Early Development and Wound Healing in the
Zebrafish Embryo
KATIE JANE WOOLLEY
A Thesis Submitted for the Degree of
Doctor of Philosophy
University of London
2001
ProQ uest Number: U 642453
All rights reserved
INFORMATION TO ALL U SE R S
The quality of this reproduction is d ep en d en t upon the quality of the copy subm itted.
In the unlikely even t that the author did not sen d a com plete manuscript
and there are m issing p a g e s, th e se will be noted. Also, if material had to be rem oved, a note will indicate the deletion.
uest.
ProQ uest U 642453
Published by ProQ uest LLC(2015). Copyright of the Dissertation is held by the Author.
All rights reserved.
This work is protected against unauthorized copying under Title 17, United S ta tes C ode. Microform Edition © ProQ uest LLC.
ProQ uest LLC
789 East E isenhow er Parkway P.O. Box 1346
ABSTRACT
In this thesis I use the Zebrafish embryo as a model to study tissue movements of embryonic wound healing as a model of natural morphogenetic movements that shape the embryo during development. In particular I am interested in the signals that initiate embryonic tissue movements, the signals that stop them, and the cytoskeletal machinery that drives them.
Re-epithelialisation of a wound is accomplished by a concerted set of cell shape changes and rearrangements which drive the epithelium over the wound area. I have used the Zebrafish embryo to study these cell shape changes both by Scanning Electron Microscopy (SEM) and in real time using time-lapse analysis of vitally-stained embryos, and made direct comparisons with the analogous natural morphogenetic tissue movement of epiboly. The cell shape changes of re-epithelialisation appear to be driven by contraction of an actin/myosin purse string which draws the epithelium forward. I have examined re organisation of the actin cytoskeleton in leading edge cells during both wound closure and epiboly using phalloidin-staining to reveal filamentous actin and have used a specific inhibitor of a key effector of the small GTPase Rho, Rho-kinase, to test its function in directing these key actin polymerisation events. I have also analysed reorganisation of the microtubule network in wound edge cells and report that while microtubules reorientate following wounding and are essential for epiboly, they are not required for initiation or maintenance of epithelial movements.
TABLE OF CONTENTS
ABSTRACT... 2
TABLE OF CONTENTS... 3
TABLE OF FIGURES...10
ABBREVIATIONS...11
ACKNOWLEDGEMENTS... 12
CHAPTER ONE... 13
INTRODUCTION... 13
Me c h a n is m s o ft is s u e r e p a ir... 13
The fibrin clot and infiammatory cells are key signals in the adult wound... 13
Re-epithelialisation occurs by lamellipodial crawling... 14
Wound edge connective tissue cells proliferate and spread into the wound defect... 15
Adult wounds result in scar tissue whereas embryonic tissue repair is perfect... 16
The inflammation response and growth factor profiles differ markedly between the adult and embryonic situation... 17
Embryonic wound re-epithelialisation is by actinomyosin purse-string contraction rather than adult-like lamellipodial crawling... 18
Purse-string formation is dependent on small GTPases... 19
Re-epithelialisation is accompanied by contraction o f underlying mesenchyme.. 21
Em b r y o n i c w o u n d e p i t h e l i a l i s a t i o n m a y b e a n a l o g o u s t o e m b r y o n i c
EPITHELIAL MORPHOGENETIC M OVEM ENTS... 2 2
Ventral enclosure in C.elegans is partially driven by a contracting actin
purse-string...23
Drosophila gastrulation involves coordinated actinomyosin contractions by large groups o f epithelial cells... 25
Dorsal closure during Drosophila embryo genesis involves spreading o f an epithelial sheet over a contracting amnioserosa substrate...28
The Zebrafish Danio rerio is an excellent model fo r studying cell movements o f wound healing and morphogenesis in vivo... 30
Genetic screens in Zebrafish have led to the isolation o f numerous mutant lines.. .30
Epiboly may be similar to re-epithelialisation... 32
Wo u n d in g st u d ie s in h s h... 35
A historical perspective... 35
Even relatively large skin lesions in adult fish close within minutes... 36
Studies in Fundulus embryos show that both EVL movements and migration o f deep cells occur during repair... 38
Molecular clues from fin regeneration studies... 39
CHAPTER TWO...41
MATERIALS AND METHODS...41
Maintenance o f Zebrafish... 41
Observation o f live embryos... 41
Wounding and culturing o f epiboly stage embryos... 42
Wounding and culturing o f 24 hour and later embryos... 42
Processing embryos fo r scanning electron microscopy (SEM)... 43
Preparing embryos fo r phalloidin staining... 44
Live staining o f embryos with flore scent vital dyes... 44
Preparing embryos fo r immunostaining o f microtubules... 45
Treatment o f embryos with specific inhibitors... 45
Imaging software fo r fixed tissue and time-lapse movies... 46
Testing the role o f Calcium during wound closure... 47
Immunostaining fo r anti-phospho-histone-3... 47
Whole-mount immunocytochemistry fo r anti-active ERK on Zebrafish embryos... 48
Whole-mount immunocytochemistry fo r anti-active p38 and anti-active JNK on Zebrafish embryos... 48
Treatment with MAP kinase inhibitors... 49
CHAPTER THREE... 50
CYTOSKELETAL CHANGES DURING WOUND HEALING IN THE
ZEBRAFISH BRACHYDANIO RERIO... 50
In t r o d u c t io n... 5 0 Embryonic epithelial wound front cells assemble an actin cable... 50
Actin cables are also observed in tissue culture and repair o f some adult tissues.. 51
Formation o f the purse-string is dependent on the small GTPase Rho... 53
Actin purse-strings are found in single cell wounds... 55
Assembly o f the actin cable in Xenopus oocytes correlates with a microtubule network... 56
Re s u l t s... 58
Wounds made to the 24 hour fish heal rapidly by epithelial cell shape changes. ...58
Wounds made to the 50% epibolyembryos heal in a standardised fashion 59 Timelapse studies reveal the dynamic cell shape changes during wound closure.. 60
A contractile actin cable assembles within minutes o f wounding in the 50% epiboly
embryo and pulls the wound closed... 62
Assembly o f a wound actin purse-string is dependent on Rho-kinase, a downstream effector o f the small GTPase Rho... 63
Microtubules reorganise in wound edge cells but are not required fo r re-epithelialisation o f the wound... 64
Di s c u s s i o n... 7 4 Cells at the wound edge undergo a reproducible programme o f cell shape changes and movements...74
What signals might initiate these actin-driven cell shape changes?...75
Membrane remodelling may play a role in re-epithelialisation...77
What triggers Rho activity?...78
How does this machinery switch off when two edges meet?... 79
How is the hole finally sealed closed?...80
What role do microtubules have during wound closure?... 81
Similar sets o f concerted cell shape changes occur during some epithelial morphogenetic processes... 83
CHAPTER FOUR... 85
A COMPARISON OF WOUND CLOSURE AND EPIBOLY OF
ZEBRAFISH EMBRYOS...85
In t r o d u c t io n... 85
Epiboly is the process whereby an epithelial sheet moves over the yolk o f the teleost embryo... 85
Is wound healing re-using morphogenetic movement machinery?... 88
The driving force o f epiboly may lie in the YSL... 89
Endocytotic events may also play a key role in EVL epiboly... 93
Cell proliferation may be repressed during morphogenetic tissue movements 95
