. No. scored, number of eggs counted for hatch rates. Percent no cuticle, number of unhatched eggs that did not contain cuticularized embryos. Mutant phenotypes, defects seen in
>lo%
of developed embryos, except for chromosomal bridges (see above). combined.tions that give adult survivors in the hemizygous condi- tion ( Cam4" and Cam3") have a detectable maternal effect on egg hatch rate (Table 4), and several mutant combinations show equivalent or more pronounced maternal effects on embryonic development (Table 4). Embryonic death occurs mainly before cuticle deposi- tion for all allelic combinations, and embryos display a range of phenotypes (Table 4). The most extreme ef- fects were seen for Cam4"'/C43 and C43/Cam3"' mothers. One particularly intriguing defect was found in embryos from mothers carrying several Cam mutant combina- tions. In the syncytial blastoderm stage, these embryos showed extensive chromosomal bridging at late ana- phase (Figure 2G). Oogenesis was also examined for all mutant combinations showing maternal effects, but there were no observable defects in the ovaries from most of these mothers. The exception was Cam8f/Cam" females, which showed nearly a 2.5-fold increase in de- generating egg chambers as compared to the is0
bw
controls (data not shown).
Male fertility studies proved inconclusive due to the
presence of a yellow mutation
(y-),
sincey-
males ex- hibit a mating disadvantage when paired with wildtype females (BASTOCK 1956). However, Cam3" males tested 100% sterile (see MATERIALS AND METHODS) and were therefore examined in a wild-typey+
background. Wild- type females mated to these males laid very few eggs and, although some hatching was observed, the hatch rate was significantly decreased. Given that Cam3" con- fers other strong behavioral defects (see below), it is possible that the Cam''' mutation may interfere with male mating behavior.1792 H. B. Nelson et al.
TABLE
5Coordination of Cum mutants
6
bw (Cam') null 4cl 3cI 5 c 4 3A. The climb test 0
bw (Cam') 2 5* 3* 36 F/27 M* 18" 4*
4cl 9 F/3 M* 40 F/33
M*
-
48 F/55 M*-
-
6c4
3cl 13" 52 F/47
M*
-
-
-
57"c 4 3 12 F/5 M*
-
-
46*-
-
?lull 9*
-
12 F/19 M* >60* 52 F/>60 M* a-
8*
-
7 F/14M*
-
-
-
5 8 F/15 M* 49 F/>60M* a
-
--
-
B. The vortex test
0
bw (Cam') 6 3 3 4 4 3
null 4
-
2 35 F/58 M* 22 F/48 M* a4cl 2 4
-
17 F/38 M*6c4 3
3 c l 4 54 F/33 M*
-
-
- 38 F/34 M*5 17 F/32
M*
41 F/86M*
-
c 4 3 2
-
-
24 F/31 M*-
-
-
-
-
-
3-
-
-
-
-
-
~~ ~ ~ ~~ ~~~
Data are presented for a subset of genotypes identified by initial screening as defective in coordination (see.text). The male ( 6 ) and female (
0
) symbols indicate the allele derived from the father and mother. Numbers represent average time in secondsto complete the test (see MATERIALS AND METHODS). Twenty-five individuals of each sex were tested. If the average scores for males and females showed a relatively large, statistically significant difference
(P
< 0.05), they are presented separately. F andM, female and male scores. In A, >60 is used to denote that most animals failed to complete the test in 60 sec. Dashes indicate genotypes that were preadult lethal or not tested.
*
Significantly worse than the control, based on a t-test. Climbing (A), then recovery from vortexing (B), were tested sequentially on the same flies.a Cam5 hemizygous individuals obtained by rearing apart from phenotypically wild-type sibs (see text). As can be seen, genetic
combinations for which the directionality of the cross produced large differences in performance are (1) Cam3c' X Uo bw and
(2) Cum*' X Cam""" for the climb test and Cam' X is0 bw for the vortex test.
showed non-wild-type characteristics, such as slight twisting. These abnormalities were not systematically studied, however.
