The Na
+
-plus-K
+
-dependent adenosine triphosphatase
(Na
+
+K
+
-ATPase) is a transmembrane protein composed of
three subunits (
α
,
β
,
γ
) responsible for pumping three Na
+
out
of animal cells in exchange for two K
+
or NH
4
+
per ATP
hydrolyzed. Situated in the basolateral membrane of most
epithelial cells, the Na
+
+K
+
-ATPase not only mediates
electrogenic transfer of Na
+
from the cytosol to the
extracellular fluid and K
+
or NH
4
+
from the extracellular fluid
to the cytosol but also establishes electrochemical gradients
used by apical and basolateral transporters such as the Na
+
/H
+
exchanger, Na
+
/K
+
/2Cl
−
cotransporter and various ion
channels. Originally described enzymatically in homogenates
of nerves from the shore crab Carcinus maenas (Skou, 1957),
the Na
+
+K
+
-ATPase
α
- and
β
-subunits were first cloned and
sequenced from mammalian kidney (Shull et al., 1985, 1986).
The
α
-subunit provides the catalytic function of the Na
+
+K
+
-ATPase, binding and hydrolyzing ATP and itself becoming
phosphorylated during the transport cycle. The
α
-subunit also
binds ouabain, a highly specific inhibitor. The
β
-subunit is
thought to participate in anchoring the complex in the
basolateral membrane and the
γ
-subunit, not always present,
appears to serve a regulatory role (for a review, see Therien
and Blostein, 2000).
Many studies have shown that the enzymatic activity of the
Na
+
+K
+
-ATPase in homogenates of ion-transporting gill tissue
is enhanced when euryhaline crustaceans are subjected to
osmoregulatory stress following transfer from sea water to
dilute salinities (for reviews, see Towle, 1990; Péqueux,
1995). For example, the specific Na
+
+K
+
-ATPase activity in
homogenates or microsomal fractions of posterior gills of
Printed in Great Britain © The Company of Biologists Limited 2001
JEB3727
Many studies have shown that hyperosmoregulation in
euryhaline crabs is accompanied by enhanced Na
+
+K
+
-ATPase activity in the posterior gills, but it remains
unclear whether the response is due to regulation of
pre-existing enzyme or to increased gene transcription and
mRNA translation. To address this question, the complete
open reading frame and 3
′
and 5
′
untranslated regions of
the mRNA coding for the
α
-subunit of Na
+
+K
+
-ATPase
from the blue crab Callinectes sapidus were amplified
by reverse transcriptase/polymerase chain reaction
(RT-PCR) and sequenced. The resulting 3828-nucleotide cDNA
encodes a putative 1039-amino-acid protein with a
predicted molecular mass of 115.6 kDa. Hydrophobicity
analysis of the amino acid sequence indicated eight
membrane-spanning regions, in agreement with
previously suggested topologies. The
α
-subunit amino acid
sequence is highly conserved among species, with the blue
crab sequence showing 81–83 % identity to those of other
arthropods and 74–77 % identity to those of vertebrate
species. Quantitative RT-PCR analysis showed high levels
of
α
-subunit mRNA in posterior gills 6–8 compared with
anterior gills 3–5. Western blots of gill plasma membranes
revealed a single Na
+
+K
+
-ATPase
α
-subunit protein band
of the expected size. The posterior gills contained a much
higher level of
α
-subunit protein than the anterior gills, in
agreement with previous measurements of enzyme
activity. Immunocytochemical analysis showed that the
Na
+
+K
+
-ATPase
α
-subunit protein detected by
α
5
antibody is localized to the basolateral membrane region
of gill epithelial cells. Transfer of blue crabs from 35 to
5 ‰ salinity was not accompanied by notable differences
in the relative proportions of
α
-subunit mRNA and
protein in the posterior gills, suggesting that the enhanced
Na
+
+K
+
-ATPase enzyme activity that accompanies the
hyperosmoregulatory response may result from
post-translational regulatory processes.
Key words: Na
+
+K
+
-ATPase, gills, blue crab, Callinectes sapidus,
cDNA sequencing, salinity, osmoregulation, quantitative polymerase
chain reaction.
Summary
Introduction
Na
+
+K
+
-ATPase in gills of the blue crab Callinectes sapidus: cDNA sequencing
and salinity-related expression of
α
-subunit mRNA and protein
David W. Towle
1,
*, Ryan S. Paulsen
1
, Dirk Weihrauch
1
, Marek Kordylewski
1
, Cristina Salvador
2
,
Jean-Hervé Lignot
3
and Céline Spanings-Pierrot
3
1
Mount Desert Island Biological Laboratory, Salsbury Cove, ME 04672, USA and Lake Forest College, Lake Forest,
IL 60045, USA,
2
Duke University, Durham, NC 27706, USA and
3
Université Montpellier II, 34095 Montpellier
Cedex 5, France
*e-mail: [email protected]
1 GTTGTCGTGAGGCGCGCCTCGAGGAGAGCAGACGTGCGCCCTGCCCAGCAGTGTCGACCACCAAACACTCCACACGCTGAAGAATGTGCTTGTAGCGAAAACCGTCACCAGTCCAAGAGG 120
121 GCAACCAGAGCGAGTGAACGAGTGTACAGAGTGAACGTAGCCTAGTGGCGTGTACATAAGTGTGTACATAAGAGTGATCGGTGGTGCTGGTGGAAGCCAGCGTCTGGCCGTTGTTCGTGG 240
241 ATACCGTGTATCCTCTGCTGCCGTCCTTCTGCTGAGCAGGGCCACCCGACGCCCGCTACTACTCCAGCAACC ATG GCTGACACGGGCAGAACAGATTCGTACCGCCACGCTACGGATCGT 360 1 M A D T G R T D S Y R H A T D R 16
361 AACATCCCTGATGATAACCGAACTGTGAAGGGAGACCCCAAGTCAAAAAAGAAGAATGTCAAGGGGAAGAGGAAAGGGGAGAAGGAGAAGGATATGGACAACCTGAAACAGGAATTGGAA 480 17 N I P D D N R T V K G D P K S K K K N V K G K R K G E K E K D M D N L K Q E L E 56
481 CTCGATGAACACAAGGTCCCCATAGAAGAACTCTTTCAGCGCCTCTCCGTCAATCCTGACACAGGTTTGACACAAGCCGAGGCCCGTCGCAGGTTAGAAAGGGATGGCCCCAATGCCCTC 600 57 L D E H K V P I E E L F Q R L S V N P D T G L T Q A E A R R R L E R D G P N A L 96
601 ACCCCTCCCAAGCAAACTCCAGAATGGGTCAAATTTTGCAAGAACCTGTTTGGTGGATTTTCACTGCTGCTGTGGATTGGTGCCATTCTCTGTTTCATTGCTTACTCCATTGAAGCTGCA 720 97 T P P K Q T P E W V K F C K N L F G G F S L L L W I G A I L C F I A Y S I E A A 136
