N O C IC E PTIN AN D THE OP4 R E C E PTO R IN THE
A UDITOR Y S Y S T E M
Wendy J N oble
A thesis presented to the University o f London f o r the degree o f
D octor o f Philosophy.
k
UCL
Institute o f Laryngology and Otology University College London
London, UK.
ProQuest Number: 10015029
All rights reserved
INFORMATION TO ALL USERS
The quality of this reproduction is dependent upon the quality of the copy submitted.
In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uest.
ProQuest 10015029
Published by ProQuest LLC(2016). Copyright of the Dissertation is held by the Author.
All rights reserved.
This work is protected against unauthorized copying under Title 17, United States Code. Microform Edition © ProQuest LLC.
ProQuest LLC
789 East Eisenhower Parkway P.O. Box 1346
Acknowledgements
/ would like to express my sincere thanks to my supervisors Dr. Hilary Dodson and Professor Andrew Forge fo r giving me the opportunity to pursue this PhD in their laboratory and fo r all their time and effort during the course o f my studentship. In particular, I would like to thank H ilary fo r her tireless effort - fo r passing on her expertise in this line o f work from the very fir st day I started. For her humour, patience, optimism and friendship over the last fe w years - it has all been greatly
appreciated and will never be forgotten.
I am very grateful to my supervisor at Pfizer, Dr. Christine Williams fo r the opportunity to study in deepest, darkest Kent. Her knowledge and help with those experiments, and valued input in all other aspects o f the work were invaluable. The near heart attack in her back garden after dinner nearly set this back a fe w years, but thanlfully I survived! Thanks also to Jo and Bernie fo r their hospitality in Broadstairs, you made me fe e l very welcome and I hope I can repay the fa vou r some day. (You can
keep the ghost cat down there with you though!)
For advice and assistance with all molecular biology work, my thanks to Dr. Stephano Casalotti. From the endless PCR runs in the beginning, to the disappointments with in situ, you were always there and always optimistic. Additional thanks fo r your input when this thesis was being written. Good luck to Italy in 2004!
Many thanks to Graham Neville fo r all the technical help and fo r making the lab an enjoyable place to work. The latest story on bringing up children was always guaranteed to raise a smile on the worst o f days! You've only about another 15 years to go.
For all their help and expertise with the in vivo Physiology, I am extremely grateful to Drs D avid McAlpine and Neil Ingham.
Thanks to all the girls in the lab - Jill, Victoria, Nerissa, Regina and Ruth fo r making the Institute a good place to be. Special thanks to Jill fo r being a good friend, fo r sharing the laughs and the bad days and fo r making me hope that Rovers have had a good game. I ’m afraid my heart will always lie with the mighty Liverpool F C though
(and Hibs o f course, dad).
Thanks to Dr. Karen D u ff and Dr. M ark B um s in New York fo r their understanding and advice when I was finishing the writing.
To my parents, I am indebted fo r all your support (financial and otherwise). From the bottle o f sauce delivered to the other end o f the country to the paper delivered to the other side o f the world, I am so grateful fo r everything you have done. You have always had fa ith in me, never let me give up on anything and have always been there when I have needed you. I appreciate it all and love you more than I ’m sure I tell you (so y o u ’ve got this in writing to remind me).
ABSTRACT.
The opioid receptor- like OP4 receptor and its endogenous ligand, nociceptin (NC),
have recently been isolated. Evidence suggests that the NC system m ay modulate
sensitivity o f the auditory system (Nishi et al., 1997).
The work presented in this thesis was undertaken to elucidate the role o f the NC system
in auditory function. NC is expressed in both central and peripheral auditory structures.
Furthermore, levels o f NC expression appear to correlate with the development and
maturation o f the auditory system, both in precocial and altricial animals, suggesting
that nociceptin expression may be related to auditory stimulation. To explore this
possibility further, NC expression was examined in models o f altered afferent input.
Results obtained suggest that NC is affected by the level o f acoustic stimulation and
may act via the olivocochlear efferent system to protect the cochlea from damage.
Evidence also suggests that NC is involved in auditory plasticity as long-term
alterations in NC expression were noted following unilateral deafening.
A differential regional and developmental pattern o f OP4 receptor splice variant
expression was observed, suggesting that the level o f receptor expression may be
related to development o f the auditory system and raising the possibility that different
variants m ay play specific roles in each tissue.
Pharmacological properties o f the OP4 receptor were determined in an OP4 receptor-
transfected CHO cell line. Peptides identified in this study were then used in in vivo
physiological experiments. These have shown that NC antagonists are able to reduce
the discharge rate o f binaurally-stimulated inferior colliculus neurones in response to
pure tones.
In conclusion, it appears that the NC system may play an important regulatory role in
neurotransmitters to modulate auditory sensitivity. These effects may be a result o f NC
receptor activation at the level o f the organ o f Corti and/or in more central auditory
TABLE OF CONTENTS
TITLE PAGE
ABSTRACT
CHAPTER 1: GENERAL INTRODUCTION.
The Inner Ear
The Organ o f Corti
Cochlear Innervation
Auditory Brainstem Nuclei
The Cochlear Nucleus
Auditory M idbrain
The Superior Olivary Complex (SOC)
The Inferior Colliculus (IC)
Function o f the Cochlea
The Olivocochlear Efferent System
Auditory Neurotransmitters
Endogenous Opioid Neuropeptides
Nociceptin
Effect o f the Nociceptin System on Neurotransmitter Release
Involvement o f Opioids in Modulating Auditory Function
Nociceptin and Auditory Function
CHAPTER 2: DISTRIBUTION OF NOCICEPTIN
DURING DEVELOPMENT. 39
2.1 Introduction 40
Development o f the Mammalian Auditory System 40
The Cochlea
(i) Anatomy 41
(ii) Neurochemistry 42
The Brainstem
(i) Anatomy 44
(ii) Neurochemistry 45
Postnatal Development 47
Distribution o f Nociceptin in the Auditory Pathway 47
Opioid Activity in the Auditory System 47
Aims 48
2.2 Materials and Methods 50
Animals 50
Tissue Preparation fo r Light Microscopy and Immunofluorescence
(i) Postnatal Animals 50
(ii) Prenatal Animals 51
Tissue Dissection 52
Preparation o f Tissue fo r Wax Sectioning 52
Preparation o f Tissue fo r Cryostat Sectioning 53
Transmission Electron Microscopy 53
Nissl Staining 54
2.3 Results 56
Nociceptin Labelling in the Adult Guinea Pig
The Cochlea 56
The Auditory Brainstem and Midbrain 57