Several mutants are available which may allow a genetic approach to identify
genes controlling epiboly and gastrulation...95
A comparison o f wound healing and epiboly... 96
Re s u l t s... 9 8 Epiboly and wound closure superficially resemble one another... 98
Cells at the blastoderm margin are seen to be rujfling and extending filopodia. .100 Proliferation is absent throughout epiboly but commences immediately thereafter... 100
TEM reveals very different characteristics o f these two movements...101
The leading edge o f a wound and the epiboly edge both show actin redistribution... 103
Microtubules, whilst not necessary fo r wound healing, are pivotal fo r epiboly.... 104
Weg embryos show a delayed wound healing response... 104
Dis c u s s io n... 119
The role o f cell shape changes and shufflings versus cell proliferation in closure o f an epithelial hole... 119
Cells at the blastoderm margin ruffle as they are pulled together... 121
Epiboly and wound closure may be driven by different cytoskeletal mechanisms... 122
What role does actin play in epiboly o f the Zebrafish embryo?... 122
What can wounding studies on mutants tell us?... 123
CHAPTER FIVE... 125
SIGNALLING EVENTS DURING WOUND CLOSURE IN
ZEBRAFISH EMBRYOS... 125
In t r o d u c t i o n... 125
What signals kick-start the embryonic wound process?... 125
Entry o f calcium into damaged wound edge cells may be a pivotal signal during wound closure... 126
Various immediate early genes are upregulated at the embryonic wound site.... 127
Growth factor expression at the embryonic wound site... 129
MAP kinases are activated and may play a role in wound healing... 129
Tissue culture studies show lEG induction downstream o f calcium entry and MAP kinase activation... 132
Various antibodies and specific inhibitors are available which test the role o f MAPK activation during in vivo wound healing...133
Re s u l t s...135
A wave o f calcium spreads back from the wound edge following wounding 135 The rate o f wound closure depends on extracellular calcium concentration.... 135
ERK is rapidly and transiently activated during wound healing in 7 hour and 24 hour embryos... 136
ERK activation is not required fo r wound closure... 137
p38 and JNK cascades appear not to be specifically activated during wound healing... 138
High concentrations o f SB230580 does not affect wound closure... 138
A cocktail o f U0126 and SB203580 does not affect wound closure... 139
Di s c u s s i o n... 145
Calcium plays an important role in wound healing... 145
CHAPTER SIX... 150
GENERAL DISCUSSION AND FUTURE DIRECTIONS...150
What we know about wound healing in the Zebrafish embryo... 150
What other signals may direct cable assembly ?... 153
Do epithelial movements repress cell division?... 155
New techniques will allow a more detailed study o f the role o f calcium during wound healing... 157
What do the Zebrafish mutagenesis screens tell us so fa r?...159
Zebrafish is entering a 'post-genomic' era... 160
Transgenic fish will provide new insights into dynamic processes underlying wound repair... 163
Other morphogenetic movements may be more analogous to wound healing than epiboly...165
Does embryonic wound healing have anything to tell us that is relevant to adult clinical repair?... 166
BTBTJOGRAPHY... 169
APPENDIX T... 187
TABTÆ OF FIGURES
Figure 1.1 Cartoon to show how an epithelium closes up 19 Figure 1.2 Cartoon showing ventral furrow formation in Drosophila 26 Figure 1.3 Crossing scheme used in Tübingen 32 Figure 1.4 Schematic of epiboly 34 Figure 3.1 Wound closure in the 24 hour Zebrafish embryo 66 Figure 3.2 Making wounds and wound closure in epiboly-stage embryos 67 Figure 3.3 Cell shape changes during wound closure in epiboly-stage
embryos 68
Figure 3.4 Epithelial relaxation following wound closure 69 Figure 3.5 Actin-cable assembly in wound edge cells 70 Figure 3.6 The effect of Rho-kinase inhibitor on wound closure 71 Figure 3.7 No actin-cable forms in embryos treated with Rho-kinase inhibitor 72 Figure 3.8 Microtubules in wound closure 73 Figure 4.1 Schematic illustration showing the different layers in a Zebrafish 86
embryo at the beginning of epiboly
Figure 4.2 Schematic of involution vs ingression 88 Figure 4.3 Schematic of microtubules in epiboly embryos 90 Figure 4.4 Diagrammatic representation of the dynamic microtubule
networks found in the Zebrafish embryo during epiboly 91 Figure 4.5 Cell shape changes appear similar during wound healing and
epiboly 106
Figure 4.6 Cell shape changes during the final stages of epiboly 107 Figure 4.7 Cells further back from the blastoderm margin retain a polygonal
shape 108
Figure 4.8 The YSL is covered in microspikes 109 Figure 4.9 The YSL projects long filopodia over the EVL, YSL and yolk 110 Figure 4.10 A burst of proliferation follows the completion of epiboly 111 Figure 4.11 No cell division occurs in cells actively changing shape during
epiboly 112
Figure 4.12 No cell division occurs in elongating wound edge cells 113 Figure 4.13 Histology of 80% epiboly embryo 114 Figure 4.14 Histology of wounded 50% epiboly embryo 115 Figure 4.15 Actin distribution in EVL cells and YSL at blastoderm margin 116 Figure 4.16 Microtubules during epiboly and wounding 117 Figure 4.17 weg embryos show a slow wound-healing response 118 Figure 5.1 MAPK cascades 130 Figure 5.2 Cells at the wound edge show an increased level of intracellular
calcium following wounding 140 Figure 5.3 Wounds heal at different rates depending on the external media 141 Figure 5.4 ERK immunostaining of wounds made to the 24 hour Zebrafish
tail 142
Figure 5.5 Active-ERK expression in wounds made to epiboly embryos and
ABBREVIATIONS
AER Apical ectodermal ridge A P I Activator protein-1
bFGF Basic fibroblast growth factor Ca^+ Calcium ion
CPAE cells Calf pulmonary aortic endothelial cells CTGF Connective-tissue growth factor CLSM Confocal laser scanning microscope DIG Differential interference contrast DNA Deoxyribonucleic acid
DnDRac Dominant-negative Drosophila Rac Dpp Decapentaplegic
ECM Extracellular matrix
EDTA Ethylenediaminetetraacetic acid
EGTA Ethylene glycol-bis(p-aminoethylether)-N,N,N’ ,N’-tetraacetic acid EGF Epidermal growth factor
Egr-1 Early gene response-1 ENU Ethylnitrosourea
ERK Extracellular regulated kinase ESTs Expressed sequence tags EVL Enveloping layer
FGF Fibroblast growth factor FITC Fluorescein isothiocyanate lEG Immediate early gene GFP Green fluorescent protein GTPase Guanosine triphosphatase
mc-6
Intestinal epithelial cell line-6 JNK Jun N-terminal kinaseJNKK Jun N-terminal kinase kinase KGF Keratinocyte growth factor MAPK Mitogen-activated protein kinase MEK MAPK effector kinase
MMP Matrix metalloproteinase PDGF Platelet-derived growth factor SEM Scanning electron microscopy SRE Serum response element SRF Serum response factor
ACKNOWLEDGEMENTS
I would like to thank, first and foremost, my supervisor Paul Martin for all his support and encouragement, patience and good humour, throughout my time at UCL, both concerning my PhD work and all other matters I’ve thrown at him. I’d also like to thank everyone in the lab over the years for help and friendship. It would be unfair to highlight any group of people as everyone has helped in some way, whether it be from help with experiments in the lab or comforting in the pub. However, I have to say this thesis would not be in its present form without computer help from Antonio and proof reading by Will.