721 TCAGAGGAGGAACCCAACAATGATAACTTGTACTTGGGCATTGTGCTCACTGCTGTCGTAATCATCACAGGCATCTTCTCATACTACCAGGAGAGCAAGAGTTCCCGTATCATGGAGTCC 840 137 S E E E P N N D N L Y L G I V L T A V V I I T G I F S Y Y Q E S K S S R I M E S 176
841 TTCAAAAACCTTGTTCCTCAATATGCTATTGTCATCCGTGAAGGTGAGAAACTGAATGTGCAGGCTGAAGAACTATGCATAGGTGACATCATTGATGTTAAGTTTGGTGATCGTATCCCT 960 177 F K N L V P Q Y A I V I R E G E K L N V Q A E E L C I G D I I D V K F G D R I P 216
961 GCTGACATGCGAGTCATCGAAGCTCGAGGTTTCAAGGTCGACAACTCTTCACTCACTGGTGAGTCTGAGCCACAGAGTCGCTCTCCTGAATTCACATCCGAGAATCCTCTTGAGACCAAG 1080 217 A D M R V I E A R G F K V D N S S L T G E S E P Q S R S P E F T S E N P L E T K 256
1081 AACCTTGCCTTCTTTTCCACCAATGCTGTTGAAGGTACTTGCAAGGGTATTGTAATCAATATTGGTGACAACACTGTGATGGGTCGCATTGCTGGTCTGGCCTCTGGCTTGGAAACTGGA 1200 257 N L A F F S T N A V E G T C K G I V I N I G D N T V M G R I A G L A S G L E T G 296
1201 GAGACTCCAATTGCCAAGGAGATCAGTCACTTCATCCACATCATTACCGGTGTGGCCGTCTTCCTGGGTGTTACATTCTTCGTCATTGCTTTCATCATGGGTTACCATTGGTTGGATGCT 1320 297 E T P I A K E I S H F I H I I T G V A V F L G V T F F V I A F I M G Y H W L D A 336
1321 GTTGTGTTCCTCATTGGTATCATTGTGGCCAATGTGCCCGAGGGTCTGCTGGCCACTGTCACAGTGTGTCTCACTCTCACTGCCAAGCGTATGGCTGCAAAGAACTGCTTGGTGAAGAAC 1440 337 V V F L I G I I V A N V P E G L L A T V T V C L T L T A K R M A A K N C L V K N 376 >>>>>>>>>>>>>>>>>>>>>>>>>>>>>>>
1441 TTGGAGGCTGTGGAAACCCTGGGTTCAACCTCTACCATCTGCTCTGACAAGACTGGTACCCTCACTCAGAACCGTATGACAGTCGCCCACATGTGGTTTGACAACACCATCATCGAGGCA 1560 377 L E A V E T L G S T S T I C S D K T G T L T Q N R M T V A H M W F D N T I I E A 416
1561 GATACCTCAGAGGACCAGTCAGGCTGCCAGTACGACAAGACTTCCGATGGATGGAAGGCTCTCTCCCGGATTGCTGCTCTCTGCAACCGTGCTGAATTCAAGACAGGTCAGGAAGATGTA 1680 417 D T S E D Q S G C Q Y D K T S D G W K A L S R I A A L C N R A E F K T G Q E D V 456
1681 CCCATTCTGAAGCGAGAGGTGAATGGTGATGCTTCTGAGGCTGCTCTGCTGAAGTGTGTGGAACTTGCTATTGGAGACGTGAGGGGCTGGCGTTCCCGCAATAAGAAGGTTTGTGAGATT 1800 457 P I L K R E V N G D A S E A A L L K C V E L A I G D V R G W R S R N K K V C E I 496
1801 CCTTTCAACTCCACCAACAAGTACCAAGTGTCTATCCACGAGACCCAGGACAAGAATGACCTCCGCTACCTGCTTGTGATGAAGGGTGCTCCAGAGAGGATCCTTGAACGATGCTCCACC 1920 497 P F N S T N K Y Q V S I H E T Q D K N D L R Y L L V M K G A P E R I L E R C S T 536
1921 ATCTTCATGAATGGTGAGGAGAAGCCTTTGGATGAGGAGATGAAGGAATCTTTCAACAATGCTTACTTGGAATTGGGTGGGCTGGGAGAGCGTGTGCTGGGCTTCTGTGACTATGTTCTG 2040 537 I F M N G E E K P L D E E M K E S F N N A Y L E L G G L G E R V L G F C D Y V L 576
2041 CCCTCCGACAAGTACCCCCTCGGCTATCCTTTCGATGCTGATGCTGTCAACTTCCCCGTGCACGGTCTTCGATTCGTCGGCCTCATGTCCATGATTGATCCTCCCCGTGCTGCTGTGCCC 2160 577 P S D K Y P L G Y P F D A D A V N F P V H G L R F V G L M S M I D P P R A A V P 616 <<<<<<<<<<<<<<<<<<<<<<<<<<<<<
2161 GACGCCGTGGCCAAGTGCCGTTCTGCTGGTATCAAGGTCATCATGGTTACTGGTGATCACCCCATCACTGCCAAGGCCATTGCCAAGTCTGTGGGTATCATTTCTGAGGGCAATGAGACT 2280 617 D A V A K C R S A G I K V I M V T G D H P I T A K A I A K S V G I I S E G N E T 656
2281 GTGGAGGACATTGCCCAGAGACTCAACATTCCCATCAAGGAGGTCGACCCCCGCGAGGCCAAGGCTGCTGTGGTCCACGGCTCTGAGTTGAGGGACATGACTTCTGAACAGCTGGATGAT 2400 657 V E D I A Q R L N I P I K E V D P R E A K A A V V H G S E L R D M T S E Q L D D 696
2401 GTCCTTATCCACCACACTGAGATTGTGTTTGCCCGTACCTCCCCACAGCAGAAGCTCATCATTGTGGAGGGCTGCCAGCGTATGGGAGCCATTGTAGCTGTAACTGGAGATGGTGTAAAT 2520 697 V L I H H T E I V F A R T S P Q Q K L I I V E G C Q R M G A I V A V T G D G V N 736
2521 GACTCCCCTGCCCTCAAGAAGGCCGATATTGGTGTGGCCATGGGTATTGCTGGATCAGACGTGTCTAAGCAGGCTGCCGACATGATTCTGTTGGATGACAACTTTGCTTCTATTGTCACT 2640 737 D S P A L K K A D I G V A M G I A G S D V S K Q A A D M I L L D D N F A S I V T 776
2641 GGTGTGGAAGAGGGCAGGCTTATTTTTGATAACCTGAAGAAATCTATTGCCTACACCCTCACTTCAAACATCCCTGAGATCTCCCCCTTCTTGTTCTTCATGATTGCCTCCGTGCCTCTA 2760 777 G V E E G R L I F D N L K K S I A Y T L T S N I P E I S P F L F F M I A S V P L 816
2761 CCTCTGGGCACTGTTACCATTCTCTGCATTGATCTGGGTACTGACATGGTGCCTGCCATTTCCCTTGCCTATGAAGAAGCTGAGTCAGATATTATGAAGCGCCAGCCCCGCAATCCCTTC 2880 817 P L G T V T I L C I D L G T D M V P A I S L A Y E E A E S D I M K R Q P R N P F 856
2881 ACGGACAAGCTTGTGAACGAGAGGCTCATCTCCATGGCCTATGGTCAGATTGGCATGATCCAAGCTCTGGCTGGATTCTATGTGTACTTTGTCATCATGGCTGAGAACGGGTTCCTGCCT 3000 857 T D K L V N E R L I S M A Y G Q I G M I Q A L A G F Y V Y F V I M A E N G F L P 896
3001 CCCGTCCTCTTTGGTATCCGTGAGCAGTGGGACTCCAAGGCCATCAACGATCTGGAGGATTACTATGGCCAGGAATGGACATACCATGACCGTAAGATCCTTGAGTACACCTGCCACACT 3120 897 P V L F G I R E Q W D S K A I N D L E D Y Y G Q E W T Y H D R K I L E Y T C H T 936
3121 GCATTCTTTGTGGCCATTGTGGTGGTGCAGTGGGCTGACTTGATCATCTGTAAGACTCGCCGTAACTCCATCCTTCACCAGGGCATGAAGAACATGGTGCTTAACTTTGGATTGTGTTTC 3240 937 A F F V A I V V V Q W A D L I I C K T R R N S I L H Q G M K N M V L N F G L C F 976
3241 GAGACTACACTGGCTGCCTTCCTGTCCTACACACCAGGCATGGACAAGGGTCTAAGGATGTACCCCCTCAAATTCTACTGGTGGCTGCCCCCTCTCCCCTTCTCATTACTCATCTTCGTG 3360 977 E T T L A A F L S Y T P G M D K G L R M Y P L K F Y W W L P P L P F S L L I F V 1016
3361 TACGACGAGTGTCGCCGGTTCGTGCTGCGCAGGAACCCTGGTGGCTGGGTGGAGATGGAGACCTATTATTAAGGTTTGTAGTGAGCACAAGTTTACAAGAGCTCAAGAGCGTGGCCACCA 3480 1017 Y D E C R R F V L R R N P G G W V E M E T Y Y 1039
3481 CCAGCAGCCTCCTCCCTCACAGCCAGCACTTGCATCACTGTCACTCCTCTGTGATGTCTGGGGATTGGAACTGTTGTATTATAACCTTCAAAAGAAATGCTATTCTTTAATTAGAATCCA 3600
3601 GAGAATATATAACAACCAAGTGTTGGTGGCTCAGTAATGTGCTCTGTCAATAATGATTCCATTGCTTATTTCTATGTTACTTACTGGGGATTTTAGTGTCATTATGGTTGTTTCTAATTT 3720
[image:2.612.74.525.69.675.2]3721 CCCCATTGTTATATATAAGATTTGCAGTAAGTAACATTGAAATATTTGTACAGAATCCTGGTGAAAGCATGTATTAAAGAGAAAAAGTGACAAAAAAAAAAAAAAAAA 3828
Fig. 1. cDNA and predicted amino acid sequence of the Na
+
+K
+
-ATPase
α
-subunit from gills of the blue crab Callinectes sapidus. Stop codons
4007
Molecular physiology of crab gill Na
+K
-ATPase