Nociceptin Labelling in the Guinea Pig during Development 61
Nociceptin Labelling in the Developing Mouse Cochlea 66
2.4 Discussion 71
(i) The Mature Auditory System 11
(ii) The D eveloping Auditory System 73
What can be Concluded fro m these Results? 75
CHAPTER 3: ALTERATIONS OF AFFERENT INPUT AND
CHANGES IN NOCICEPTIN EXPRESSION 80
3.1 Introduction 81
General Effects o f Altered Auditory Input 81
(I) Unilateral D eafening 82
(i) Long Term Structural Effects 82
(ii) Neurochemistry 83
(Hi) Physiological Effects 84
(II) Acoustic Overexposure 85
(i) Anatomical Effects 85
(ii) Neurochemical Effects 86
(Hi) Physiological Effects 86
3.2 Materials and Methods 89
Animals 89
Unilateral Deafening 89
Otoacoustic Emission Recordings 90
Transiently Evoked Otoacoustic Emissions (TEOAEs) 90
Acoustic Overexposure 91
Tissue Preparation fo r Light Microscopy and Immunofluorescence 91
Tissue Dissection 92
Wax Sections 92
Cryostat Sections 92
N issl Staining 93
Immunofluorescence 93
3.3 Results 95
Control Sections 95
(I) Unilaterally Deafened Animals 95
6 Hours after Deafening 91
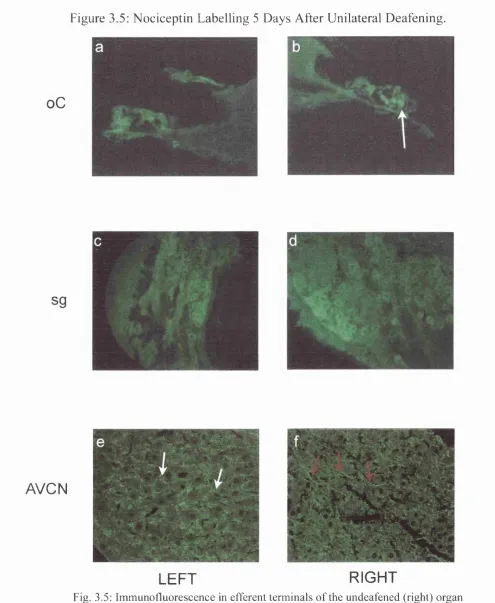
5 Days after Deafening 101
Two Weeks after Deafening 101
Five Weeks After Deafening 105
(II) A coustic Overexposure 109
3.4 Discussion 111
Anatom ical Effects o f Unilateral Deafening 111
Expression o f Nociceptin Following Unilateral Deafening
(ii) The Anteroventral Cochlear Nucleus 114
A coustic Overstimulation 115
Protective Effects o f the Olivocochlear System 115
CHAPTER 4: M OLECULAR BIOLOGY OF THE OP4
RECEPTOR AND NOCICEPTIN 117
4.1 Introduction 118
A natom ical Distribution o f the OP4 Receptor 118
OP4 Receptor Splice Variants 120
D ifferential Tissue Distribution o f OP4 Receptor Splice Variants 121
Functional Organisation o f the OP4 Receptor 121
M olecular Biology o f Nociceptin 126
Alternative Splice Variants o f Nociceptin 126
D ifferential Expression o f PreproNC Splice Variants 127
A im s 129
4.2 Materials and Methods 130
Animals 130
Tissue Dissection and Total RNA Extraction 130
RNA Preparation 130
Reverse Transcription PCR 131
RNA Precipitation 134
Gel Extraction and cDNA Purification 134
Expression o f OP4 Transcripts in A dult M ouse Tissues 139
Expression o f OP4 during Development 140
Expression o f N C and the OP4 Receptor in the Immortomouse Cell Line 146
Expression o f Nociceptin in the A dult M ouse Brain 147
4.4 Discussion 149
Expression o f OP4 mRNA in the Auditory System 150
Expression o f Prepronociceptin mRNA in the Auditory System 152
Alternative RNA Splicing in the Nervous System 154
CHAPTER 5: PHARMACOLOGICAL CHARACTERISATION
OF THE OP4 RECEPTOR 158
5.1 Introduction 159
Nociceptin and G-Protein Activation 159
Nociceptin: Structure-Activity Relationships 161
Active Fragments o f N C 162
Amination o f N C 162
[Tyr^] Nociceptins 163
Other Peptides o f Interest 164
O P4 Antagonists 164
The OP4 Receptor Ligand [F/G]NC(1-13)NH2 165
The Selective OP4 Antagonist [Nphe^]NC(I-13)NH2 165
In Vivo Effects o f OP4 Receptor Antagonists in the Auditory System 166
Binaural H earing and the Inferior Colliculus 167
5.2 Materials and Methods 169
(I) Unifilter Binding Assays 169
Cells and Culture Conditions 170
Preparation o f Cell Membranes 170
Determination o f Protein Concentration 171
Saturation Binding Assays 171
Competition Binding Assays 172
(II) Functional Assays 172
Cells and Culture Conditions 173
Plating Cells fo r FLIPR 173
Loading Transfected Cells with Dye 173
Preparation o f Peptide Plates 174
(a) Agonist Assays 174
(b) Antagonist Assays 175
(III) In Vivo Physiology 175
Animals 178
Stereotaxis and Recording 179
5.3 Results 180
(I) Binding
Determination o f Optimum Membrane Concentration 180
Binding o f [^HJNC to Transfected CHO-OPyGqiS Cells 180
Displacement Studies 180
(II) Functional Assays
Antagonist Assays 183
(III) In Vivo Physiology 189
5.4 DISCUSSION 192
Receptor Binding 192
Functional FLIPR Assays 193
Peptide Ligands
Nociceptin Fragments and Amination 195
[Tyr^] Nociceptin 196
Other Peptides o f Interest 196
Opioid Peptides 197
Antagonists 198
Effect o f [Nphe^]NC(1-13)NH2 on Auditory Neurones in vivo 199
CHAPTER 6: GENERAL DISCUSSION 202
The N C Ligand and A uditory Function 203
The OP4 Receptor in the Auditory System 207
Physiological Effects o f an OP4 Antagonist in vivo 209
H ow is N C Involved in Regulating Auditory Function ? 210
N C and Opioid Effects on Neurotransmitter Release 210
N C in the Cochlea 211
N C in the Auditory Brainstem 214
The Auditory Midbrain 215
Conclusions 219
Possibilities f o r Future Investigations 220
FIGURE REFERENCE LIST.
CHAPTER 1 PAGE
1.1 The M ammalian Cochlea 22
1.2 The Organ o f Corti 23
1.3 The Olivocochlear Efferent System 25
CHAPTER 2
2.1 Myelination o f Spiral Ganglion Cells during Development 43
2.2 Increase in Spherical Cell Size During Development 46
2.3 Electron M icroscopy Images o f Auditory Structures 58
2.4 Nissl Stained Sections o f Guinea Pig Auditory Structures 59
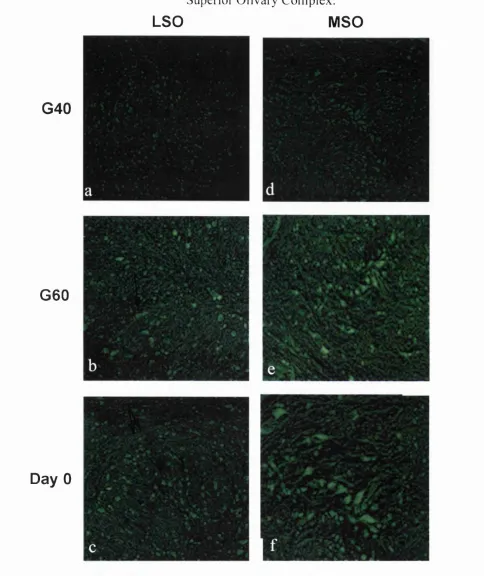
2.5 Nociceptin Labelling in the Mature Guinea Pig 60
2.6 Developmental Expression o f Nociceptin in the Guinea Pig Cochlea 62
2.7 Developmental Expression o f NC in the Guinea Pig Cochlear Nucleus 63
2.8 Developmental Expression o f NC in the Guinea Pig SOC 65
2.9 Postnatal Development o f the Mouse Organ o f Corti 6 8
2.10 Developmental Expression o f NC in the Mouse Organ o f Corti 69
2.11 Developmental Expression in Mouse Spiral Ganglion Cells 70
CHAPTER 3
3.1 Control Sections 96
3.2 Histology o f the Cochlea Following Unilateral Deafening 98
3.3 Histology o f the AVCN Following Unilateral Deafening 99
3.4 Nociceptin Labelling 6 Hours after Unilateral Deafening 100
3.5 Nociceptin Labelling 5 Days after Unilateral Deafening 103
3.7 Nociceptin Labelling 5 Weeks after Unilateral Deafening 106
3. 8 TEOAE Amplitude Following Unilateral Deafening 108
3.9 The Cochlea and Brainstem Following Acoustic Overexposure 110
CHAPTER 4
4.1 The OP4 Gene 124
4.2 Structural Representation o f the Nociceptin Receptor 124
4.3 Truncated OP4 Receptor Resulting from the Inclusion o f Intron 3 125
4.4 The PreproNociceptin Gene 128
4.5 Primers used in rt-PCR Experiments 135
4.6 Nociceptin Receptor Splice Variants 137
4.7 Sequence Alignment 138
4.8 Expression o f OP4 mRNA 142-43
4.9 Expression o f Nociceptin mRNA in Adult Mouse Brain 148
CHAPTER 5
5.1 G-protein Activation and Deactivation 160
5.2 Determination o f Optimum Protein Concentration 179
5.3 Determination o f Kd 180
5.4 Competition Binding Assays 181
5.5 Functional Agonists Assays 187
5.6 Antagonist Assays 188
5.7 Effects o f [Nphe^]NC( 1-13)NH2 on Individual IC Neurones 190-91
5. 8 Determined versus Literature pKi Values 194
TABLES
PAGE
C H A P T E R 1
1.1 Opioid Receptors and their Ligands 34
C H A P T E R 4
4.1 Expression o f OP4 Receptor Variants during Development 144-5
C H A P T E R 5
5.1 Binding Study Summary 182
5.2 Evaluation o f Different Assay Conditions 185
5.3 Agonist Assay Summary 186
1
CHAPTER 1: GENERAL INTRODUCTION.