I’m also grateful to Mary Rahman for all her molecular advice, Darren Clements for help in the confocal suite, Brian Gasking for providing embryos every day and Mark Turmaine for EM expertise and windsurf tuition.
I’ve been guided into the fish world by a lot of people, mainly from the UCL Zebrafish facility, headed by Steve Wilson,
Chapter 1 Introduction
CHAPTER ONE
Introduction
Wound healing is crucial to all organisms. Any lesion made to the skin must be rapidly repaired to protect the body from the external environment and risk of infection. The process of healing a wound involves the re-covering of the wound by surrounding epithelium and the filling in and contraction of connective tissue defects. In many respects, this process is not dissimilar to various morphogenetic events that take place during natural development to shape the embryo. Therefore it is not surprising that embryos are exceptionally good at healing wounds: indeed in embryos, the process is rapid, efficient and is scar-free. For these reasons wound healing is an important process to investigate. Not only can the tissue movements of embryonic wound healing be used as a model of the natural morphogenetic movements that shape the embryo during development, but in addition we can use information gleaned from embryonic wound healing to help us understand the events happening during adult tissue repair.
MECHANISMS OF TISSUE REPAIR
* The fibrin clot and inflammatory cells are key signals in the adult wound
Chapter 1 Introduction
mechanisms underlying this repair process, and the collaborative efforts of the many different tissues involved at the wound edge, are now becoming clear. Because the skin acts as a protective barrier to the external environment, any damage must be rapidly repaired. This is initially achieved by the temporary formation of a fibrin clot, comprising platelets embedded in a mesh of cross-linked fibrin fibres with plasma fibronectin, vitronectin, and thrombospondin. This clot has a dual function: it initially protects exposed wound edges and provides a rough matrix through which cells can migrate during the healing process; and in addition releases cytokines and growth factors which recruit inflammatory cells to the wound site, and stimulate the subsequent tissue movements of repair (Martin, 1997). Inflammatory cells, such as neutrophils and macrophages, act at the wound site to clear contaminating bacteria and cell and matrix debris, but also to amplify earlier wound signals by the release of further pro-inflammatory cytokines and growth factors.
• Re-epithelialisation occurs by lamellipodial crawling
Chapter 1 Introduction
metalloproteinases (Mignatti et al., 1996). These proteases also aid in degrading components of focal adhesion attachment to the dermal substratum, allowing the cells to disengage from their substrate and to migrate forwards. Once re-epithelialisation is complete and the wound is closed, a whole array of events proceed; growth factors and proteases which were upregulated following wounding are now down-regulated, a basal lamina is now resynthesised, and new adhesion complexes between the epithelium and the basal lamina are established.
• Wound edge connective tissue cells proliferate and spread into the wound defect
C hapter 1 Introduction
• Adult wounds result in scar tissue whereas embryonic tissue repair is perfect
Despite most adult skin lesions healing fairly rapidly the end result is not perfect - a connective tissue scar persists at the wound site and epidermal appendages, such as sweat glands and hair follicles, which were originally present at the site of damage, do not regenerate.
Chapter 1 Introduction
• The inflammation response and growth factor profiles differ markedly between
the adult and embryonic situation
One key difference between the adult and embryonic situation appears to be the absence of an inflammatory response in embryos (Hopkinson-Woolley et al., 1994), as will be discussed in a leter section, with TGF-pi being the key switch involved. In the embryonic wound T G ppi is expressed and released transiently at the wound site but rapidly cleared before the wound is closed (Martin et al., 1993), whereas in the adult wound situation TG ppi is delivered to the wound site by degranulating platelets and invading macrophages and is expressed at high levels throughout the repair process. In the adult it is likely that TGpp acts as a chemotactic cue initially, and then induces extracellular matrix production, as is likely in the embryonic situation. However the rapid clearance of TGPp from the embryonic wound site seems to suggest that this may be why embryos do not over-contract or scar. Addition of exogenous TGPP to healing rat incisional wounds increases tensile strength and production of granulation tissue. However this is implicated in fibrosis which is involved in scar tissue formation. Anti-TGppi antibodies administered to the adult wound site reduce fibrosis which subsequently reduces scarring indicating that this is a main signal leading to scar tissue formation (Shah et al., 1995).
C hapter 1 Introduction
• Embryonic wound re-epithelialisation is by actinomyosin purse-string contraction
rather than adult-like lamellipodial crawling
Chapter 1 Introduction
actin cable
adherens junction
disorganised actin
Figure 1.1 C artoon to show how an epithelium closes up (B rock et al., 1996). Cells at the
w o u n d e d g e (b lu e ) a sse m b le an a c tin -c a b le at th e ir le a d in g e d g e (g re e n ), lin k e d
in te rce llu la rly by ad h erens ju n ctio n s (black). C o ntraction o f this cable causes cells to
c o n stric t at th eir leading edge (cell A) and ad h erens ju n c tio n s to m ove so that cell A
eventually m oves out o f the w ound edge. As the cells m ove, the actin cable shortens and
disassem bles leaving disorganised actin in the excluded cells.
• Purse-string formation is dependent on small GTPases
T he actin c a b le is a sse m b le d from o rie n ta te d b u n d led stre ss-fib re s. C lu es as to the
m olecular sw itches w hich m ight trigger cable assem bly of this in tracellular structure com e
from tissue cu lture studies in 3T3 fibroblasts w here the sm all G T P ase m o lecu lar sw itch
R ho has been show n to m ediate stress fibre assem bly (R idley and H all, 1992). Indeed
Chapter 1 Introduction
C hapter 1 Introduction
• Re-epithelialisation is accompanied by contraction of underlying mesenchyme
As during adult wound closure, connective tissue contraction plays a pivotal role in embryonic repair but unlike adult contraction there is no conversion of fibroblasts into myofibroblasts at the wound site (McCluskey and Martin, 1995). This may be because reduced levels of growth factors at the embryonic wound site are insufficient to trigger conversion to the adult-stage myofibroblasts, or because embryonic fibroblasts are more active and contractile than their adult counterparts and can exert sufficient tensile force to close the wound without conversion to myofibroblasts. In an organ culture model of fetal rat skin wounds, connective tissue contraction, like re-epithelialisation, can be blocked by cytochalasin B and is therefore dependent on the actin cytoskeleton (Ihara and Motobayashi, 1992).
• There is little inflammation at the embryonic wound site but various potential “kick-start” genes are upregulated
Chapter 1 Introduction
play key roles in embryo repair. For example, TG Fpl is upregulated at the embryonic wound site during wound closure but is only transiently expressed and is cleared before the wound is fully closed (Martin et al., 1993). This local expression of TG Fpi may be downstream of the rapid induction of the transcription factor c-fos which is transiently upregulated within minutes by the front 4 or 5 rows of epithelial cells at the wound (Martin and Nobes, 1992). Many other members of the API transcription factor family are also upregulated at the wound site (Okada et al., 1996). It is likely that the induction of immediate early genes leads to transcriptional activation of downstream genes, such as growth factors and cytokines, which are directly responsible for triggering the tissue movements of wound closure.