Three tests were initially used to survey adult Cam mutant flies for behavioral abnormalities. These were tests for (1) heat-induced paralysis,
(2)
sensitivity to mechanical shock (bang sensitivity) and (3) flight abil- ity. In an initial survey of all mutant combinations that gave adult survivors, none showed heat-induced or bang-induced paralysis, although several Cam mutant combinations showed some delay in righting themselves after the bang-sensitivity (vortex) test. For these allelic combinations, general mobility and coordination were therefore further examined by testing 50 individuals in both the vortex test and a climb test (see MATERIALSAND METHODS). Both tests were performed on each
fly
so that performance in the two paradigms could be compared.The climb test proved the more sensitive assay for locomotor abnormalities,
with
all the allelic combina- tions tested displaying statistically significant defects compared to is0bw
(compare Table 5 , A and B). In fact, all of the mutations tested produced statistically significant dominant effects in this assay when heterozy- gous to is0 bw (Table 5). These findings suggest thatmore transheterozygous allelic combinations would prove defective if tested in this paradigm. Although there is an overall correlation between performance in the climb test and the vortex test, there are notable differences. In particular, Cam4"' hemizygous flies are essentially wild-type with respect to bang sensitivity but show marked abnormality in the climb test.
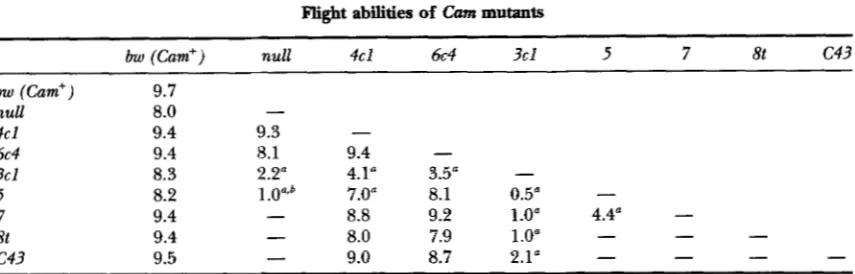
In both the climb and vortex tests, male performance was significantly worse than female performance for most mutant combinations. However, for flight ability, no marked differences in male and female performance were detected (Table 6). The most striking finding from flight testing was that Cam'"' is essentially flightless in all allelic combinations.
As detailed above, when Cam5 hemizygous individuals are reared separately from phenotypically wild-type sib- lings, some of them survive to the adult stage. These animals are noticeably ill coordinated and weak and do not live long as adults. A small number were tested in the vortex, climb and flight tests and were found to perform very poorly in all three assays (Tables 5 and 6).
CAM Affects Drosophila Development
TABLE 6
Flight abilities of Cum mutants
1793
bw (Cam") null 4cl 6c4 3cl 5 7 8t c 4 3
bw (Cam") 9.7
null 8.0
4c1 9.4 9.3
6c4 9.4 8.1 9.4
3cl 8.3 2.2" 4.1" 3.5"
-
5 8.2 1.0"b 7.0" 8.1 0.5"
-
7 9.4
-
8.8 9.2 1.04 4.4"-
st 9.4
-
8.0 7.9 1 .O"-
c 4 3 9.5
-
9.0 8.7 2.1"-
-
-
-
-
-
-
-
-
~ ~~~~~~ ~ ~
The ability to fly was tested for all mutant combinations as described in MATERIMS AND METHODS. Average scores are shown; 0 = no flight ability; 10 = wild-type flight ability. Data for males and females were comparable and were pooled. Each cross was set up reciprocally and data for both versions of the cross were collected. Data averaged for both versions of each cross are shown since only one allele combination ( Cam5 X Cam4c')
produced a large, statistically significant difference in performance for individuals from the two versions of the cross. For this combination, maternally derived Cam' produced an average score of 5.2 and paternally derived Cam5 produced an average score of 8.5.
a Scores indicating serious flight impairment (statistically significant from control and <5.0). Dashes as
previously.
Cam5 hemizygous individuals obtained by rearing apart from sibs (see text).
ual Cam mutations might depend on whether a particu- lar mutant CAM was present in the newly laid egg. Most of the allelic combinations studied were therefore gen- erated from reciprocal versions of the crosses so that phenotypes could be examined with each allele pre- sented from either the male or female parent. Combi- nations for which the directionality of the cross made a noticeable difference are discussed in the legends of Tables 1-6. In general, these are few in number, and, although for one allelic pair (C43 and Cum"") the direc- tionality of the cross appears to affect at least three different parameters, it does not affect them in a consis- tent manner (see legends to Tables 1, 2 and 4). No consistent effects of maternally derived mutant CAM were therefore distinguished with this series of Cam mutations.