The auditory pathway is a highly integrated communication network, which transmits
incoming auditory information to the auditory cortex where we perceive and interpret
sound. The transduction o f sound from acoustic into neural signals occurs in the
cochlea. The transduced information travels via the cochlear nerve, is then subdivided
and projects tonotopically through the various nuclei that constitute the auditory
pathway (see below). All o f these nuclei have a role in modifying and analysing
incoming auditory information and are also subject to descending, efferent influences
from higher centres. Efferent feedback mechanisms can modulate sensitivity
throughout the system and are crucial in normal hearing, as well as in the development
and maintenance o f neural connections (Brugge et al., 1978; Favre and Sans, 1978;
Lenoir et al., 1980; Ginzberg and Morest, 1983).
The transduction and transmission o f auditory signals involves inhibitory and
excitatory mechanisms mediated by neurochemicals. Neurones communicate via the
release o f neurotransmitters, which can initiate action potentials in adjacent neurones.
In this way, auditory information is passed from one part o f the system to another.
Other chemicals react to environmental and intrinsic cues to regulate and adapt neural
transmission. These neuromodulators may also function as neurotransmitters, and can
act to excite or inhibit the release o f other neurochemicals (Fex and Altschuler, 1981;
Hoffrnan et al., 1983, 1984, 1985; Eybalin et al., 1984; Eybalin and Pujol, 1984;
Altschuler et al., 1984a,b, 1985a,b; Drescher and Drescher, 1991, 1992; Scholtz et al.,
Nociceptin, an opioid-like neuropeptide may play a role as an auditory neuromodulator,
regulating the responsiveness o f the system to auditory stimulation. Despite its
structural homology to other opioids, nociceptin binds specifically to its own receptor,
OP4. In OP4 receptor knockout mice, Nishi et al (1997) described a significant
elevation in the threshold o f the auditory brainstem response following exposure to
intense noise (105dB SPL, 60 minutes). Furthermore, there was a significant delay in
recovery in comparison to normal animals. This would suggest that the nociceptin
system plays an important role in modulating auditory sensitivity and as such is an
important area o f research.
Before it is possible to consider a specific role o f nociceptin in the auditory system, it is
necessary to have an understanding o f auditory function. The anatomy, physiology and
neurochemistry o f the auditory pathway from the cochlea to the inferior colliculus will
be described below, together with a general description o f the nociceptin system and its
possible involvement in auditory function.
The Inner Ear.
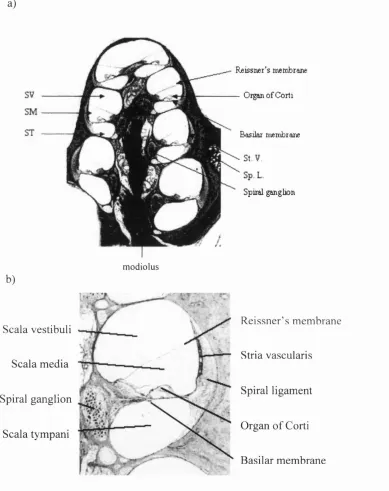
The inner ear is a fluid-filled bony structure embedded deep in the temporal bone o f the
skull. It contains the semicircular canals, the vestibule and the cochlea. The cochlea
consists o f a coiled canal, divided longitudinally into three scalae, spiralling around the
central modiolus. The scala media contains the potassium rich endolymph. It is
separated fi"om the scala vestibuli above by Reissner’s membrane and from the scala
tympani below by the basilar membrane together with the osseous spiral lamina
The ossicles in the air-filled middle ear transfer sound vibrations from the tympanic
membrane, causing a displacement o f fluids in the cochlea. The pattern o f displacement
exhibited by the mammalian cochlear duct takes the form o f a travelling wave with
several unique features (von Bekesy 1951). This wave is unidirectional, originating at
the base and travelling towards the apex o f the cochlea. It exhibits a peak o f maximum
displacement at a specific location along the cochlear duct that is strictly determined by
the fi-equency o f the acoustic stimulus; high frequency stimulation causes maximum
displacement near the base o f the cochlear duct, lower frequencies at progressively
more apical locations. This allows the generation o f a cochlear frequency-place map,
which underlies the ability o f mammals to analyse a spectrum o f sounds. This
cochleotopic and tonotopic organisation is maintained throughout the auditory
pathway.
The organ o f Corti.
Acoustic transduction within the cochlea takes place in the organ o f Corti, a complex
epithelial ridge that protrudes into the scala media and rests on the basilar membrane
throughout the length o f the cochlea duct (Figures 1.2a and 1.2b).
Two main categories o f cell are found in the organ o f Corti. There are the cochlear hair
cells, sensory receptors which transduce acoustic into neural signals. In contrast, the
supporting cells do not respond to acoustic stimulation but provide support,
surrounding and underpinning the auditory receptors.
Cochlear hair cells are capped, on their apical surface, by several rows o f stereocilia.
H air cells can be divided into two morphological and functional types, inner hair cells
shaped with a centrally located nucleus. The contractile outer hair cells may contribute
to mechanical feedback processes, playing an important role in maintaining normal
frequency selectivity and hearing sensitivity. (Kiang et al., 1970; Ryan and Dallos,
1975; Cody and Russell, 1985). These are cylindrical in shape, with a nucleus located
at the base o f the cell.
There is typically one row o f IHCs and between three and five rows o f OHCs, with a
human ear containing approximately 15000 hair cells. A shearing motion between the
tectorial membrane and the reticular lamina stimulates the hair cells. This movement
brings about a deflection o f the stereocilia bundles at the apical surfaces o f the hair
cells, resulting in the generation o f a receptor potential. The stereocilia o f the OHCs are
embedded within the tectorial membrane, whereas IHC stereocilia are mechanically
F ig u r e 1.1 T h e M a m m a lia n C o c h le a
a)
Rjeissner’s membrme
Oïgan o f Corti
Basilar mertibrarie
Spiral ^ g b o n
m odiolus
b)
Scala vestibuli
Scala media
Spiral ganglion
Scala tympani
Reissner’s membrane
Stria vascularis
Spiral ligament
Organ of Corti
Basilar membrane
Fig. 1.1a: A midmodiolar section through the guinea pig cochlea. The scala vestibuli (SV), scala media (SM) and scala tympani (ST) can be seen, separated from each other by the basilar membrane and Reissner’s membrane. The three scalae spiral together to form four turns. The organ o f Corti can be seen sitting on the basilar membrane. Also labelled are the modiolus (Mo), stria vascularis (St. V) and the spiral ligament (Sp. L). Taken from Slepecky(1996, Fig 2.1).