EMBRYONIC WOUND EPITHELIALISATION MAY BE ANALOGOUS TO EMBRYONIC EPITHELIAL MORPHOGENETIC MOVEMENTS
Chapter 1 Introduction
• Ventral enclosure in C.elegans is partially driven by a contracting actin purse-string
Epiboly in the worm, Caenorhabditis elegans, involves the bilateral spreading of an epithelial sheet from the dorsal side of the embryo past the equator to the ventral midline. This process, known as ventral enclosure, resembles re-epithelialisation during vertebrate embryo wound closure. The epidermis, also called the hypodermis, originates from a set of cells on the dorsal side of the embryo. Two pairs of anterior hypodermal cells lead the lateral migration of this epithelia towards the equator by extending actin-rich filopodia and crawling downwards stretching the hypodermis away from the dorsal side. These cells appear to migrate in the same way that adult wound edge kératinocytes crawl forward. However, once these four leading cells have made contact the entire epithelial leading edge assembles an actin-purse string and this contractile purse-string draws the ventral hole closed. At the ventral midline, the cells halt and form a seam by establishing epithelial junctions with cells from the opposite side. Laser inactivation of the leading cells immediately after ventral enclosure has begun causes either permanent retraction of the hypodermis or serious delay in the process (Williams-Masson et al., 1997). Even after epiboly is complete and the ventral midline cells have fused and formed junctions, ablation of these cells results in rupture of the embryo, showing how important this strong contiguous epithelium at the midline seam is.
Worms mutant for hmr-1 (encoding cadherin), hmp-2, (encoding 6-catenin) and hmp-1,
Chapter 1 Introduction
front row cells that is required for sealing of the contralateral epithelial fronts (Raich et al., 1999).
Both the initial filopodial extensions of the leading cells and the later purse-string movements towards the ventral midline are actin-dependent as shown by disruption of actin by cytochalasin D (Williams-Masson et al., 1997) but as yet there are no studies to test the role of Rho family small GTPases in these two steps of ventral enclosure.
C hapter 1 Introduction
• Drosophila gastrulation involves coordinated actinomyosin contractions by large
groups of epithelial cells
Chapter 1 Introduction
I
Ventral epitheliumactin c a b le
a d h e r e n s junction
F igure 1.2 C artoon show ing ventral furrow fo rm ation in D ro so p h ila. C ells w ithin the
ventral ep ithelium (top illustration) begin to elongate and co n strict at their leading edge,
driven by an a ctin -cab le. C o n strictio n o f this cable leads to in fo ld in g o f the ventral
C hapter 1 Introduction
The epithelial cells which first begin constricting their apices to drive gastrulation appear to be randomly located along the ventral midline, but within 10 minutes all the cells within this specific 12 cell-wide strip, have initiated apical constriction and the whole strip then invaginates into the mesoderm primordia within the embryo. Genetic analysis in
Drosophila has revealed the transcription factors twist, snail, huckebein, and tailless to be involved in these invaginations, twist and snail are expressed in the ventral cells at the site of ventral furrow invagination whilst huckebein and tailless control specification of the posterior and anterior limits of midgut invagination respectively. However, these four genes are involved in determining the area of invagination, rather than actually directing the cell shape changes. It is likely that genes downstream of these four will provide clues to the mechanisms of invagination. One gene involved in both ventral furrow formation and posterior midgut invagination is folded gastrulation, fog, whose expression precedes invagination in both cases. Fog mutant embryos show normal initiation of gastrulation with random cells constricting but the majority of ventral cells remain columnar and although ventral furrow formation eventually proceeds it is delayed and irregular in appearance and midgut invagination fails altogether. The concertina mutant, which encodes the a subunit of a G protein, shares a very similar phenotype, leading to the suggestion that a subset of cells constrict early in the ventral furrow and signal to neighboring cells to follow suit, with Fog operating as a candidate for the secreted signal. Activated Fog receptor then presumably activates the G -a protein Concertina, fo g and eta
Chapter 1 Introduction
• Dorsal closure during Drosophila embryogenesis involves spreading of an epithelial sheet over a contracting amnioserosa substrate
Towards the end of embryogenesis the Drosophila embryo undergoes dorsal closure whereby the lateral epithelium spreads dorsally to cover the amnioserosa and zips up at the dorsal midline. Genetic analysis has revealed this movement to comprise three stages: initiation which involves the elongation of front row cells of the lateral ectoderm, the spreading of the ectoderm which involves the elongation of the more laterally located ectodermal cells, and the zipping up of the two lateral sheets at the dorsal midline. The movement of dorsal closure involves forward movement of a free epidermal edge without cell recruitment or cell proliferation, only cell shape rearrangements, suggesting close analogy to wound closure. In addition, this epithelial movement is accompanied by assembly of an actin cable within the leading edge of the migrating epithelium. Flies mutant for zygotic non-muscle myosin, zipper, fail to complete dorsal closure. In this respect, the cellular machinery driving dorsal closure closely resembles that of embryonic re-epithelialisation of a wound.
A number of dorsal hole mutants in Drosophila have allowed dissection of the pathway of genes controlling dorsal closure. It is now clear that in the leading edge cells of the lateral epithelium a JUN kinase cascade is initiated which leads to activation of AP-1 (jun/fos). Mutations in this cascade, notably basket, which codes for a JNK, and its activator
hemipterous which encodes the homologue of vertebrate JNKK, fail to undergo dorsal closure, possibly due to failure of elongation of the leading edge cells. Activation of the JNK cascade also leads to expression of p u c k e r e d in the leading edge cells, and
C hapter 1 Introduction
Dpp acts by interacting with the Dpp receptors encoded by the genes thick veins and punt in the epithelial cells back from the leading edge, possibly triggering their elongation, as well as on the amnioserosa cells ahead of the leading edge, which may constrict in response to Dpp signals in a way analogous to the TGpp directed control of wound mesenchyme cells (Noselli and Agnes, 1999).
The idea that dorsal closure was driven solely by an actin purse-string around the advancing epithelial edges has recently been superceded by findings that implicate filopodia as a key regulator of this movement (Jacinto et al., 2000). Imaging of living fly embryos expressing GFP-tagged actin reveals dynamic filopodia and lamellipodia extending from the advancing epithelial fronts. These filopodia appear to 'sense' the opposing epithelial front in order to find the correct partner to fuse to, and appear to play a role in the final adhesion and fusion between the two opposing epithelial faces. In addition, expression of dominant-negative Cdc42 blocks filopodial extension and epithelial adhesion, as does blocking JNK signalling. Although dominant-negative Rho, Rac or Cdc42, all show some level of failure of dorsal closure, it now appears that DN-Cdc42-mediated effects resemble Dpp signaling components whereas DNRac share phenotypes with the JNK cascade mutant phenotypes (Ricos et al., 1999). It may be that different GTPases are required across the lateral epithelial cells for different processes such as contraction, adhesion and fusion (Stronach and Perrimon, 1999).
Chapter 1 Introduction
activated by stretch in several tissue culture models (Li et al., 1999). Another possibility is that a signal from the amnioserosa somehow triggers the leading edge cells closest to it, to activate small GTPase switches and the JNK cascade.
• The Zebrafîsh Danio rerio is an excellent model for studying cell movements of wound healing and morphogenesis in vivo
The Zebrafish Brachydanio rerio is a tropical freshwater cyprinid with many features that make it amenable for developmental, cellular and genetic analyses. The advantages of using Zebrafish for both sophisticated embiy'ological manipulations and as a genetic tool have been well documented in recent years (Driever et al., 1994; Stemple and Driever, 1996). Zebrafish are hardy and easy to care for in the laboratory, and inexpensive to maintain. Eggs are fertilised externally and embryos are completely transparent, allowing one to follow the development of each single cell. Fish embryos can develop at room temperature and are amenable to various cell manipulations. It is a simple matter to manipulate the extracellular environment that the fish develop in, and they can be observed at a greater range of stages than most other embryo models. These advantages allow visualisation of cell and tissue movements for extended developmental periods which are ideal for my planned wound healing studies.