* 2 0 * 4 0 * 6 0 * 8 0 * 1 0 0 C A L L I N E C T E S : M A D T G R T D S Y R H A T D R N I P D D N R T V K G D P K S K K K N V K G K R K G E . . . K E K D M D N L K Q E L E L D E H K V P I E E L F Q R L S V N P D T G L T Q A E A R R R L E R D G P N A L T : 9 7 D R O S O P H I L A : . . . M P A K V N . . . K K E N L D D L K Q E L D I D F H K I S P E E L Y Q R F Q T H P E N G L S H A K A K E N L E R D G P N A L T : 6 0 A R T E M I A 1 : . . . M A K G K Q K . . . K G K D L N E L K K E L D I D F H K I P I E E C Y Q R L G S N P E T G L T N A Q A R S N I E R D G P N C L T : 6 1 A R T E M I A 2 : . . . M G K K Q G . . . K Q L S D L K K E L E L D Q H K I P L E E L C R R L G T N T E T G L T S S Q A K S H L E K Y G P N A L T : 5 8 T O R P E D O : . . . M G K G A A S E K Y Q P A A T S E N A K N S K K S K S . . K T T D L D E L K K E V S L D D H K L N L D E L H Q K Y G T D L T Q G L T P A R A K E I L A R D G P N A L T : 8 1 A N G U I L L A : . . . M G R G T G H D Q Y E L A A T S E G G R K K K R D K . . . K K K D M D D L K K E V D L D D H K L T L D E L H R K Y G T D L T R G L T S S R A A E I L A R D G P N A L T : 8 0 X E N O P U S : . . . M G Y G A G R D K Y E P A A T S E Q G G K K K K G K G K G K E K D M D E L K K E V T M E D H K L S L D E L H R K F G T D M Q R G L T T A R A A E I L A R D G P N A L T : 8 3 G A L L U S : . . . M G D K G . . . E K E S P K K G K G . . . K R D L D D L K K E V A M T E H K M S I E E V C R K Y N T D C V Q G L T H S K A Q E I L A R D G P N A L T : 6 8 H O M O : . . . M G D K K D . . . D K D S P K K N K G K . . . E R R D L D D L K K E V A M T E H K M S V E E V C R K Y N T D C V Q G L T H S K A Q E I L A R D G P N A L T : 7 1 * 1 2 0 * 1 4 0 * 1 6 0 * 1 8 0 * 2 0 0 C A L L I N E C T E S : P P K Q T P E W V K F C K N L F G G F S L L L W I G A I L C F I A Y S I E A A S . E E E P N N D N L Y L G I V L T A V V I I T G I F S Y Y Q E S K S S R I M E S F K N L V P Q Y A I V I R E G E K L N V : 1 9 6 D R O S O P H I L A : P P K Q T P E W V K F C K N L F G G F A M L L W I G A I L C F V A Y S I Q A S T . S E E P A D D N L Y L G I V L S A V V I V T G I F S Y Y Q E S K S S K I M E S F K N M V P Q F A T V I R G G E K L T L : 1 5 9 A R T E M I A 1 : P P K T T P E W I K F C K N L F G G F A L L L W T G A I L C F L A Y G I E A S S G N E D M L K D N L Y L G I V L A T V V I V T G I F S Y Y Q E N K S S R I M D S F K N L V P Q Y A L A L R E G Q R V T L : 1 6 1 A R T E M I A 2 : P P R T T P E W I K F C K Q L F G G F Q M L L W I G S I L C F I A Y T M E K Y K . N P D V L G D N L Y L G L A L L F V V I M T G C F A Y Y Q D H N A S K I M D S F K N L M P Q F A F V I R D G K K I Q L : 1 5 7 T O R P E D O : P P P T T P E W I K F C R Q L F G G F S I L L W T G A I L C F L A Y G I Q V A T . V D N P A N D N L Y L G V V L S T V V I I T G C F S Y Y Q E A K S S K I M D S F K N M V P Q Q A L V I R D G E K S S I : 1 8 0 A N G U I L L A : P P P T T P E W V K F C R Q L F G G F S M L L W I G A I L C F L A Y G I Q A A S . E D E P A N D N L Y L G V V L S A V V I I T G C F S Y Y Q E A K S S R I M D S F K N L V P Q Q A L V I R D G E K K C I : 1 7 9 X E N O P U S : P P P T T P E W V K F C R Q L F G G F S M L L W I G A I L C F L A Y G I Q A A M . E E E P Q N D N L Y L G V V L S A V V I I T G C F S Y Y Q E A K S S K I M E S F K N M V P Q Q A L V I R S G E K L S I : 1 8 2 G A L L U S : P P P T T P E W V K F C R Q L F G G F S I L L W I G A I L C F L A Y G I Q A G T . E D E P S N D N L Y L G I V L A A V V I I T G C F S Y Y Q E A K S S K I M E S F K N M V P Q Q A L V I R E G E K M Q L : 1 6 7 H O M O : P P P T T P E W V K F C R Q L F G G F S I L L W I G A I L C F L A Y G I Q A G T . E D D P S G D N L Y L G I V L A A V V I I T G C F S Y Y Q E A K S S K I M E S F K N M V P Q Q A L V I R E G E K M Q V : 1 7 0 * 2 2 0 * 2 4 0 * 2 6 0 * 2 8 0 * 3 0 0 C A L L I N E C T E S : Q A E E L C I G D I I D V K F G D R I P A D M R V I E A R G F K V D N S S L T G E S E P Q S R S P E F T S E N P L E T K N L A F F S T N A V E G T C K G I V I N I G D N T V M G R I A G L A S G L E T G : 2 9 6 D R O S O P H I L A : R A E D L V L G D V V E V K F G D R I P A D I R I I E A R N F K V D N S S L T G E S E P Q S R G A E F T H E N P L E T K N L A F F S T N A V E G T A K G V V I S C G D H T V M G R I A G L A S G L D T G : 2 5 9 A R T E M I A 1 : K A E E L T M G D I V E V K F G D R V P A D L R V L E A R S F K V D N S S L T G E S E P Q A R S P E F T N D N P L E T K N L A F F S T N A V E G T M R G I V I G I G D N T V M G R I A G L A S G L D T G : 2 6 1 A R T E M I A 2 : K A E E V T V G D L V E V K F G D R I P A D I R I T S C Q S M K V D N S S L T G E S E P Q S R S T E C T N D N P L E T K N L A F F F T N T L E G T G R G I V I N V G D D S V M G R I A C L A S S L D S G : 2 5 7 T O R P E D O : N A E Q V V V G D L V E V K G G D R I P A D L R I I S A C S C K V D N S S L T G E S E P Q S R S P E Y S S E N P L E T K N I A F F S T N C V E G T A R G I V I N I G D H T V M G R I A T L A S G L E V G : 2 8 0 A N G U I L L A : N A E E V V A G D L V E V K G G D R I P A D L R V A S A Q G C K V D N S S L T G E S E P Q T R S P D F S N E N P L E T R N I A F F S T N C V E G T A R G V V I N T G D R T V M G R I A T L A S S L E V G : 2 7 9 X E N O P U S : N A E E V V L G D L V E V K G G D R I P A D L R V I S S H G C K V D N S S L T G E S E P Q T R S P D F T N E N P L E T R N I A F F S T N C V E G T A R G I V V N T G D R T V M G R I A T L A S G L D G G : 2 8 2 G A L L U S : N A E E V V V G D L V E V K G G D R V P A D L R I I S A H G C K V D N S S L T G E S E P Q T R S P D C T H D N P L E T R N I T F F S T N C V E G T A R G V V I A T G D R T V M G R I A T L A S G L E V G : 2 6 7 H O M O : N A E E V V V G D L V E I K G G D R V P A D L R I I S A H G C K V D N S S L T G E S E P Q T R S P D C T H D N P L E T R N I T F F S T N C V E G T A R G V V V A T G D R T V M G R I A T L A S G L E V G : 2 7 0 * 3 2 0 * 3 