F ig u r e 1.2. T h e O r g a n o f C o r ti.
a)
Outer hair
c e lls C e lls o f H ensen
T ectorial m em brane
Inner hair c ell
O
b)
T ectorial m em brane
Supporting (Deiters’ cells)
\
Irrher hair cell Outer hair cells
mm
Fig. 1.2a: Cross sections of the organ of Corti. One row of inner hair cells and three rows of outer hair cells can be seen, together with a variety of supporting cells. The entire structure sits on the basilar membrane and is covered by the fibrous tectorial membrane. The stereociliary bundles of the outer hair cells can be seen to make contact with the tectorial membrane. Taken from Pickles (1988, Fig. 3 .ID).
Cochlear Innervation.
Inner hair cells are the primary transducers o f sound waves, whereas outer hair cells are
involved in regulating the responsiveness o f this sensory organ (Kiang et al, 1970;
Ryan and Dallos, 1975; Cody and Russell, 1985). The difference in function is made
more apparent if we consider their pattern o f innervation.
Approximately 20 radial afferent fibres contact each IHC. These Type 1 afferent
neurones are bipolar, their peripheral process projecting to the hair cells and the central
process to the cells o f the cochlear nucleus in the brainstem. Outer hair cells are
contacted by few, convergent. Type II afferents, each neurite branching to innervate ten
or more OHCs. Therefore, the great majority (approximately 93%) o f the cochlear
spiral ganglion neurones innervate the IHCs only, whereas about 7% innervate OHCs
(Spoendlin, 1972; Morrison, Schindler & Wersall, 1975). The IHC afferents are
myelinated central to the habenula and are o f larger diameter than the unmyelinated
OHC afferents, suggesting they conduct nerve impulses more rapidly. Axons from both
types o f afferent arise from the type I and type II ganglion cells lying in Rosenthals
canal. Both types o f afferent run via the cochlear nerve to the cochlear nucleus. An
efferent pathway parallels the ascending pathway throughout.
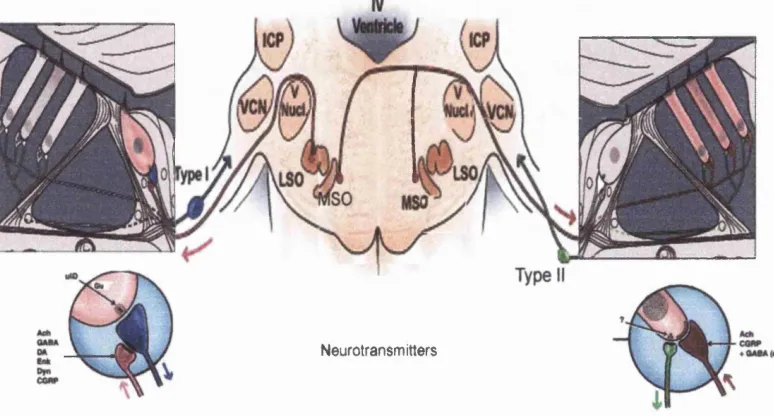
The olivocochlear bundle comprises the efferent innervation o f the cochlea. The nerves
o f this system can be anatomically divided into two components (W arr and Guinan,
1979; Warr, 1992). The somata o f the lateral olivocochlear efferent system (LOC) are
found in the lateral superior olive (LSO). They project mainly to the ip si lateral cochlea
where their thin, unmyelinated axons synapse with radial auditory dendrites below
inner hair cells. Medial olivocochlear efferent fibres (MOC) originate from neurones
their axons projecting to the contralateral cochlea (the crossed olivocochlear bundle),
the remaining 30% to the ipsilateral ear. There, they synapse directly with the basal
pole o f the outer hair cells (Fig 1.3).
Figure 1.3: The Olivocochlear Efferent System,
Neurotransmitters
Figure 1.3: The olivocochlear efferent system. The main efferent pathways from the auditory brainstem nuclei to the organ o f Corti are shown. As can be seen there are two main divisions o f this system. The lateral efferent system arises from the LSO and projects to the ipsilateral cells. The medial efferent system arises mainly from die MSO and projects to the contralateral cochlea, where the fibres innervate the outer hair cells. The main neurotransmitters involved are also shown.
VCN: ventral cochlear nucleus, LSO: lateral superior olive, MSO: medial superior olive, ICP: inferior cerebellar peduncle, IHC: inner hair cell, OHC: outer hair cell.
Auditory Brainstem N uclei
The Cochlear Nucleus
The cochlear nuclear complex (CNC) is the first nucleus o f the central auditory
pathway where the neural integration o f acoustic information occurs. It comprises the
anteroventral (AVCN), posteroventral (PVCN) and dorsal (DCN) cochlear nuclei and
plays a role in encoding binaural differences in the time and intensity o f acoustic
stimuli. The cochleotopic projection o f afferent fibres in different nuclear divisions
leads to an orderly arrangement o f the characteristic frequencies o f the neurones they
innervate. Axons tuned to high frequencies are located dorsally, with low frequency
axons occurring more ventrally.
In the ventral cochlear nucleus there are three major classes o f neurones: spherical,
octopus and multipolar cells. Spherical cells are found in the rostral pole o f the AVCN.
These are contacted axosomatically by one or more ascending auditory nerve fibres,
which form a chalice like structure, the end bulb o f Held. Spherical cells project to the
ipsilateral superior olive and then to the medial superior olivary complex o f both sides.
There are three layers in the dorsal cochlear nucleus: the molecular, granular and deep
layers. Small intrinsic excitatory neurones, the granule cells, send axons from the
granular layer into the outer molecular layer. In the granular layer, fusiform or
pyramidal cells receive input from afferent cochlear nerve fibres. They send projections
to the central nucleus o f the contralateral inferior colliculus, and are believed to have an
important role in binaural processing (Kitzes, 1978; Moore, 1991). Giant or multipolar
cells in the deep layer also project to the inferior colliculus, although some project to
connections with the pyramidal cells, the granule cells and each other. They also
project axons to the inferior colliculus and the AVCN.
The Auditory Midbrain,
The Superior Olivary Complex (SOC).
The superior olivary complex is located in the midbrain at the level o f the lower pons.
In mammals, there are three main nuclei: the lateral and medial nuclei o f the superior
olive (LSO and MSO) and the medial nucleus o f the trapezoid body (MNTB). The LSO
and M SO are surrounded by the smaller cell groups o f the pre- and peri-olivary nuclei
(Rasmussen et al., 1946; Elverland, 1978).
The LSO receives both ipsi- and contra-lateral connections from the VCN and
indirectly from the VCN via the MNTB. The region as a whole is predominantly
responsive to direction o f a sound source.
The M SO comprises bipolar neurones arranged in parallel layers. The medial dendrites
receive contralateral input from the AVCN, the lateral dendrites receiving the
ipsilateral input. This is the first level o f the auditory system at which there is
integration o f information from the two ears onto individual neurones. This integrated
information is transferred to other cells within the SOC for further processing and to
higher centres, the lateral lemniscus and inferior colliculus, as well as back down to the
CNC and cochlea. The main role o f the MSO is to respond to disparities in interaural
The Inferior Colliculus (IC).