• Genetic screens in Zebrafish have led to the isolation of numerous mutant lines
C hapter 1 Introduction
Chapter 1 Introduction
F1
F2
F3
ENU treatm ent of sp e r m a to g o n ia
fam ilies
+ / + X + / + + / + X -f/i
random m a tin g s
X +/+ + /@ x
< © =
< e = =
Figure 1.3 C rossing schem e using in Tubingen (H affter et al., 1996).
• Epiboly may be similar to re-epithelialisation
Z ebrafish lay large num bers of sm all eggs, about 1mm in diam eter, w hich are covered by a
tran sp aren t and relatively soft chorion. F o llo w in g fertilisation the cytoplasm o f the egg
stream s tow ards one pole o f the egg raising the blastodisc o f the one-cell egg to form a cap
C hapter 1 Introduction
Chapter 1 Introduction
A
EVL
d e e p c e lls
YSL
yolk cell
ep ib la st
h yp ob last
yolk cell
Figure 1.4 Schem atic o f epiboly (K ane et al., 1996). (A) S tructure o f the em bryo at 50%
epiboly. T he blastoderm com prises the en v eloping layer (purple) w hich covers the deep
cells (blue). The yolk syncytial layer (green) covers the anim al pole hem isphere of the yolk
cell (yellow ). (B) S tructure o f the em bryo at 80% epiboly. The Y SL m oves tow ards the
vegetal pole with the EV L follow ing. At the blastoderm m argin the deep cells involute to
form the hypoblast (red) and the non-involuting cells becom e the epiblast (blue).
T he driving force underlying epiboly is not com pletely u nderstood. C ells at the leading
edge o f the advancing EV L do not appear to extend protrusions suggesting that craw ling is
probably not the m echanism (B etchaku and T rinkaus, 1978). In stead it appears that the
E V L (purple) is draw n over the yolk by its association with the underlying yolk syncytial
layer (Y SL) (green) w hich form s from the collapse o f m arginal b lastom eres into the yolk
cell. T he Y S L ep ib o lises ahead o f the E V L and can c o m p lete epiboly even w hen the
blastoderm is rem oved (Trinkaus, 1951b). In F undulus, the en v eloping layer is attached to
the Y SL by tight ju n ctio n s (B etchaku and T rinkaus, 1978). It has been show n that UV
treatm ent o f zebrafish em bryos undergoing epiboly severely retarded or prevented epiboly,
C hapter 1 Introduction
separation of epiboly and gastrulation movements) can be mimicked by the microtubule- depolymerising drug nocadazole, suggesting that microtubules, and their associated motors, are the driving force behind epiboly. Microtubules can be seen radiating from the YSL to the yolk cytoplasmic layer, aligned in the direction of advancement, showing how microtubule motors might provide the force required for movement of the YSL over the yolk. This provides a mechanism for the YSL to be propelled over the yolk, but in addition a circumferential tension must be required for the constriction of the blastoderm margin as it nears completion of epiboly. This is probably actin-mediated as high amounts of actin have been detected in the external YSL (Betchaku and Trinkaus, 1978), and cytochalasin D, the actin depolymerising agent, prevented contraction of the blastoderm during epiboly in
Medaka (Sguigna et al., 1988). The fact that the advancing edge is not smooth suggests that tension may not be the only force acting and makes it unclear whether an actin purse- string operates at the leading edge of either the EVL or YSL, or whether cells are actively crawling at any point.
Several mutants which fail to complete epiboly or are defective in gastrulation movements were found in the Tübingen and Boston zygotic screens (Kane et al., 1996; Solnica-Krezel et al., 1996) and are discussed in detail in chapter 4.
WOUNDING STUDIES IN FISH
• A historical perspective
Chapter 1 Introduction
(1935), (reviewed in Mittal et al., 1977). Fin regeneration has been studied ever since and recently several groups have begun to dissect out the molecular mechanism underlying the repair process using the Zebrafish as a model..
During the 1970’s several authors contributed to describing the sequential histopathology of skin wounds in teleosts, culminating in work by Mittal et al (1977), who described the pattern of wound healing and overall structural changes in the skin and underlying muscles as a result of a skin wound in the adult Cat-Fish Heteropneustes fossilis (Bloch). Repair of the comeal epithelium has also been studied in marine and freshwater fish (Ubels and Edelhauser, 1982). A more recent paper (Quilhac and Sire, 1999) described healing of superficial skin wounds made to the adult cichlid fish, Hemichromis bimaculatus, detailing the spreading, proliferation and differentiation of the epidermis and the interactions between the epidermal cells and the underlying substrate.
It was not until 1988 that embryonic wound healing in fish first began to be studied using the killifish Fundulus heteroclitus (Fink and Trinkaus, 1988). Since this time no further work has been published in the field of embryonic fish wounds.
• Even relatively large skin lesions in adult fish close within minutes
re-Chapter 1 Introduction
epithelialised within 4 hours, at a rate of 1.25mm / hour but the underlying tissues are much slower at recovering. Granulation tissue forms from the blood capillaries and fibroblasts, and gradually resumes normal synthesis of collagen. It takes 35 days for the epidermis to resume its normal structure and appearance, and interestingly, there is no trace of a scar left at the wound site (Mittal et al., 1977).
A more recent study focused on the healing of superficial skin wounds in adult cichlids,
Hemichromis biaculatus (Quilhac and Sire, 1999). Again, re-epithelialisation was fast, with a 1cm square wound surface completely covered by the healing epidermis 9 hours after surgery. Although it takes longer for the epithelium to resume its normal structure there is no mention of whether a scar forms. Re-epithelialisation does not require proliferation of surrounding epithelium, as revealed by incorporation of tritiated thymidine, but there does appear to be a burst of proliferation following completion of re- epithelialisation. This lack of proliferation required for epithelialisation has been noted previously (Misof and Wagner, 1992). Therefore re-epithelialisation occurs by the spreading and migration of pre-existing cells from around the wound area. This spurt of new division seen in areas around the wound, especially in intermediate layers, lasts for a few days, indicating that new cell division is required in the restructuring of the tissue around the wound.
C hapter 1 Introduction
None of the studies above describe the mode of motility utilised by the rapidly repairing epidermis in adult fish wounds so it is unknown whether a crawling or a purse-string mechanism may be involved.
• Studies in Fundulus embryos show that both EVL movements and migration of deep cells occur during repair
Chapter 1 Introduction
controlling wound closure. To address these issues we have looked for clues from other systems.
• Molecular clues from fin regeneration studies
Teleosts have the capacity to regenerate fins following their amputation. Fin regeneration in zebrafish leads to re-establishment of full fin pattern and is preceded by upregulation of a battery of genes that may be important in the repair process. Fin regeneration proceeds through several stages similar to those which occur during urodele limb regeneration. Within 24 hours of fin amputation, a wound epithelium has migrated over the fin stump. It is in direct contact with the underlying mesenchyme and is thought to play a role similar to the epithelium of developing fin buds. This epithelium is believed to operate in a similar way to the apical ectodermal ridge (AER) of developing limbs in higher vertebrates because it is positioned at the exact boundary of ventrally located expression of engrailedl
gene (Ekker et al., 1992; Hatta et al., 1991). Also expression of sonic hedgehog (shh)
Chapter 1 Introduction
In a temperature-sensitive screen for mutations affecting growth and regeneration of the zebrafish caudal fin several mutant lines were identified in which fin regeneration fails, (Johnson and Weston, 1995). Characterisation of these mutants will provide more clues about the downstream signals involved and the genes required for patterning the regenerate, but is unlikely to shed light on the basic repair processes that are the topic of my thesis. No screens have yet been undertaken that address re-epithelialisation or closure of a wound in Zebrafish.
