Scents and sex: insect pheromones
Full text
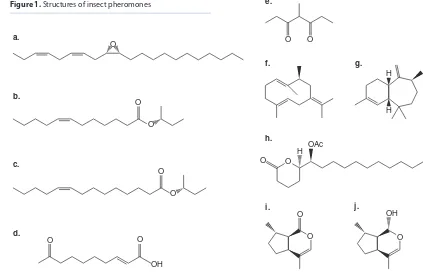
Figure


Related documents
The event dummy variables (Extended Dummy Variable, One-Month Dummy) have a greater impact on meat shares than do the word-count variables, perhaps because the dummy
On the contrary, after the scleral spurs are marked by the clinician, the internal software of SL-OCT automatically calculates the anterior and posterior corneal
Dotknutá osoba má právo na základe písomnej žiadosti vyžadovať od prevádzkovateľa najmä potvrdenie, či sú alebo nie sú jej osobné údaje spracúvané, presné informácie
Upon comparing those with parents with chronic illness to those without, we found that emerging adults with a parent with chronic illness have significantly higher levels of anxiety,
Students are informed to collect their admit cards from examination department after account clearance. No student will be admitted to the examination hall
In this way, the objectives of this research are to predict the future of Spain’s public broadcasting media system according to the current political context, and to determine
to explicit shape or assumed size dummies That is in the advanced slides on procedures Sorry – but it ’ s complicated to explain..