The IC can be subdivided into a central nucleus (IC) an external cortex (EC) and a
dorsal cortex (DC).The central nucleus has a lamellar organisation. It is the main link in
the ascending auditory pathway and is reciprocally connected with the contralateral
central nucleus. The cochleotopic organisation o f the auditory pathway is maintained in
the IC, with high CF units found ventromedially and low ones dorsolaterally. The IC is
multi-sensory, receiving ascending input from somatosensory afferents together with
auditory input from the central nucleus and lower auditory centres. Almost all
brainstem auditory nuclei terminate in the inferior colliculus en route to the thalamus
and auditory cortex. Each region o f the IC is connected with its contralateral
counterpart, enabling information to be integrated across the parallel ascending
pathways.
Function o f the Cochlea,
There are three main elements o f cochlear function: the sound-induced motion o f the
basilar membrane, the transduction o f this motion by the inner hair cells into a neural
signal, and the neuro-mechanical feedback introduced by the outer hair cells. It is
possible that the nociceptin system may be involved in this feedback response in some
way.
The Olivocochlear Efferent System,
The organisation o f the olivocochlear efferent system has been described in some detail
previously. In this section, the main function o f this system will be discussed.
The absolute sensitivity o f the ear is determined by the minimal acoustic energy
determined, primarily, at the level o f the outer hair cells by fibres o f the olivocochlear
efferent system (Guinan, 1996 review).
Both the inner hair cell and its afferent fibres are susceptible to acoustic
overstimulation, which results in an elevated threshold to sound. Threshold shifts can
be temporary or permanent and are known to correlate with structural changes in the
stereocilia, cuticular plate, hair cell body, synaptic region o f the hair cell and post-
synaptic afferent nerve terminals (Saunders et al, 1985 review). Electrophysiological
evidence (Galambos, 1956) has determined that stimulation o f the efferent system can
alter the structure o f OHCs and their efferent terminals and also inhibits afferent fibre
activity. Furthermore, Dodson et al. (1986) electrically stimulated the afferent pathway
on one side via a round window electrode. The structural changes in OHCs and efferent
terminals observed in both cochleae were considered to have occurred through
feedback mechanisms via the crossed olivocochlear bundle (COCB).
Electrical stimulation o f the olivocochlear bundle has been shown to raise thresholds o f
primary afferent neurones within 100ms (Galambos, 1956; Wiederhold and Kiang,
1970; Guinan and Gifford, 1988a,b). Furthermore, this 'protective' mechanism is
blocked by systemically applied strychnine, a known blocker o f cholinergic efferent
synapses. This fast effect is thought to arise from the hyperpolarisation o f OHCs by
calcium influx through nicotinic receptors. This acts to decrease their amplification of
basilar membrane motion and hence decreases stimulation o f IHCs (Reiter and
Liberman, 1995). The OHCs aid the sharp tuning o f the IHCs via what is believed to be
an amplification o f the travelling waves along the basilar membrane (Ashmore and
be responsible for the increased timing to the basilar membrane. Thus, the medial OC
efferents would exert their effect mechanically. Sridhar et al (1995) reported an
additional slower response o f sound-evoked nerve activity that is also efferent-
modulated. They propose that the slow effect is attributable to release o f calcium from
the subsurface cistemae o f the OHC, causing calcium-gated potassium channels to
open and leading to hyperpolarisation o f the hair cell on a much larger time-scale
(Sridhar et al, 1997). Reiter and Liberman (1995) believe that protection against
temporary threshold shifts caused by high-level tones is correlated with the OC slow
effect. Efferent stimulation also reduces the level o f otoacoustic distortion products
(Mountain, 1980; Siegel and Kim, 1982). Otoacoustic emissions (OAEs) are low-
intensity sounds, produced specifically by the cochlea and, most probably, by the
cochlear outer hair cells as they undergo fast, reversible length changes. Changes in
their amplitude as a result o f acoustic overstimulation reflect structural and fimctional
damage to the hearing organ.
In summary, the olivocochlear system provides a mechanism by which the inner ear
can (i) improve the detection o f signals in noise: olivocochlear stimulation suppresses
continuous background noise, (ii) be protected fi*om noise damage, due to changes
elicited in the mechanical response o f the cochlea and (iii) control the mechanical state
o f the cochlea. This may be used to adjust for the effects o f changes in the cochlea
during aging, thus maintaining a relatively constant mechanical state in the cochlea.
Auditory Neurotransmitters
Neurones transmit their spikes to other neurones over their axon and contact other
neurones through synapses. M ost synapses release neurotransmitters into the cleft
presynaptic cell. The neurotransmitter binds to specific receptors in the postsynaptic
cell, causing specific ion channels to open. The type o f neurotransmitter involved
depends on the type o f synapse, with its effect determined by the specific class o f
receptor and its cellular effectors (discussed in more detail in Chapter 5).
The principal inhibitory neurotransmitters in the auditory system are y-aminobutyric
acid (GABA) and glycine, which open chloride channels in the post-synaptic cell,
eliciting inhibitory post-synaptic potentials (IPSPs) and in most cases hyperpolarising
the cell.
In the mammalian cochlea, glutamate is the excitatory afferent neurotransmitter.
Glutamate is both spontaneously released by IHCs, and released in response to
stimulation. Released glutamate binds to both N-methyl-D-aspartate (NMD A) and non-
NMDA receptors in type I auditory neurones (Puel, 1995). These fibres, which
innervate the modiolar IHC surface, are contacted by a relatively large number o f
lateral efferent terminals.
Neurochemically, the lateral and medial olivocochlear systems are quite distinct. It is
reported that the medial efferent system is nearly exclusively cholinergic, with
calcitonin gene-related peptide (CGRP) and GABA most probably used as additional
neurotransmitters and/or neuromodulators (Eybalin and Pujol, 1987, Fig 4). At the level
o f lateral efferent terminals, acetylcholine (ACh), GABA and CGRP have again been
proposed, with the additional involvement o f dopamine and the opioid peptides
enkephalins and dynorphins (Fex and Altschuler, 1981; Usami et al, 1988a,b; Gil-
(co)-release different neurotransmitters depending on certain physiological and/or
pathological conditions (Drescher et al, 1983; Altschuler et al, 1984a, b, c).
Endogenous Opioid Neuropeptides,
Although substantial evidence indicates that ACh is the main neurotransmitter for
medial and lateral OC efferents, these fibres also use neuroactive opioid peptides.
The term ’opioid' is used to describe all substances that are opium- or morphine- like in
action, and retain their capacity to modulate sensory input via their binding to
designated (opioid) receptors (Sahley and Nodar, 1994). These substances include
synthetic and natural drug alkaloids as well as the neuroactive endogenous enkephalin-
and dynorphin-related peptides (Simon and Hiller, 1978 a,b). Three main classes o f
opioid receptor have been identified, p, ô and k , which are activated by morphine-,
enkephalin- and dynorphin- related peptides respectively (Evans et al., 1992; Kieffer et
al., 1992; Chen et al., 1993; Minami et al., 1993).
Further attempts to clone additional opioid receptor subtypes led to the isolation o f
cDNA encoding a homologous protein with a high degree o f sequence similarity to the
opioid receptors, but did not bind any existing opioid peptides with high affinity. This
'orphan' opioid receptor has now been identified from human (O R L l, Mollereau et al,
1994), mouse (MOR-C, Pan et al, 1995) and rat genomic libraries (LC132, X O R l, C3,
ROR-C, oprl and H yp8, Bunzow et al, 1994; Fukuda et al, 1994). In accordance with
recent UIPHAR recommendations, the term OP4 will be used to describe the receptor
In 1995, two groups simultaneously reported the isolation o f the natural ligand for OP4.