C hapter 2 G eneral M aterials and M ethods
CHAPTER TWO
General Materials and Methods
• Maintenance of Zebrafish
A breeding colony of zebrafish {Danio rerio) were maintained on a 14 hour light/10 hour dark cycle (Westerfield, 1993). Embryos were collected by natural spawning and staged according to Kimmel et al. (1995). Normal embryos were generated from the wild-type lines kcwt (King's College wild-type), uwt (University College wild-type), *AB (AB wild- type), Tübingen and Tup Longfin. Non pigmented embryos were generated by raising in 0.2 mM phenylthioarbam ide at 28.5 °C. After dechorination, and throughout manipulations, embryos were maintained in embryo medium containing 10 mM Hepes (from IM stock of Hepes in dHjO pH7.2) (hence forward referred to as buffered EM).
• Observation of live embryos
C hapter 2 G eneral M aterials and M ethods
• Wounding and culturing of epiboly stage embryos
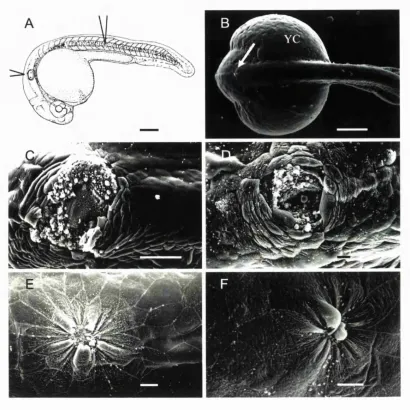
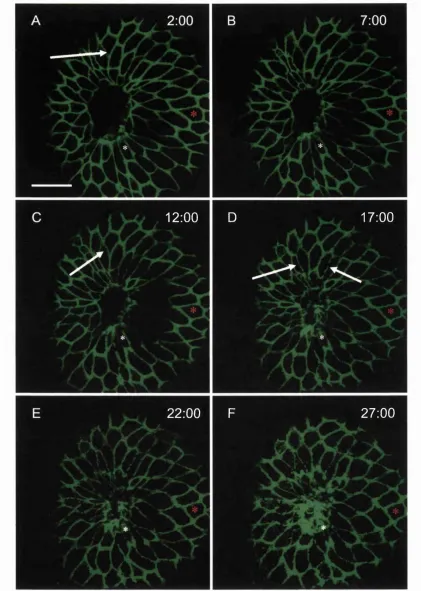
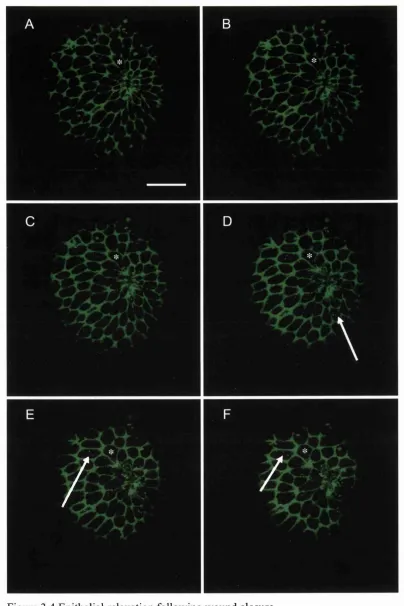
Embryos were dechorinated in petri dishes lined with 1 % Agarose in EM. For wounding, transplantation pipettes were pulled from 1 mm borosilicate glass capillaries (World Precision Instruments, IB 100-4) and broken off using watchmaker's forceps to an inner diameter of approximately 100 pm. The pipettes were loaded onto an oil-filled pipette holder (World Precision Instruments, 5430-10) and held by a 3-axis micromanipulator (Narishige, MN-153) connected by a continuous stream of mineral oil to a 50 pi Hamilton syringe driven by a micrometer-controlled syringe pump (Stoelting, 51218). Embryos at approximately 50% epiboly were transferred to wounding wells, preformed using an acrylic mould (made by Anatomy, UCL workshops) in 1% agarose in EM. The mould left wells approximately 1.0 mm deep by 1.0 mm wide, with the bottom surface of the well sloping slightly down from the back of the well. Embryos were placed into these wells and orientated with the animal pole uppermost. To make the wound, tissue from the animal pole was gently drawn in and out of the pipette about 3 or 4 times until the tissue became removed from the surrounding tissue and yolk. This leaves a circular wound approximately 100 pm in diameter (Fig 3.2 A, C). After wounding, embryos were left in their wells and allowed to heal in buffered EM for defined timepoints at 28.5°C.
• Wounding and culturing of 24 hour and later embryos
C hapter 2 G eneral M aterials an d M ethods
watchmaker's forceps into buffered EM where they were allowed to heal in petri-dishes for defined timepoints at 28.5°C.
• Processing embryos for scanning electron microscopy (SEM)
Embryos were fixed overnight in freshly prepared, ice-cold 1/2 strength Kamovsky's fixative (Kamovsky, 1965) at 4°C. After fixation the embryos were washed in O.IM sodium cacodylate buffer for 20 minutes, post-fixed in a solution of 1% osmium tetroxide / 0.1 M sodium cacodylate at 4°C for 20 minutes, and rinsed twice in 0.1 M sodium cacodylate for 10 minutes each. Specimens were then dehydrated through a graded series of alcohols - 30%, 50%, 70%, 90%, 95% and 100% - 5-10 minutes in each with two final rinses in 100% ethanol. Embryos were then washed in 50:50 100% ethanol:acetone for 10 minutes, and then washed twice in 100% acetone for 10 minutes before being critical-point dried in CO2. Once dry, embryos were mounted on carbon adhesive discs on SEM stubs
(Agar Scientific) and coated with approximately 30nm of gold and finally viewed using a JEOL 5410LV.
• Processing embryos for light histology and transmission electron microscopy
(TEM)
C hapter 2 G eneral M aterials an d M ethods
in individual plastic moulds and Araldite was added to fill the capsule. The moulds were cured overnight at 60°C.
The plastic moulds containing embedded embryos were cut away using a razor blade leaving an Araldite stub containing the embryos in random orientation. Thick, 10 p,m, sections were cut on a microtome (Reichert, OmU3), dried onto Tespa-coated slides, and stained with Toluidine Blue for 2 minutes at 65°C. For TEM, ultra-thin sections (60-70 nm) were cut on an ultra-microtome (Reichert ultra-cut E) and floated off into distilled water, prior to mounting on copper grids. Sections were stained with lead citrate (BDH) and viewed on a Jeol Transmission Electron Microscope (TEM)-1010.