4 0 * 3 6 0 * 3 8 0 * 4 0 0 C A L L I N E C T E S : E T P I A K E I S H F I H I I T G V A V F L G V T F F V I A F I M G Y H W L D A V V F L I G I I V A N V P E G L L A T V T V C L T L T A K R M A A K N C L V K N L E A V E T L G S T S T I C S D K T G T : 3 9 6 D R O S O P H I L A : E T P I A K E I H H F I H L I T G V A V F L G V T F F V I A F I L G Y H W L D A V I F L I G I I V A N V P E G L L A T V T V C L T L T A K R M A S K N C L V K N L E A V E T L G S T S T I C S D K T G T : 3 5 9 A R T E M I A 1 : E T P I A K E I A H F I H I I T G V A V F L G V T F F I I A F V L G Y H W L D A V V F L I G I I V A N V P E G L L A T V T V C L T L T A K R M A S K N C L V K N L E A V E T L G S T S T I C S D K T G T : 3 6 1 A R T E M I A 2 : K T P I A R E I E H F I H I I T A M A V S L A A V F A V I S F L Y G Y T W L E A A I F M I G I I V A K V P E G L L A T V T V C L T L T A K R M A K K N C L V R N L E A V E T L G S T S T I C S D K T G T : 3 5 7 T O R P E D O : Q T P I A A E I E H F I H I I T G V A V F L G V S F F I L S L I L G Y T W L E A V I F L I G I I V A N V P E G L L A T V T V C L T L T A K R M A R K N C L V K N L E A V E T L G S T S T I C S D K T G T : 3 8 0 A N G U I L L A : R T P I S I E I E H F I H I I T G V A V F L G V S F F I L S L I L G Y A W L E A V I F L I G I I V A N V P E G L L A T V T V C L T L T A K R M A K K N C L V K N L E A V E T L G S T S T I C S D K T G T : 3 7 9 X E N O P U S : R T P I A I E I E H F I H I I T G V A V F L G V S F F I L S L I L Q Y T W L E A V I F L I G I I V A N V P E G L L A T V T V C L T L T A K R M A R K N C L V K N L E A V E T L G S T S T I C S D K T G T : 3 8 2 G A L L U S : K T P I A V E I E H F I Q L I T G V A V F L G I S F F V L S L I L G Y T W L E A V I F L I G I I V A N V P E G L L A T V T V C L T L T A K R M A R K N C L V K N L E A V E T L G S T S T I C S D K T G T : 3 6 7 H O M O : K T P I A I E I E H F I Q L I T G V A V F L G V S F F I L S L I L G Y T W L E A V I F L I G I I V A N V P E G L L A T V T V C L T V T A K R M A R K N C L V K N L E A V E T L G S T S T I C S D K T G T : 3 7 0 * 4 2 0 * 4 4 0 * 4 6 0 * 4 8 0 * 5 0 0 C A L L I N E C T E S : L T Q N R M T V A H M W F D N T I I E A D T S E D Q S G C Q Y D K T S D G W K A L S R I A A L C N R A E F K T G Q E D V P I L K R E V N G D A S E A A L L K C V E L A I G D V R G W R S R N K K V C E I : 4 9 6 D R O S O P H I L A : L T Q N R M T V A H M W F D N Q I I E A D T T E D Q S G V Q Y D R T S P G F K A L S R I A T L C N R A E F K G G Q D G V P I L K K E V S G D A S E A A L L K C M E L A L G D V M N I R K R N K K I A E V : 4 5 9 A R T E M I A 1 : L T Q N R M T V A H M W F D G T I T E A D T T E D Q S G A Q F D K S S A G W K A L V K I A A L C S R A E F K P N Q S T T P I L K R E V T G D A S E A A I L K C V E L T T G E T E A I R K R N K K I C E I : 4 6 1 A R T E M I A 2 : L T Q N R M T V A H M W F D Q K I V T A D T T E N Q S G N Q L Y R G S K G F P E L I R V A S L C S R A E F K T E H A H L P V L K R D V N G D A S E A A I L K F A E M S T G S V M N I R S K Q K K V S E I : 4 5 7 T O R P E D O : L T Q N R M T V A H M W F D N Q I H E A D T T E N Q S G I S F D K T S L S W N A L S R I A A L C N R A V F Q A G Q D S V P I L K R S V A G D A S E S A L L K C I E L C C G S V S Q M R D R N P K I V E I : 4 8 0 A N G U I L L A : L T Q N R M T V A H M W F D N Q I H E A D T T E N Q S G T S F D R S S A T W A A L A R I A G L C N R A V F L A E Q S N V P I L K R D V A G D A S E S A L L K C I E L C C G S V N D M R D K H V K I A E I : 4 7 9 X E N O P U S : L T Q N R M T V A H M W F D N Q I H E A D T T E N Q S G A S F D K S S P T W T A L S R V A G L C N R A V F Q A G Q E N T P I L K R D V A G D A S E S A L L K C I E L C C G S V R D M R E K N H K V A E I : 4 8 2 G A L L U S : L T Q N R M T V A H M W F D N Q I H E A D T T E D Q S G T S F D K S S A T W V A L S H I A G L C N R A V F K G G Q E N V P I L K R D V A G D A S E S A L L K C I E L S S G S V K V M R E R N K K V A E I : 4 6 7 H O M O : T Q N R M T V A H M W F D N Q I H E A D T T E D Q S G T S F D K S S H T W V A L S H I A G L C N R A V F K G G Q D N I P V L K R D V A G D A S E S A L L K C I E L S S G S V K L M R E R N K K V A E I : 4 7 0 * 5 2 0 * 5 4 0 * 5 6 0 * 5 8 0 * 6 0 0 C A L L I N E C T E S : P F N S N K Y Q V S I H E T Q D K N D L R Y L L V M K G A P E R I L E R C S T I F M N G E E K P L D E E M K E S F N N A Y L E L G G L G E R V L G F C D Y V L P S D K Y P L G Y P F D A D A V N F P V : 5 9 6 D R O S O P H I L A : P F N S T N K Y Q V S I H E T E D T N D P R Y L L V M K G A P E R I L E R C S T I F I N G K E K V L D E E M K E A F N N A Y M E L G G L G E R V L G F C D F M L P S D K Y P N G F K F N T D D I N F P I : 5 5 9 A R T E M I A 1 : P F N S A N K F Q V S I H E N E D K S D G R Y L L V M K G A P E R I L E R C S T I F M N G K E I D M T E E L K E A F N N A Y M E L G G L G E R V L G F C D Y L L P L D K Y P H G F A F N A D D A N F P L : 5 6 1 A R T E M I A 2 : P F N S A N K Y Q V S V H E R . . E D K S G Y F L V M K G A P E R I L E R C S T I L I D G T E I P L D N H M K E C F N N A Y M E L G G M G E R V L G F C D F E L P S D Q Y P R G Y V F D A D E P N F P I : 5 5 5 T O R P E D O : P F N S T N K Y Q L S I H E N . K A D S R Y L L V M K G A P E R I L D R C S T I L L N G E D K P L N E E M K E A F Q N A Y L E L G G L G E R V L G F C H L K L S T S K F P E G Y P F D V E E P N F P I : 5 7 9 A N G U I L L A : P F N S T N K Y Q L S I H K N A N S E E S K H L L V M K G A P E R I L D R C S T I M I H G K E Q P L D D E M K D A F Q N A Y V E L G G L G E R V L G F C H Y F L P D D Q F A E G F Q F D T E E V N F P T : 5 7 9 X E N O P U S : P F N S T N K Y Q L S V H K N A N P S E S R Y I L V M K G A P E R I L D R C T S I I L Q G K E Q P L D E E L K D A F Q D A Y L E L G G L G E R V L G F C H L A L P D