This peptide neurotransmitter was biologically related to the opioids, but heightened
sensitivity to pain when injected into the brains o f mice. Hence, M eunier et al (1995)
named it nociceptin (NC). As the receptor was found before the ligand, Reinscheid et al
(1995) referred to the peptide as orphanin FQ, the peptide for the ‘orphan receptor’, and
because its amino acid sequence begins with phenylalinine (F) and ends with glutamine
(Q) (discussed in more detail in chapter 5).
Nociceptin,
Like the endogenous opioid ligands, NC is derived from a larger precursor,
prepronociceptin (preproNC). The nucleotide sequence o f the preproNC gene has
similar sequence characteristics and organisation to the opioid peptide precursors,
particularly preproenkephalin and preprodynorphin, suggesting these peptides may
share a common evolutionary ancestor. However, despite the structural similarities
between the opioid and NC systems, they are pharmacologically distinct. As a general
rule, NC does not bind to opioid receptors and opioid ligands have no affinity for the
OP4 receptor (Table 1.1). However there have been recent studies suggesting that
opioid peptides may bind with specific subtypes o f the nociceptin receptor (Mathis et
Table 1.1: Opioid Receptors and their Ligands.
R ecepto r N am e O Pi O P2 O P3 O P4
Previous name Ô K O RLl
Coupling Gi/o Gi/o Gi/o Gi/o
Natural ligands Leu-enkephalin
Met-enkephalin
Dynorphin
Dynorphin (1-8)
P-endorphins
endomorphins
nociceptin
Synthetic
agonists
DPDPE Spiradoline DAMGO NC(1-13)NH2
Selective
antagonists
Natrindole Nor-BNI CTOP [Nphe‘]N C
(l-13)NH2
Non-s elective
antagonists
Naloxone Naloxone Naloxone Naloxone
insensitive
Table 1.1: G protein-coupled opioid receptors and their specificity. Opioid ligands do
not bind to the OP4 receptor, just as nociceptin does not bind to any opioid receptors.
(Data from Corbett et al, 1982; Zadina et al, 1997; Guerrini et al, 1998; Calo et al,
Effect o f the Nociceptin System on Neurotransmitter release.
In the central nervous system, nociceptin inhibits the release o f neurotransmitters such
as dopamine (Murphy et al., 1996), glutamate (Nicol et al., 1998), GABA (Meis and
Pape, 2001), acetylcholine (Neal et al., 1997), noradrenaline (Schlicker et al., 1998)
and serotonin (Sbrenna et al., 1999). Similarly, in the periphery, NC inhibits the release
o f neurotransmitters from sympathetic (Nicholson et al., 1998), parasympathetic (Calo
et al., 1996) and peptidergic nerves (Giuliani et al., 1997; Rizzi et al., 1999 a,b). NC
has been shown to inhibit dopamine release in anaesthetised rats when administered
intracerebroventrically ((icv), M urphy et al., 1996). However, it has also been shown to
increase dopamine release in the striatum o f conscious rats (Konya et al., 1998),
indicating its effects on neurotransmitter release are not clear-cut. NC also has a
biphasic effect on enkephalin release. Low doses (1-lOnM) inhibit, whereas high doses
(lOO-lOOOnM) stimulate release.
In various preparations, NC has been reported to inhibit neurotransmitter release due to
a reduction in excitability o f the presynaptic neuronal elements and/or a reduction in
presynaptic neurotransmitter secretion (Calo et al., 2000). NC may act by inhibiting
Ca^^ channels within the nerve terminals, activating presynaptic K^ channels or through
direct modulation o f the release apparatus, usually thought to result from the
spontaneous exocytosis o f transmitter-containing vesicles occurring in the absence o f
Ca^"^ influx (Miller, 1998). Meis and Pape (2001) have reported that NC inhibits the
release o f GABA and glutamate in the rat lateral amygdala by such presynaptic
mechanisms.
suggests that nociceptin, like the classical opioids may be a very important
neuromodulator. In the auditory system, little has been published regarding the
distribution o f nociceptin or its receptor. However, it has been shown in other systems
that nociceptin and the opioids are found in parallel, yet distinct fibre routes. This may
also be true in the auditory pathway (discussed in more detail in chapter 2).
Involvem ent o f Opioids in M odulating Auditory Function,
Neuroactive enkephalins and dynorphins typically produce inhibition within the
nervous system (Sahley and Nodar, 1994). Opioid receptors can exist pre- or post-
synaptically (Cherubini and North, 1985; Reisine and Bell, 1993; Hiller et al., 1997).
Evidence for the existence o f opioid receptors within the mammalian cochlea has been
determined mainly from stereospecific binding studies (Hoffinan, 1986) and the
observation that inhibition o f receptor-linked adenylate cyclase in the cochlea is
reversible by naloxone, a specific opioid antagonist (Eybalin et al, 1987 a,b).
There is a growing amount o f evidence to suggest that opioid-containing OC neurones
are involved in modifying auditory function. Sahley and Nodar (1994, 1996) present
results that indicate that dynorphin peptides contained within lateral efferent OC
neurones act via k - opioid receptors to improve auditory sensitivity to low intensity
stimuli. Interestingly, interactions between opiates and cochlear acetylcholine receptors
have been shown in Xenopus auditory organs by Lioudyno et al. (2000). There is
evidence that opioids may also be acting at the level o f the auditory brainstem
(Mansour et al, 1994; Kungel and Friauf, 1995; Code, 1996; Code and McDaniel,
neurotransmitters such as acetylcholine to exert their effect on hearing sensitivity
(Sahley and Nodar, 1996).
Nociceptin and Auditory Function,
Further evidence for an opioid-mediated effect on hearing has been shown using
nociceptin-receptor knockout mice. Nishi et al (1997) reported that after exposure to
intense sound homozygous ORL-1 knockout mice showed a significant increase in
auditory brainstem response as compared to baseline readings (average: 37dB SPL) and
with heterozygous mutants and wild-type controls. This threshold shift returned to
normal levels within 3 days and was therefore deemed temporary.
The auditory system is able to adjust following injury or sensory receptor loss to
function in ways that can often compensate for these losses. For example, auditory
ftmction is impaired by unilateral deafening, noise-induced hearing loss or by nerve
damage resulting from acoustic over-exposure. Under such circumstances, the 'plastic'
properties o f the auditory system allow the reorganisation o f neurones formerly
involved in areas that have been damaged (discussed in more detail in Chapters 2). This
reorganisation o f neurones is not fully understood, and can often result in unwanted
side effects, such as mislocalisation, hyperacusis and tinnitus (Kaas, 1995).
Opioid peptides may be involved in such physiological states as hyperacusis and
tinnitus. Opioids are involved in the overall biological response to stress (Akil et al,
1984) and can also enhance the regulatory functions o f other neurotransmitters (Lewis
et al, 1987 a,b). During stress reactions opioid peptides, specifically dynorphins, may
NM DA receptors in the cochlea. Consequently, the spontaneous activity generated
during periods o f quiet may be altered or amplified, producing symptoms o f tinnitus.
Nociceptin is known to be heavily involved in pain pathways and mechanisms
throughout the CNS. A pathophysiology has been suggested between chronic pain and
chronic tinnitus (Tonndorf, 1987; Moller, 1997, 2000). In these models, both pain and
tinnitus consist o f neural sensitisation, involving a reduction in threshold and/or an
increased neural excitability to stimuli. In this case, intense sound would exacerbate
tinnitus and repeated exposure to such stimuli in tinnitus sufferers may lower auditory
tolerance to all sounds (Gabriels, 1996; Jastreboff, 1996).
As has been discussed above, initial evidence supports a role for the nociceptin system
in the modulation o f auditory function (Nishi et al., 1996). This is supported by
localisation studies, which have shown that nociceptin is highly expressed in auditory
brainstem nuclei (Anton et al., 1996; Boom et al., 1999; M ollereau et al., 2000,
Kakimoto et al., 2001). There has been no further published information regarding the
possible involvement o f nociceptin in auditory function.