• Preparing embryos for phalloidin staining
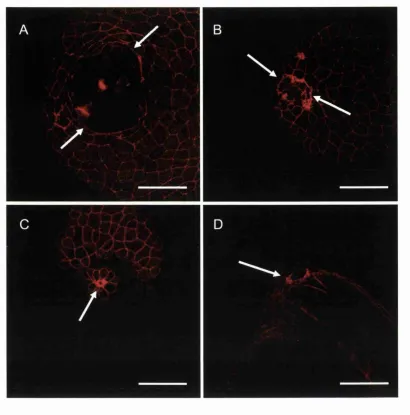
Embryos were fixed overnight in 4% paraformaldehyde at 4°C, then rinsed 3 times for 10 minutes in PBS, permeabilised for 30 minutes at room temperature on a shaker in PBS containing 0.5% Triton X-100 (PBT), washed in PBS, and then stained in PBT containing 250ng/ml FITC-phalloidin or Alexa 566 phalloidin (Sigma) for several hours at 4°C with gentle agitation. Embryos were then rinsed three times in PBT for 10 minutes each and mounted in Citifluor (UKC, Kent, U.K.) under a coverslip and filamentous actin was imaged using a Leica confocal laser scanning microscope (Leica SP).
• Live staining of embryos with fluorescent vital dyes
C hapter 2 G eneral M aterials and M ethods
distilled H2O). The lecEmbryos were then rinsed twice in buffered EM for 5 minutes and
mounted in 1 % low melting point agarose in buffered EM, in a purpose-built depression slide made by sticking a glass ring onto a microscope slide using silicone grease (Dow Coming). Buffered EM was added to the well to cover the embryo and the embryos were viewed using 20x and 40x water-immersion lenses on a Leica confocal laser scanning microscope (Leica SP).
• Preparing embryos for immunostaining of microtubules
Embryos were transferred to glass vials containing Microtubule fixative (see Appendix No 1) for 4-5 hours at room temperature. The specimens were then transferred to 100% methanol and stored in fresh methanol overnight at 4°C. The following day, embryos were rehydrated to PBS through 3:1, 1:1,1:3 methanohPBS and followed by several PBS washes throughout the day. Embryos were incubated overnight in 1:500 dilution of mouse monoclonal anti-a-tubulin (Sigma T-5168) in PBS. Embryos were washed in PBS for the following day and then incubated in a 1:200 dilution of Fluorescein-conjugated goat anti mouse IgG (H+L) (Pierce No. 31544). The following day embryos were washed several times in PBS and mounted in Citifluor under a glass coverslip and viewed using 20x, 40x and 63x oil-immersion lenses on a Leica confocal laser scanning microscope.
• Treatment of embryos with specific inhibitors
C hapter 2 G eneral M aterials and M ethods
Japan), embryos were dechorinated two hours prior to wounding, at approximately sphere - dome stage (4 hours post fertilisation), and incubated at 28.5°C in 750 |xM Y27632 in buffered EM (stock solution of lOmM in DMSO). Embryos had one of two fates. Embryos were either wounded and allowed to heal for 30 minutes, whilst still bathing in Y 27632, before being removed from the medium and processed for scanning electron microscopy. In other cases, immediately following wounding, embryos were stained for 5 minutes in lectin and subsequently time-lapse imaged during wound closure using the CLSM (Leica SP) as described above.
For treatment with the microtubule depolymerising blocker nocodazole, embryos were dechorinated approximately one hour prior to wounding (40% epiboly), and incubated in 5 p,g/ml nocodazole in EM (stock solution 2mg/ml in DMSO) (Sigma M-1404). Some embryos from each experimental batch were fixed after one hour and processed for anti-a- tubulin immunocytochemistry to confirm disruption of microtubules after one hour. The remaining embryos were wounded in the presence of nocodazole, and either allowed to heal for 10 minutes and processed for anti-a-tubulin to study microtubule orientation, or allowed to heal for 30 minutes, and processed for SEM to analyse wound closure.
• Imaging software for fixed tissue and time-lapse movies
C hapter 2 G eneral M aterials and M ethods
averaged twice and samples were collected every 30 or 60 seconds. The time-lapse series were assembled and analysed using NIH image software 1.6 (http://rsb.info.nih.gov/nih- image/). Optical sections corresponding to the same time point were projected into a single image and the background noise was reduced using a median filter. For fixed tissues, images were compiled from up to 15 optical sections accumulated up to 4 times and saved as a single projection using NIH image software. The final images were optimised and assembled into a plate using Adobe Photoshop 5.5.
• Testing the role of Calcium during wound closure
Shield-stage embryos were incubated in either 0.3 x or 1 x Danieau solution (Ix Danieau: 58 mM NaCl, 0.7 mM KCl, 0.4 mM MgSO^, 0.6 mM Csl(NO^)2, 5 mM Hepes, pH 7.6),
(Shih and Fraser, 1995), and wounded as described above. Embryos were then allowed to heal in 0.3 or 1 x Danieau's and fixed after either 0, 10 or 20 minutes and assessed for wound closure by SEM.
• Immunostaining for anti-phospho-histone-3
C hapter 2 G eneral M aterials an d Methods
reacted against Fluorescein conjugated goat anti-rabbit (Pierce No. 31572) at 1:200 in PBS at room temperature for 4 hours. Specimens were washed in PBS and incubated with 250ng/ml Alexa 566 phalloidin (Sigma) for 4 hours at room temeperature before washing overnight in PBS and mounting in Citiflor for viewing on the CLSM the following day.
• Whole-mount immunocytochemistry for anti-active ERK on Zebrafish embryos
Embryos were fixed in 10% formalin in PBS for 2 hours whilst shaking gently. Embryos were then washed for 10 mins in PBS, and then treated with 10 )xg/ml proteinase K in PBS for 5 mins, washed for 10 minutes in PBS and postfixed in 10% formalin in PBS for 20 minutes, and then washed again in PBS for 10 minutes. Embryos were then washed for 2 x 1 hour in BBT and blocked for one hour in 5% horse serum in BBT. Embryos were then incubated overnight at 4°C in monoclonal anti-activated-MAP kinase (Sigma M-8159) at 1:1000 dilution in BBT with 5% horse serum. The following days the embryos were washed all day in BBT with a final block with 5% horse serum. Embryos were incubated overnight in peroxidase-conjugated goat anti-mouse 2° antibody (Stratatech Sci/Jackson Labs 115-035-044) at 1:100 in BBT with 5% goat serum. The following day embryos were washed in BBT for one hour and several 30 minute washes in PBT before DAB reaction.
• Whole-mount immunocytochemistry for anti-active p38 and anti-active JNK on
C hapter 2 G eneral M aterials an d M ethods
Embryos were fixed overnight in 4% paraformaldehyde in PBS at 4°C. The following day embryos were washed several times in PBS, before being transferred to PBT to continue washing. Embryos were blocked for 1 hour in 5% goat serum in PBT before being incubated with primary antibody overnight at 4°C. For detection of active p38, NEB Phospho-p38 MAP Kinase (Thrl80/Tyrl82) polyclonal Rabbit Antibody was used at 1:10 dilution in PBT (NEB 921 IS). For detection of active-JNK, Promega Anti-ACTIVE JNK pAb Rabbit polyclonal was used (Promega V7931) at 1:100 dilution in PBT.
The following day embryos were washed all day in PBST and then reacted against Fluorescein conjugated goat anti-rabbit (Pierce No. 31572) at 1:200 in PBS at 4°C overnight. The following day embryos were washed several times in PBS before mounting in Citiflor and viewed on a CLSM.
• Treatment with MAP kinase inhibitors
1 mg of MEK Inhibitor U0126 (Promega V I 121) was resuspended in 234 pi of DMSO to produce a stock solution of 10 mM. Embryos were incubated in a working dilution of 100 pm in buffered EM for one hour prior to wounding and throughout the procedure.