D Q F P D G F Q F D T E E V N F P T : 5 8 2 G A L L U S : P F N S T N K Y Q L S I H E T E D P N D N R Y L L V M K G A P E R I L D R C S T I L L Q G K E Q P L D E E M K E A F Q N A Y L E L G G L G E R V L G F C H F Y L P E E Q Y P K G F A F D C D D V N F A T : 5 6 7 H O M O : P F N S T N K Y Q L S I H E T E D P N D N R Y L L V M K G A P E R I L D R C S T I L L Q G K E Q P L D E E M K E A F Q N A Y L E L G G L G E R V L G F C H Y Y L P E E Q Y P Q G F A F D C D D V N F T T : 5 7 0 * 6 2 0 * 6 4 0 * 6 6 0 * 6 8 0 * 7 0 0 C A L L I N E C T E S : H G L R F V G L M S M I D P P R A A V P D A V A K C R S A G I K V I M V T G D H P I T A K A I A K S V G I I S E G N E T V E D I A Q R L N I P I K E V D P R E A K A A V V H G S E L R D M T S E Q L D D : 6 9 6 D R O S O P H I L A : D N L R F V G L M S M I D P P R A A V P D A V A K C R S A G I K V I M V T G D H P I T A K A I A K S V G I I S E G N E T V E D I A Q R L N I P V S E V N P R E A K A A V V H G A E L R D V S S D Q L D E : 6 5 9 A R T E M I A 1 : T G L R F A G L M S M I D P P R A A V P D A V A K C R S A G I K V I M V T G D H P I T A K A I A K S V G I I S E G N E T V E D I A A R L N I P V S E V N P R D A K A A V V H G G E L R D I T P D A L D E : 6 6 1 A R T E M I A 2 : S G L R F V G L M S M I D P P R A A V P D A V S K C R S A G I K V I M V T G D H P I T A K A I A R Q V G I I S E G H E T V D D I A A R L N I P V S E V N P R S A Q A A V I H G N D L K D M N S D Q L D D : 6 5 5 T O R P E D O : T D L C F V G L M S M I D P P R A A V P D A V G K C R S A G I K V I M V T G D H P I T A K A I A K G V G I I S E G N E T V E D I A A R L N I P V N Q V N P R D A K A C V V H G T D L K D L S H E N L D D : 6 7 9 A N G U I L L A : E N L C F I G L M S M I D P P R A A V L D A V G K C R S P G I K V I M V T G D H P I T A K A I A K G V G I I S E G N E T V E D I A A R L N I P I N E V N P R D A K A C V V H G G E L K D L T P E Q L D D : 6 7 9 X E N O P U S : E N L C F V G L I S M I D P P R A A V P D A V G K C R S A G I K V I M V T G D H P I T A K A I A K G V G I I S E G N E T V E D I A A R L N I P V N Q V N P R D A K A C V I H G T D L K D M T E E Q I D D : 6 8 2 G A L L U S : D N L C F V G L M S M I D P P R A A V P D A V G K C R S A G I K V I M V T G D H P I T A K A I A K G V G I I S E G N E T V E D I A A R L N I P V S Q V N P R D A K A C V I H G T D L K D M S S E Q I D E : 6 6 7 H O M O : D N L C F V G L M S M I G P P R A A V P D A V G K C R S A G I K V I M V T G D H P I T A K A I A K G V G I I S E G N E T V E D I A A R L N I P V S Q V N P R D A K A C V I H G T D L K D F T S E Q I D E : 6 7 0 * 7 2 0 * 7 4 0 * 7 6 0 * 7 8 0 * 8 0 0 C A L L I N E C T E S : V L I H H T E I V F A R T S P Q Q K L I I V E G C Q R M G A I V A V T G D G V N D S P A L K K A D I G V A M G I A G S D V S K Q A A D M I L L D D N F A S I V T G V E E G R L I F D N L K K S I A Y T L : 7 9 6 D R O S O P H I L A : I L R Y H T E I V F A R T S P Q Q K L I I V E G C Q R M G A I V A V T G D G V N D S P A L K K A D I G V A M G I A G S D V S K Q A A D M I L L D D N F A S I V T G V E E G R L I F D N L K K S I A Y T L : 7 5 9 A R T E M I A 1 : I L R H H P E I V F A R T S P Q Q K L I I V E G C Q R Q G A I V A V T G D G V N D S P A L K K A D I G V A M G I A G S D V S K Q A A D M I L L D D N F A S I V T G V E E G R L I F D N L K K S I V Y T L : 7 6 1 A R T E M I A 2 : I L R H Y R E I V F A R T S P Q Q K L I I V E G V Q R Q G E F V A V T G D G V N D S P A L K K A D I G V A M G I A G S D V S K Q A A D M I L L D D N F A S I V T G V E E G R L I F D N I K K S I A Y T L : 7 5 5 T O R P E D O : I L H Y H T E I V F A R T S P Q Q K L I I V E G C Q R Q G A I V A V T G D G V N D S P A L K K A D I G V A M G I A G S D V S K Q A A D M I L L D D N F A S I V T G V E E G R L I F D N L K K S I A Y T L : 7 7 9 A N G U I L L A : I L K H H T E I V F A R T S P Q Q K L I I V E G C Q R Q G A I V A V T G D G V N D S P A L K K A D I G V A M G I A G S D V S K Q A A D M I L L D D N F A S I V T G V E E G R L I F D N L K K S I A Y T L : 7 7 9 X E N O P U S : I L R H H T E I V F A R T S P Q Q K L I I V E G C Q R Q G A I V A V T G D G V N D S P A L K K A D I G I A M G I A G S D V S K Q A A D M I L L D D N F A S I V T G V E E G R L I F D N L K K S I A Y T L : 7 8 2 G A L L U S : I L Q N H T E I V F A R T S P Q Q K L I I V E G C Q R Q G A I V A V T G D G V N D S P A L K K A D I G V A M G I R G S D V S K Q A A D M I L L D D N F A S I V T G V E E G R L I F D N L K K S I A Y T L : 7 6 7 H O M O : I L Q N H T E I V F A R T S P Q Q K L I I V E G C Q R Q G A I V A V T G D G V N D S P A L K K A D I G V A M G I A G S D V S K Q A A D M I L L D D N F A S I V T G V E E G R L I F D N L K K S I A Y T L : 7 7 0 * 8 2 0 * 8 4 0 * 8 6 0 * 8 8 0 * 9 0 0 C A L L I N E C T E S : T S N I P E I S P F L F F M I A S V P L P L G T V T I L C I D L G T D M V P A I S L A Y E E A E S D I M K R Q P R N P F T D K L V N E R L I S M A Y G Q I G M I Q A L A G F Y V Y F V I M A E N G F L P : 8 9 6 D R O S O P H I L A : T S N I P E I S P F L A F I L C D I P L P L G T V T I L C I D L G T D M V P A I S L A Y E H A E A D I M K R P P R D P F N D K L V N S R L I S M A Y G Q I G M I Q A A A G F F V Y F V I M A E N G F L P : 8 5 9 A R T E M I A 1 : T S N I P E I S P F L L F I L F D I P L P L G T V T I L C I D L G T D M V P A I S L A Y E E A E S D I M K R R P R N P V T D K L V N E R L I S L A Y G Q I G M I Q A S A G F F V Y F V I M A E C G F L P : 8 6 1 A R T E M I A 2 : T S K I P E L S P F L M Y I L F D L P L A I G T V T I L C I D L G T D V V P A I S M A Y E G P E A D P R K . . P R D P V K E K L V N E R L I S M A Y G Q I G V M Q A F G G F F T Y F V I M G E C G F L P : 8 5 3 T O R P E D O : T S N I P E I T P F L V F I I A N V P L P L G T V T I L C I D L G T D M V P A I S L A Y E R A E S D I M K R Q P