The results presented in this thesis describe research undertaken to elucidate the
possible role o f nociceptin and its receptor in the auditory system. This has been
achieved with several studies, each concerned with one particular aspect o f the
2
Distribution o f Nociceptin
CHAPTER 2: DISTRIBUTION OF NOCICEPTIN DURING DEVELOPMENT.
2.1 INTRODUCTION.
During development, neurones are often functional before they become fully mature, a
process dependent on both excitatory and inhibitory influences (Kaas, 1996). For
example, release and binding o f main auditory neurotransmitters such as ACh have
been determined in several brainstem auditory nuclei prior to maturity. If nociceptin
plays a role in auditory function, it may also contribute to these developmental
processes. Therefore, by examining NC immunoreactivity during development, insights
may be provided into possible functions o f NC in the auditory system.
Two species o f animal have been used in this study. Guinea pigs are precocial, like
man they begin to hear in utero and undergo a sudden exposure to a more complex
acoustic environment at birth. At this time, there is a major shift fi*om fluid and tissue
conducted sound to air-home noise, thus involving exposure to a very different sound
spectrum. Conversely, the altricial mouse is bom in an immature, deaf state, resembling
the auditory system o f a human embryo o f 15 weeks (Kikuchi and Hilding, 1965). With
the onset o f auditory function not occurring until postnatal day 1 2 (P I2), this species
allows for the study o f early developmental processes.
Development o f the mammalian auditory system.
An understanding o f the major events occurring during maturation o f the auditory
system is cmcial to the interpretation o f the results o f any developmental study. A brief
description o f development o f the neural circuitry o f the brainstem and cochlea,
In both mice and guinea pigs, various levels o f the auditory system develop
simultaneously. Some processes occurring during development are intrinsic to the
tissue while others depend on the functioning o f synaptic inputs. During the initial
stages o f maturation the auditory neurones are generated, neurites migrate to their adult
locations, axons are sent out, dendrites begin to differentiate and synaptogenesis
begins. Programmed cell death also occurs during this time. In this way, the adult
pattern o f circuitry is established at an early stage o f development.
The Cochlea (i) Anatomy
There are two types o f adult mammalian cochlear hair cells. Inner hair cells (IHCs)
primarily transduce incoming auditory stimuli into neural signals. Outer hair cells
(OHCs) are believed to be involved in feedback mechanisms, protecting the inner ear
from acoustic overstimulation. These different functions are reflected in their pattern o f
innervation, with IHCs having predominantly afferent innervation, and OHCs with a
dense medial olivocochlear efferent innervation. This is not, however, always the case.
As soon as IHCs begin to differentiate they are contacted with nerve endings o f radial
type I spiral ganglion neurones, as well as lateral olivocochlear fibres. Around the onset
o f hearing, spiral ganglion neurones become myelinated (Figure 2.1) and there is an
increase in the number and size o f afferent nerve endings at the basal pole o f IHCs
(Romand et al, 1980; D odson and Mohuiddin, 2000). The efferent fibres decrease in
number, with the remaining efferent terminals contacting afferent dendrites. It has been
suggested that this early efferent innervation may act to protect the afferent fibres from
W hen OHCs begin differentiation they are contacted exclusively by spiral afferents
(Romand, 1983). However, a short time after the onset o f cochlear function there are
very few afferent endings around them, with the cell contacted by many efferent
terminals. The adult appearance o f efferent synapses has been described as one o f the
final events in the maturation o f cochlear sensory and neural structures (Pujol, Carlier
and Lenoir, 1980).
(ii) Neurochemistry.
Auditory neurotransmitters such as ACh and GABA are both present in the cochlea
well before the onset o f cochlear function (Lenoir, Shnerson and Pujol, 1980; Whitlon
and Sobkowicz, 1989; Merchan-Perez et al., 1990), perhaps suggesting a role for these
efferent neurotransmitters in cochlear synaptogenesis (Pujol et al, 1979). It has been
hypothesised that, as in other systems, these neurotransmitters may play a role in the
formation and maintenance o f ACh synapses (Pujol and Puel, 1999). Finally, opioid
peptides, such as met-enkephalin have been detected in the inner spiral bundle around
the time o f the onset o f cochlear function (Pujol and Hilding, 1973). This presynaptic
localisation implicates opioid peptides in efferent neurotransmission processes. Such
early involvement o f neurotransmitters is believed to provide trophic support for
postsynaptic neurones and may contribute to the stabilisation o f specific neuronal
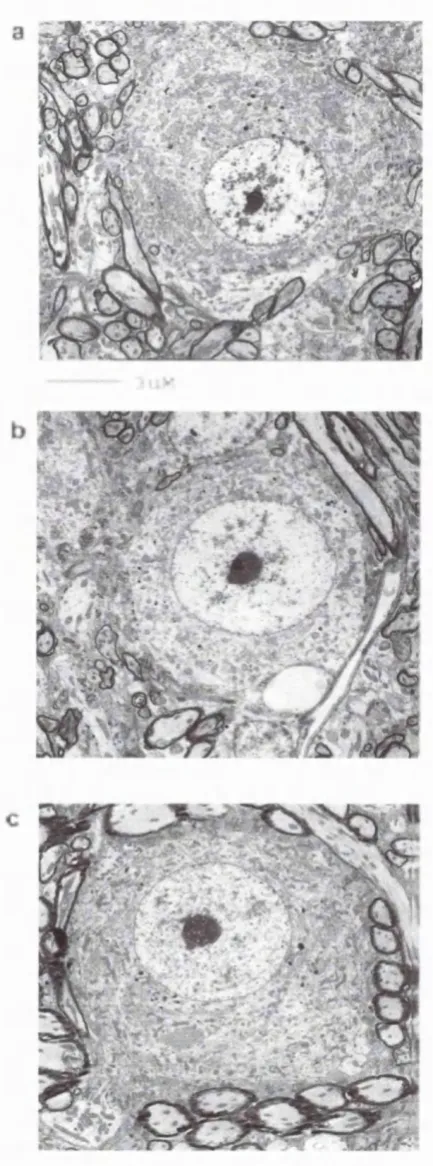
Figure 2.1: Myelination o f Guinea Pig Spiral Ganglion Cells
During Development
%
2 m icroiis
Fig. 2.1. Arrows indicate site of myelination. There is no myelination o f spiral ganglion cells at gestational day 40 (040, a). At 06 0 , the approximate time of onset o f hearing, loosely packed myelin is apparent around ganglion cells (b). As the guinea pigs age,
the myelin compacts and appears darker (c & d).
The Brainstem (i) Anatomy
Even in highly altricial species, the brain stem circuitry at birth has patterns o f
connections characteristic o f the adult. Both the radial afferent and spiral afferent
fibres, comprising the auditory nerve, enter the brain stem while the neurones o f the
future cochlear nucleus are still being generated. Similarly, axons o f the cochlear
nucleus (CN) reach the superior olivary complex (SOC) and inferior colliculus (IC) at a
time when genesis o f the neurones that will populate these higher auditory centres is
still underway (Altman and Bayer, 1980; Repetto-Antoine and Meininger, 1982;
Kandler and Friauf, 1993). M any o f these new connections already have a clear
tonotopic organisation along the presumptive hearing axis (Moore, 1982; Henkel and
Brunso-Bechtold, 1993).
Gleich et al. (1998) observed that following the onset o f hearing the antero ventral
cochlear nucleus almost doubles in size. Further studies by Dodson (1986) have shown
increases in spherical cell size in this region, both before and after the onset o f cochlear
function in guinea pigs (Figure 2.2). Kudo et al (1988, 1990) demonstrated that within
the lateral superior olive (LSO), neurones projecting ipsilaterally and contralaterally are
generated at different times, ipsilateral connections being generated earlier. In the
medial superior olive (MSO), the dendrites develop their characteristic bipolar
orientation some time before the onset o f hearing (Henkel and Brunso-Bechtold, 1990).