C h a p te r s Introduction
CHAPTER THREE
Cvtoskeletal changes during wound healing in the
Zebrafish Brachvdanio Rerio
Introduction
• Embryonic epithelial wound front cells assemble an actin cable
As discussed in my Introductory chapter, embryonic wounds close by a combination of re- epithelialisation and contraction of the underlying mesenchymal wound bed. Re- epithelialisation is driven by an actin purse-string which forms in the leading edge of basal epidermal wound edge cells (reviewed in Woolley and Martin, 2000). This wound-induced purse-string was first observed in the chick embryo (Martin and Lewis, 1992) and in the mid-gestational mouse embryo (McCluskey and Martin, 1995), and most recently in
C h a p te rs Introduction
In the chick embryo, an actin cable begins to assemble in leading edge basal epithelial cells within minutes of wounding and, although discontinous within the first minutes, has become continuous by 5 minutes. The cable reaches its full thickness after 30 minutes and it persists until the wound is closed. After the advancing epithelial edges have made contact, the cable disassembles, transiently leaving disorganised actin in the cells which it has just zippered closed (Brock et al., 1996).
When cytochalasin D, which blocks actin polymerisation, is administered to the wound, assembly of the cable fails and wounds completely fail to epithelialise, further supporting the model that the actin cable is responsible for driving embryonic wound repair (McCluskey and Martin, 1995).
In the chick embryo, immunohistochemistry has revealed that large clusters of cadherins, the transmembrane adhesion components of adherens junctions, localise to the leading edge of basal wound edge cells co-incident with actin cable formation and presumably reflect where the cable is linked from cell to cell by adherens junctions. These junctions are seen within 5 minutes of wounding, and persist throughout wound closure.
• Actin cables are also observed in tissue culture and repair of some adult tissues
C h a p te rs Introduction
embryonic stage of development, coincident with epithelial stratification of the developing skin (J.Brock, thesis London 1997). In addition to embryonic wounds, some epithelial cell- lines also use contractile purse-string machinery to close a wound. Caco-Z^^^ cells, an intestinal epithelial cell line, are able to repair small wounds in confluent monolayers using a multicellular actinomyosin purse-string continuous from cell to cell, just as seen in embryonic skin wounds (Bement et al., 1993). However, in wounds that are larger than ten or so cell diameters across, they resort to lamellipodial crawling to repair, in a way more like adult skin healing. In the smaller wounds, several cytoskeletal proteins were found to co-localise with the actin filaments. Myosin-II was found in a similar pattern to actin, at the edge of the wound, tightly associated with the actin cable, along with the actin filament binding protein, tropomyosin (Bement et al., 1993).
C h a p te r s Introduction
Strong cellx ell junctions instead utilise adhesions to their underlying matrix substrate to
drag themselves forward.
• Formation of the purse-string is dependent on the small GTPase Rho
Multicellular purse-string assembly in the wounded embryonic epithelium is dependent on firing of small GTPases. Studies in 3T3 fibroblasts have demonstrated that the small GTPase Rho mediates stress fibre assembly in response to extracellular cues (Ridley and Hall, 1992), whilst Cdc42 and Rac, mediate assembly of filopodia and lamellae respectively (Nobes and Hall, 1995; Ridley et al., 1992). Since the wound actin-cable resembles oriented, bundled stress-fibres it seemed likely that Rho might be the controlling switch. Indeed loading of wound edge cells with the Rho blocker, C3 transferase, does prevent actin-cable assembly and subsequent re-epithelialisation of the wound, whilst blocking Rac from firing does not inhibit re-epithelialisation (Brock et al., 1996).
Rho has also been shown to be important during in vitro wound healing. A selection of Rho-inhibitors, including C3 transferase, toxins A and B, and dominant negative form of Rho A, prevented wound repair in two epithelial cell lines, (IEC-6 cells and Caco-2 cells), by inhibiting stress fibre formation and subsequent cell migration (Santos et al., 1997).
C h a p te r s Introduction
just as initial clues as to the action of Rho came from tissue culture, similarly we can guess from the downstream events in tissue culture cells, which pathways are most likely to be important during wound healing. One of Rho’s downstream targets is Rho-kinase. Rho- kinase family members include ROCK-I (also known as ROKp and p i 60^°^^) and ROCK- II (also known as RO K -a or Rho-kinase). ROCK-I and ROCK-II induce stress fibre assembly and assembly of focal contacts, but are not involved in other downstream functions of Rho, such as SRF-regulated transcription, cell transform ation or phosphoinositide metabolism. During cytokinesis, actinomyosin contraction is thought to be controlled by another Rho-effector, citron kinase, which co-localises with Rho at the cleavage furrow in Swiss3T3 cells (Madaule et al., 1998). However a dominant-negative ROCK construct inhibits cytokinesis when injected into Xenopus embryos and mammalian cells implicating a role for Rho-kinase too (Yasui et al., 1998). Therefore, Rho-kinase is an ideal candidate for mediating the signal as it directs actin purse-string assembly at the wound site.
C h a p te r s Introduction
proteins may also be targets and are also known to be involved in stress fibre assembly (reviewed in Aspenstrom, 1999). To test the role of Rho-kinase during wound healing, an inhibitor to this protein is now available. This inhibitor, Y-27632, is a high-affinity inhibitor of Rho-kinase and inhibits the formation of stress fibres in HeLa cells, but not any of the other activities of RhoA, such as gene transcription (Uehata et al., 1997).
• Actin purse-strings are found in single cell wounds
C h a p te rs Introduction
as a nucléation site to which more actin and myosin could be recruited. Single cells, in contrast, have to assemble the purse-string de novo. The single cell oocyte purse-string, like a multicellular embryonic wound, is also dependent on Rho activation and treatment of wounds with C3 transferase prevents assembly and subsequent wound closure (Bement et al., 1999).
• Assembly of the actin cable in X en o p u s oocytes correlates with a microtubule
network
Oocyte wounding also leads to a dramatic reorganisation of the microtubule cytoskeleton - a dense microtubule network rapidly localises to the region of cortical cytoplasm adjacent to the wound site (Bement et al., 1999). Inhibition of microtubule polymerisation at the time of wounding results in a failure of actinomyosin purse-string formation and blocks cytoplasmic repair, suggesting that microtubules are in some way required for delivery of at least some of the components of the actinomyosin purse-string to the wound site. However, as soon as the cable is assembled, microtubules seem no longer to be required, and may even be a hindrance to repair, since microtubule inhibitors, delivered minutes after wounding, lead to faster repair of the cortex than in untreated oocytes.
C h a p te r s Introduction
C h a p te r s Results
Results
I wounded embryos at two different developmental timepoints during Zebrafish embryogenesis, allowing me to address two different sets of questions. Wounds made to the epithelial sheet overlying a 50% epiboly embryo allow me to directly observe assembly of the cytoskeletal machinery that drives re-epithelialisation and to analyse the cell shape changes and shufflings that contribute to closure of the epithelial hole. Also, importantly, this early stage gives an opportunity to test the genetics of epithelial repair by wounding mutants that are defective in the cell movements of morphogenetic movements like gastrulation and epiboly. In addition to the early stages of development I also wounded the 24 hour embryo, which has well developed skin with mature epidermis and underlying mesenchyme, and which is more similar to the developmental stages of chick and mouse embryo wound healing studies previously performed in this lab.
• Wounds made to the 24 hour fish heal rapidly by epithelial cell shape changes.
Using a tungsten needle I made stab wounds to either the tail of the 24 hour zebrafish (approximately one third of the way between the yolk extension and the tail tip), or to the dorsal side of the embryo between the otic vesicles (see Fig 3.1 A, B)