R N P K T D K L V N E R L I S M A Y G Q I G M I Q A L G G F F S Y F V I L A E N G F L P : 8 7 9 A N G U I L L A : T S N I P E I T P F L L F I I A N I P L P L G T V T I L C I D L G T D M V P A I S L A Y E A A E S D I M K R Q P R N P R T D K L V N E R L I S I A Y G Q I G M M Q A T A G F F T Y F V I L A E N G F L P : 8 7 9 X E N O P U S : T S N I P E I T P F L I F I I A N I P L P L G T V T I L C I D L G T D M V P A I S L A Y E Q A E S D I M K R Q P R N P K T D K L V N E R L I S M A Y G Q I G M I Q A L G G F F T Y F V I L A E N G F L P : 8 8 2 G A L L U S : T S N I P E I T P F L L F I M A N I P L P L G T I T I L C I D L G T D M V P A I S L A Y E A A E S D I M K R Q P R N P R S D K L V N E R L I S M A Y G Q I G M I Q A L G G F F S Y F V I L A E N G F L P : 8 6 7 H O M O : T S N I P E I T P F L L F I M A N I P L P L G T I T I L C I D L G T D M V P A I S L A Y E A A E S D I M K R Q P R N P R T D K L V N E R L I S M A Y G Q I G M I Q A L G G F F S Y F V I L A E N G F L P : 8 7 0 * 9 2 0 * 9 4 0 * 9 6 0 * 9 8 0 * 1 0 0 0 C A L L I N E C T E S : P V L F G I R E Q W D S K A I N D L E D Y Y G Q E W T Y H D R K I L E Y T C H T A F F V A I V V V Q W A D L I I C K T R R N S I L H Q G M K N M V L N F G L C F E T T L A A F L S Y T P G M D K G L R M : 9 9 6 D R O S O P H I L A : K K L F G I R K M W D S K A V N D L T D S Y G Q E W T Y R D R K T L E Y T C H T A F F I S I V V V Q W A D L I I C K T R R N S I F Q Q G M R N W A L N F G L V F E T V L A A F L S Y C P G M E K G L R M : 9 5 9 A R T E M I A 1 : W D L F G L R K H W D S R A V N D L T D S Y G Q E W T Y D A R K Q L E S S C H T A Y F V S I V I V Q W A D L I I S K T R R N S V F Q Q G M R N N I L N F A L V F E T C L A A F L S Y T P G M D K G L R M : 9 6 1 A R T E M I A 2 : N R L F G L R K W W E S K A Y N D L T D S Y G Q E W T W D A R K Q L E Y T C H T A F F I S I V I V Q W T D L I I C K T R R L S L F Q Q G M K N G T L N F A L V F E T C V A A F L S Y T P G M D K G L R M : 9 5 3 T O R P E D O : I D L I G I R E K W D E L W T Q D L E D S Y G Q Q W T Y E Q R K I V E Y T C H T S F F V S I V I V Q W A D L I I C K T R R N S I F Q Q G M K N K I L I F G L F E E T A L A A F L S Y T P G T D I A L R M : 9 7 9 A N G U I L L A : S T L L G I R V K W D D K Y V N D L E D S Y G Q Q W T Y E Q R K I V E Y T C H T S F F A S I V I V Q W A D L I I C K T R R N S I I Q Q G M K N K I L I F G L F E E T A L A A F L S Y C P G M D V A L R M : 9 7 9 X E N O P U S : W T L L G I R V N W D D R W T N D V E D S Y G Q Q W T Y E Q R K I V E F T C H T S F F I S I V V V Q W A D L I I C K T R R N S V F Q Q G M K N K I L I F G L F E E T A L A A F L S Y C P G M D V A L R M : 9 8 2 G A L L U S : S C L V G I R L S W D D R T I N D L E D S Y G Q Q W T Y E Q R K V V E F T C H T A F F V S I V V V Q W A D L I I C K T R R N S V F Q Q G M K N K I L I F G L F E E T A L A A F L S Y C P G M D V A L R M : 9 6 7 H O M O : G N L V G I R L N W D D R T V N D L E D S Y G Q Q W T Y E Q R K V V E F T C H T A F F V S I V V V Q W A D L I I C K T R R N S V F Q Q G M K N K I L I F G L F E E T A L A A F L S Y C P G M D V A L R M : 9 7 0
* 1 0 2 0 * 1 0 4 0 C A L L I N E C T E S : Y P L K F Y W W L P P L P F S L L I F V Y D E C R R F V L R R N P G G W V E M E T Y Y : 1 0 3 9 D R O S O P H I L A : Y P L K L V W W F P A I P F A L A I F I Y D E T R R F Y L R R N P G G W L E Q E T Y Y : 1 0 0 2 A R T E M I A 1 : Y P L K I N W W F P A L P F S F L I F V Y D E A R K F I L R R N P G G W V E Q E T Y Y : 1 0 0 4 A R T E M I A 2 : Y P L K I W W W F P P M P F S L L I L V Y D E C R K F L M R R N P G G F L E R E T Y Y : 9 9 6 T O R P E D O : Y P L K P S W W F C A F P Y S L I I F L Y D E A R R F I L R R N P G G W V E Q E T Y Y : 1 0 2 2 A N G U I L L A : Y P L K P S W W F C A F P Y S L L I F L Y D E A R R F I L R R N P D G W V E R E T Y Y : 1 0 2 2 X E N O P U S : Y P L K P T W W F C A F P Y S L I I F I Y D E V R K L I I R R S P G G W V E K E S Y Y : 1 0 2 5 G A L L U S : Y P L K P S W W F C A F P Y S F L I F V Y D E I R K L I L R R N P G G W V E K E T Y Y : 1 0 1 0 H O M O : Y P L K P S W W F C A F P Y S F L I F V Y D E I R K L I L R R N P G G W V E K E T Y Y : 1 0 1 3
D L
[image:3.612.88.531.71.708.2]T
Fig. 2. Multiple alignment of the Callinectes sapidus
α
-subunit
amino acid sequence with examples from other arthropods
(Drosophila melanogaster and two isoforms from Artemia
franciscana) and from vertebrates (Torpedo californica,
Anguilla anguilla, Xenopus laevis, Gallus gallus and Homo
sapiens). Alignment was produced using ClustalW and GeneDoc software. Blue, 100 % agreement; green, 80 %; yellow, 60 %. Putative
Callinectes sapidus approximately doubles following transfer
of the animals from 35 to 5 ‰ salinity (Neufeld et al., 1980;
Piller et al., 1995; Towle et al., 1976). The increased Na
+
+K
+
-ATPase activity is thought to drive uptake of Na
+
and Cl
−
across
the gill epithelium, leading to hyperosmoregulation of the
hemolymph in low-salinity environments. However, it remains
unclear whether the activity of pre-existing Na
+
+K
+
-ATPase
molecules is regulated in response to a change in salinity or
whether the observed increase in enzymatic activity (and
presumably in pumping activity) may be the result of enhanced
gene transcription and/or translation. In the present study, we
used reverse transcription and the polymerase chain reaction
(RT-PCR) to identify and characterize Na
+
+K
+
-ATPase
α
-subunit cDNA prepared from gills of the euryhaline blue crab
Callinectes sapidus. Quantitative RT-PCR, western blotting
and immunocytochemistry were employed to investigate the
correlation between
α
-subunit mRNA and protein abundance in
relation to the osmoregulatory response. Some of our data have
appeared in abstract form (Paulsen et al., 2000).