DISCUSSION
Mutability of the D. melunogmter Cam gene: In a mu- tagenesis screen of 8733 chromosomes, we identified six unique mutations in the Drosophila Cam gene. This point mutation frequency of one per -1500 chromo- somes is in marked contrast to findings for the single S. cerevisiae Cam gene. A screen of 35,000 colonies car- rying a randomly mutagenized yeast Cam gene yielded only null mutations (DAVIS 1992b). No random point mutations that prevented growth were identified in this or other studies (OWA and BOTSTEIN 1994b), leading to the suggestion that Cum is not readily inactivated in yeast.
We can identify several factors that could contribute to the very different outcome of our screen compared to those in yeast. Selection for reduced viability coupled with the deleterious effects of competition from pheno-
typically wild-type siblings may have allowed us to isolate mild mutations that might go undetected in yeast. But, more importantly, the complex organization and devel- opment of Drosophila presumably provides many more target interactions in which a given CAM mutant pro- tein must prove competent. An amino acid change that can support growth in yeast may not fulfill the more complex demands of multicellular existence.
Morphological and maternal effect phenotypes of Cam mutants; indications of potential targets: Given previous studies, it seemed possible that point muta- tions in Drosophila Cam might affect a limited range of CAM-target interactions and thus delineate develop mental and cell biological processes in which CAM par- ticipates. Our findings from this screen have validated this approach. We have identified at least three develop mental/cellular processes requiring CAM and gained insight into the possible targets that may mediate the involvement of CAM in these processes.
1794 H. B. Nelson rf al.
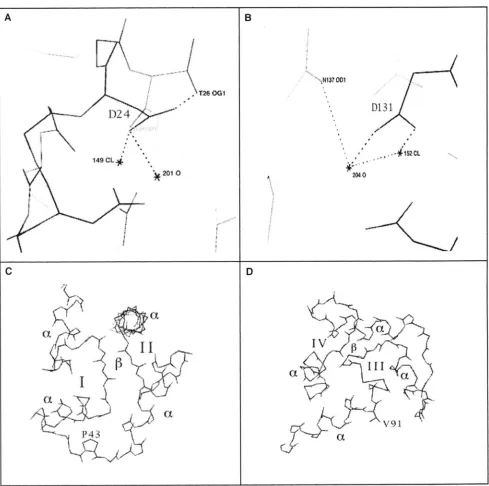
C
B
D
FIGURE 3.-Predicted roles of point mutations to Drosophila CAM from examination of the crystal structure. The roles of the residues affected by the four new point mutations as visualized using the program CHAIN to examine the crystal structure of Drosophila CAM (TAYI.OR rf 01. 1991). (A) Aspartate 24 (D24). One of the carboxylate oxygens of the sidechain is one of the seven oxygen atoms that coordinate the calcium ion (149 CL) in calcium binding site I and is also involved in a hydrogen bond to the water molecule present in the calcium binding site (201 0 ) . The second carboxylate oxygen, which is changed to an amino group in the D24N mutation (Cam'"), forms a hydrogen bond to the side-chain of threonine 26. This hydrogen bond may play a role in positioning the sidechain of D24 correctly for its role in calcium coordination. Loss of this hydrogen bond in the D24N mutation may therefore decrease calcium binding affinity at site I. ( R ) Aspartate 131 (D131). One of the side- chain carboxylate oxygens coordinates the calcium ion ( I 5 2 CL), while the second oxygen forms a hydrogen bond to the H 2 0 molecule in the calcium binding site (204 0 ) . This water molecule provides one of the oxygens that coordinates the calcium ion and thus loss of the hydrogen bond to this molecule as a result of the D131N mutation in Cam'"' could destabilize the water molecule and weaken calcium binding at site IV. (C) Proline 43 (P43). This residue is in the rigid linker between calcium binding sites I and 11. By providing rigidity in this linker, the imino bond of the proline is predicted to contribute to the close positioning of loop regions of calcium binding sites I and I1 and the formation of the short stretch of antiparallel