Other evidence suggests that early connections are made between the brainstem and
cochlea via the olivocochlear efferent fibres. Even after the onset o f cochlear function,
it appears that an ongoing process o f change and rearrangement o f these neurones
(ii) Neurochemistry.
Electron microscopy and physiological techniques have shown that spherical cells in
the cochlear nucleus have synaptic contact with both the endbulbs o f Held and the
inhibitory GABA-positive terminals before hearing begins (Kane and Habib, 1978;
Brugge and O'Connor, 1984; Wu and Oertel, 1987; Schweitzer et al, 1993). Studies
have also shown that both GABA receptors and glutamate-responsive receptors are
functional at this stage. Thus, excitatory and inhibitory receptors are functional before
the circuitry is completely operational (Walsh et al., 1987, 1990, 1993).
In the adult, several immunohistochemical studies have shown the presence o f
endogenous opioids within these efferent neurones (Wamsley et al, 1980). Both
proenkephalin and prodynorphin gene products are found prominently in the lateral
efferent terminals innervating eighth nerve dendrites under mature IHCs (Fex and
Altschuler, 1981; Hoffman et al, 1983, 1984, 1985; Altschuler et al, 1984, 1985;
Eybalin et al, 1984a,b 1987; Drescher et al., 1992; Scholtz et al, 1998). Further studies
provide evidence that these two opioid peptide families co-exist within the same
neurones (Altschuler et al, 1986; Abou-Madi, 1987). The distribution o f opioid
peptides within the medial efferent system is less clear. Recently Ryan et al (1991)
found no enkephalin-ir in lateral OC neurones o f the gerbil, with h alf o f the medial OC
population positively labelled, suggesting there may be two, biochemically distinct
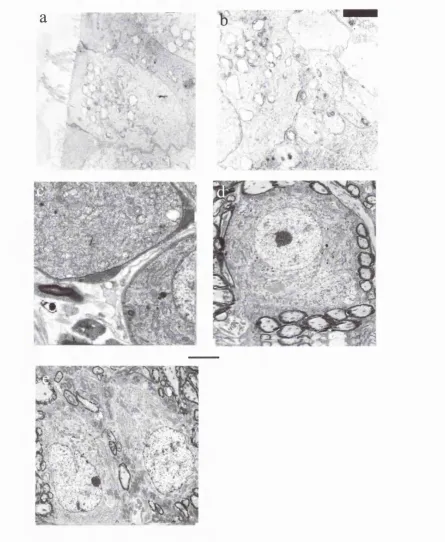
Figure 2.2: Increase in Guinea Pig Spherical Cell Size During Development.
Fig. 2.2: Spherical Cells in the antero ventral cochlear nucleus. Spherical cells increase in size between 0 4 0 (a) and 0 6 0 (b). There is no further increase in size before birth (c). (EM Plates were provided by Dr. Hilary Dodson, ILO).
Postnatal Development,
The most obvious maturational change after hearing has begun is the rearrangement
and growth o f dendrites, together with the axonal arbors that innervate them. There also
appears to be an environmental cause for the refinement o f axonal connections from the
CN to areas such as the IC. There is much evidence to suggest that an intact cochlea is
important for the normal development o f such axons (Sanes and Takacs, 1993; Moore
and Kitzes, 1985; Moore and Kowalchuk, 1988; Moore, 1990, 1994) and the
maintenance o f cell size. In this way, at the onset o f hearing, the auditory system o f all
mammalian species has reached essentially the same stage (Morey and Carlile, 1990),
with the basic structures in place and basic patterns o f circuitry established.
Distribution o f Nociceptin in the Auditory Pathway,
There is relatively little information concerning the distribution o f nociceptin in these
structures. Nociceptin mRNA expression in the brainstem was examined by Boom et
al (1999). M oderate to high levels o f prepronociceptin mRNA were expressed in the
inferior colliculus, lateral lemniscus, periolivary nuclei and superior olivary complex.
Houtani et al (1996) also described a moderate to high distribution o f NC in the IC,
superior olive and periolivary region. More recently, Kakimoto et al (2001) identified
nociceptin mRNA positive cells in several auditory brainstem nuclei, such as the dorsal
and ventral nucelei o f the lateral lemniscus, the periolivary nuclei and the inferior
colliculus. The distribution does appear to be quite similar to that o f the opioids. As yet,
there is no published data showing NC expression in the cochlea.
Opioid Activity in the Auditory System.
enkephalin-containing olivocochlear neurones discharge with noise stimuli.
Furthermore, Sahley et al (1996a, b) used a selective k-opioid receptor antagonist, Nor-
Bni, to show the reversibility o f opioid- mediated neural effects in the cochlea. The
same group has also shown the non-selective opioid antagonist, naloxone, has similar
effects in blocking the opioid receptor- mediated actions o f (-) pentazocine in the
cochlea. Pentazocine alone was shown to improve auditory sensitivity (Sahley and
Nodar, 1994).
Results obtained from localisation studies can often be used as an indicator for
physiological actions. For example, the NC system is known to be involved in pain
perception mechanisms and Boom et al (1999) reported that NC mRNA expression was
detected in areas implicated in descending antinociceptive pathways, such as the
gigantocellular nucleus and raphe magnus nucleus. Similarly, Boom et al (1999)
observed high NC mRNA expression in association with auditory relay nuclei such as
the inferior colliculi, lateral lemniscus nuclei, medioventral preolivary nucleus and the
superior olivary complex. It was observations similar to these that prompted Nishi et al
(1997) to investigate the hearing ability o f NC receptor knockout mice. Following
exposure to intense noise there was a significant increase in the threshold o f the
auditory brainstem response in NC knockout mice compared to that in controls. These
experiments provided the first evidence that the NC system is involved in modulating
auditory function.
Aims,
Although, evidence shows that NC is found in more central auditory nuclei, so far there
the expression o f NC in the auditory pathway during development. The aims o f this
study were to determine:
1. The distribution o f NC in the adult mouse and guinea pig.
2. Changes occurring in distribution during development that could be associated with
hearing function in
a) the altricial mouse, which has postnatal development o f hearing,
2.2 MATERIALS AND METHODS.
Animals.
CBA/ca mice (2-lOg) and pigmented guinea pigs (40-400g) bred in the Animal Facility
o f Biological Services, University College London, were used. Animals were housed in
a room maintained at 23 ± 2°C and a 12-hour light/dark cycle (light on at 6:30 A.M.).
Food and water were available ad libitum. The treatment o f animals was in accordance
with the Animals (Scientific Procedures) Act 1986.
Developmental and maturational changes in NC expression were examined in guinea
pigs at gestational day 40 and 60 (G40, G60), the day o f birth (PO) and at three
postnatal weeks (mature). The distribution o f NC in mice was examined from
embryonic day 15.5 (E15.5) and at 2-day intervals until 4 postnatal weeks. Two
animals were used per group.
Tissue Preparation fo r Light Microscopy and Immunofluorescence.
(i) Postnatal Animals
Guinea pigs and mice were deeply anaesthetised with an intraperitoneal injection o f
sodium pentobarbital (Reckitt and Coleman, Hull, UK, 0.05mg kg'^ body weight).
Once the rib cage was completely exposed, bone scissors were used to cut each side o f
the ribcage, lateral to the midline, allowing the mid-portion o f the ribcage to be raised,
exposing the underlying organs. A needle was inserted into the left ventricle o f the
heart. This was held in place using a butterfly clip. A small incision was made in the