Materials and methods
Blue crabs (Callinectes sapidus Rathbun) were obtained from
Gulf Specimens, Panacea, Florida, USA, and were maintained
in recirculating biologically filtered aquaria at 20 °C containing
natural sea water at salinities of either 35 or 5 ‰. Crabs were
fed cleaned mussels twice weekly. Prior to the removal of the
gills, the crabs were anesthetized on ice for 30 min.
Total RNA was extracted from the gills under RNAse-free
conditions using materials provided by Promega Corporation
(Chomczynski and Sacchi, 1987). Poly(A) mRNA in the total
RNA preparation was reverse-transcribed to cDNA using
oligo-dT primer and Superscript II reverse transcriptase (Life
Technologies). Degenerate primers for the polymerase chain
reaction (PCR) were based on conserved regions of published
Na
+
+K
+
-ATPase
α
-subunit sequences from other arthropods,
including Drosophila melanogaster (Lebovitz et al., 1989) and
Artemia franciscana (Baxter-Lowe et al., 1989; Macías et al.,
1991). Primers were designed with the assistance of Primer
Premier software and synthesized by Operon Technologies.
Polymerase chain reactions were carried out in an MJ Research
thermocycler using Sigma RedTaq DNA polymerase and
nucleotides. The usual incubation protocol included an initial
denaturation at 92 °C for 5 min followed by cooling to 60 °C
and addition of the polymerase. Using degenerate primers, the
reaction tubes were cycled 30 times through 1 min at 92 °C,
1 min at 45 °C and 2 min at 72 °C, followed by an extension at
72 °C for 5 min and storage at 4 °C. With non-degenerate
primers, the annealing temperature was raised from 45 to
55 °C. PCR products were separated electrophoretically on
agarose gels, visualized by ethidium bromide staining and
extracted from the gels using the QiaQuick protocol (Qiagen).
An initial 700-base-pair PCR product obtained with
degenerate primers NAK10F (5
′
-ATGACIGTIGCICAYATG-TGG-3
′
) and NAK16R (5
′
-GGRTGRTCICCIGTIACCAT-3
′
)
was sequenced at the Marine DNA Sequencing Center of the
Mount Desert Island Biological Laboratory using ABI 377 and
3100 automated sequencers. Callinectes-specific primers were
then designed to complete the sequencing with the aid of 3
′
rapid amplification of cDNA ends (3
′
-RACE) (Life
Technologies) and 5
′
-RACE (Ambion) techniques. Sequence
assembly and analysis were performed with DNASTAR and
DNASIS software packages. Comparison with published
sequences in GenBank was made with the BLAST algorithm
(Altschul et al., 1997) and multiple alignments were produced
with ClustalW (http://antheprot-pbil.ibcp.fr/ie_sommaire.html)
and GeneDoc software (http://www.psc.edu/biomed/genedoc/).
Putative transmembrane regions were identified by
hydrophobicity analysis using AnTheProt
(http://antheprot-pbil.ibcp.fr/ie_sommaire.html).
The abundance of
α
-subunit mRNA in total RNA extracts
was estimated by quantitative RT-PCR using identical amounts
of total RNA in each reverse transcription reaction. The
polymerase chain reaction was carried out with biotinylated
dUTP replacing a portion of the dTTP, under conditions in
which product formation was directly dependent on cDNA
template availability (Towle et al., 1997). Following transfer to
nylon membranes, biotinylated products were visualized with
the Phototope protocol (New England Biolabs). A lower limit
of considerably less than a twofold difference in mRNA
abundance can be detected using this method (Weihrauch et al.,
2001).
Identification of
α
-subunit protein was achieved by western
blotting of partially purified plasma membranes (Lucu
and Flik, 1999) and detection with the
α
5 monoclonal
antibody against a highly conserved cytosolic epitope of the
α
-subunit of avian Na
+
+K
+
-ATPase (Lebovitz et al., 1989).
The
α
5 antibody was obtained from Developmental Studies
Hybridoma Bank or was kindly provided by Dr D. M.
Fambrough. Identical quantities of membrane protein were
loaded into each well of the polyacrylamide gel, as assayed
using Coomassie Blue binding and bovine serum albumin as
standard (Bradford, 1976).
Immunocytochemical localization of
α
-subunit protein was
carried out with the
α
5 antibody using previously published
methods (Lignot et al., 1999; Ziegler, 1997) following fixation
in Bouin’s fixative. Transverse sections (3
µ
m) were incubated
in primary antibody, followed by washing and incubation in
secondary antibody (fluorescein-isothiocyanate-conjugated
goat anti-mouse IgG; Jackson Immunoresearch), and examined
in a Leitz Diaplan fluorescence microscope. Controls included
omitting primary antibody, revealing only a low background
of autofluorescence.
Results
Amplification of the
α
-subunit cDNA from total RNA of C.
sapidus gill was accomplished initially with degenerate
primers NAK10F and NAK16R, yielding a 700-base-pair
product. After sequencing and a BLAST search of GenBank,
the initial PCR product was clearly identified as encoding a
4009
Molecular physiology of crab gill Na
+K
-ATPase
Callinectes-specific primers enabled the amplification of the
complete
α
-subunit cDNA using 3
′
- and 5
′
-RACE techniques.
The resulting 3828-nucleotide cDNA encodes a putative
1039-amino-acid polypeptide with a predicted molecular mass
of
115.6 kDa (Fig. 1). A Kozak consensus sequence
(CAACCATGG) brackets the likely start codon, an A
replacing the second position C in the most conserved
arrangement (Kozak, 1991). Several stop codons, one
in-frame, occur prior to the putative start site. The predicted
translational stop site is followed by six nucleotides and then
two additional in-frame stop codons. A polyadenylation signal
(ATTAAA, the second-most common polyadenylation signal)
(Wickens, 1990) precedes the start of the poly-A tail by 13
nucleotides. No evidence of additional isoforms or alternative
splicing products was observed.
Hydrophobicity analysis of the
α
-subunit amino acid
sequence led to a prediction of eight transmembrane domains
(Fig. 1), in agreement with other topological models of the
Na
+
+K
+
-ATPase
α
-subunit (for a review, see Horisberger et
al., 1991). The C. sapidus
α
-subunit amino acid sequence was
aligned with
α
-subunit sequences from other invertebrates
including Artemia franciscana (Baxter-Lowe et al., 1989;
Macías et al., 1991) and Drosophila melanogaster (GenBank
Accession No. AF044974) and vertebrate species including
Torpedo californica (Kawakami et al., 1985), Anguilla
anguilla (Cutler et al., 1995), Xenopus laevis (Verrey et al.,
1989), Gallus gallus (Takeyasu et al., 1990) and Homo sapiens
(Ovchinnikov et al., 1988). The multiple alignment reveals
extensive regions of highly conserved amino acid sequence,
particularly in the fourth predicted transmembrane region and
in the putative cytosolic domain, which includes the ATP
binding site (Fig. 2). Statistical analysis of the alignment using
GeneDoc software showed 81–83 % amino acid identity of the
C. sapidus
α
-subunit sequence with other arthropod
α
-subunit
sequences and 74–77 % identity with vertebrate sequences.
The posterior ion-transporting gills of euryhaline crabs are
known to have substantially higher Na
+
+K
+
-ATPase enzymatic
activity than the anterior respiratory gills. Furthermore, the
enzymatic activity in the posterior gills is enhanced upon
transfer of crabs from high to low salinity (for reviews, see
Towle, 1990; Péqueux, 1995). To determine whether gene
activation might be responsible for the observed differences in
enzymatic activity, we measured the abundance of
α
-subunit
mRNA using quantitative RT-PCR. Using arginine kinase
mRNA as an invariant internal standard for comparison
(Kotlyar et al., 2000), we found that the relative proportion of
Na
+
+K
+
-ATPase
α
-subunit mRNA is much higher in the
posterior gills (gills 6–8) than in the anterior gills (gills 3–5)
(Fig. 3), supporting the conclusion that transcriptional
regulation or differential mRNA stability may be responsible
for the gill differences in Na
+
+K
+
-ATPase enzymatic activity.
However, little difference in
α
-subunit mRNA abundance was
noted following transfer from high (35 ‰) to low (5 ‰) salinity
(Fig. 3). Although it is likely that our quantitative RT-PCR
technique was insufficiently sensitive to recognize small
differences in mRNA abundance, it is clear that the doubling of
enzymatic activity following a reduction in salinity (Neufeld et
al., 1980; Piller et al., 1995; Towle et al., 1976) is not the result
of a doubling of
α
-subunit mRNA levels.
The
α
-subunit protein was identified in C. sapidus gills by
western blotting of partially purified plasma membrane proteins
separated by SDS–polyacrylamide gel electrophoresis. The
α
5 antibody detected a single polypeptide of approximately
114 kDa (on the basis of a comparison with Bio-Rad
Kaleidoscope prestained standards) (Fig. 4), consistent with the
size of the
α
-subunit protein predicted from the cDNA
sequence. Membrane preparations from the posterior gills
contained a much greater proportion of
α
-subunit protein than
those from the anterior gills, paralleling our estimations of
α
-subunit mRNA abundance. However, acclimation salinity
appeared to have little effect on the relative amount of
α
-subunit
protein in the posterior gills (Fig. 4). Although visual inspection
suggested a modest increase in levels of the
α
-subunit protein
[image:5.612.203.568.534.698.2]following transfer from 35 to 5 ‰ salinity, digitizing the band
Fig. 3. Estimate of Na
+
+K
+
-ATPase
α
-subunit mRNA abundance in total
RNA extracts of anterior and posterior
gills of Callinectes sapidus acclimated
for at least 2 weeks to 35 or 5 ‰
salinity. Gills 3–5 (anterior) and 6–8
(posterior) were pooled from three
individuals for each RNA preparation.
mRNA levels were evaluated by
duplex quantitative RT-PCR under
conditions of limiting template, with
arginine kinase mRNA serving as an
invariant standard (Kotlyar et al.,
2000). The primers employed to
amplify Na
+
+K
+
-ATPase
α
-subunit
cDNA were NAK10F and NAK16R
and for arginine kinase AKF5 (5
′
-CGCTGAGTCTAAGAAGGGATT-3
′
) and AKCALLR1 (5
′
-CCCAGGCTTGTCTTCTTGTCC-3
′
).
Biotinylated PCR products were visualized after 22 cycles using a streptavidin/alkaline phosphatase procedure (Phototope, New England
Biolabs). The data are representative of three separate experiments.
Na
+
+K
+
-ATPase
α
-subunit mRNA
Arginine kinase mRNA
35
5
35
5
DNA marker
Anterior
Posterior
densities with NIH Image software (Scion Corporation) failed
to support such a conclusion.
Immunocytochemical analysis of
α
-subunit localization in
cross sections of posterior gill lamellae indicated its
predominance in basolateral membrane regions of the
epithelium and its absence from the apical region (Fig. 5), in
agreement with earlier ultracytochemical studies (Towle and
Kays, 1986). Comparison of immunostaining between crabs
acclimated to 35 or 5 ‰ salinity revealed little difference in
intensity, consistent with western blot analysis of
α
-subunit
protein abundance. An examination of anterior gills by
immunocytochemistry indicated very low levels of
α
-subunit
protein (data not shown).
Discussion
Ion-transporting cells in the branchial epithelium of
brachyuran crabs are identified by an abundance of
mitochondria and high specific Na
+
+K
+
-ATPase activity
localized in the basolateral membrane (Péqueux, 1995; Towle
and Kays, 1986). These cells are arranged in a recognizably
dark-colored ‘patch’ adjacent to the afferent blood vessel in
each lamella of the posterior gills (Barra et al., 1983; Compère
et al., 1989; Copeland and Fitzjarrell, 1968). The anterior gills
are essentially devoid of the ion-transporting cell type and are
thought to play a major role in respiratory processes (Taylor
and Taylor, 1992). The area of the dark-colored patch in the
posterior gills is known to increase following transfer of blue
crabs from high to low salinity, reaching a maximum at
approximately 7 days post-transfer (Aldridge and Cameron,
1982; Towle and Burnett, 2001). Thus, it might be expected
that Na
+
+K
+
-ATPase activity would demonstrate a parallel
increase following a reduction in environmental salinity;
indeed, such a change has been documented in a broad variety
of euryhaline crab species (for reviews, see Harris and Bayliss,
1988; Lucu, 1993; Péqueux, 1995; Towle, 1990).
Despite numerous studies of salinity-related changes in
Na
+
+K
+
-ATPase activity in crustacean gills, it remains unclear
whether the measured increases in activity result from de
novo synthesis of Na
+
+K
+
-ATPase mRNA and/or protein or
from post-translational processes such as subunit assembly,
membrane trafficking or cell signaling. Amplification and
sequencing of the
α
-subunit cDNA has allowed us to assess
whether differences in mRNA availability might explain some
of the variation in enzymatic activity. It is clear that the high
Na
+
+K
+
-ATPase activity in the posterior gills compared with
the anterior gills can be explained largely by the much greater
abundance of
α
-subunit mRNA in total RNA extracts of
posterior gills. Whether the high level of
α
-subunit mRNA in
[image:6.612.324.551.72.374.2]the posterior gills is the result of higher gene transcription rates
Fig. 4. Western blot analysis of Na
+
+K
+
-ATPase
α
-subunit protein
in partially purified plasma membrane preparations from anterior and
posterior gills of Callinectes sapidus acclimated for 2 weeks to 35 or
5 ‰ salinity. Gills 3–5 (anterior) and 6–8 (posterior) from four crabs
were pooled for each treatment. Following SDS–polyacrylamide gel
electrophoresis and transfer to nylon membranes,
α
-subunit protein
was detected by incubation with
α
5 monoclonal antibody. The
left-hand lane contains Kaleidoscope prestained standards with the
indicated molecular masses. The data are representative of three
separate experiments.
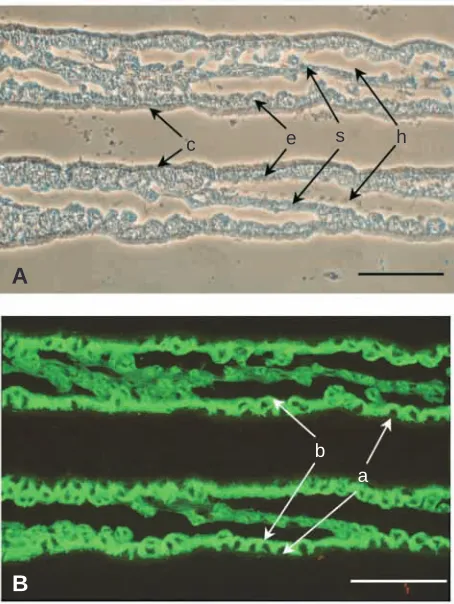
Fig. 5. Immunocytochemical localization of Na
+
+K
+
-ATPase
α
-subunit protein in the basolateral membrane of gill epithelial cells in
the blue crab Callinectes sapidus. (A) Phase-contrast
photomicrograph of portions of two cross-sectioned gill lamellae. c,
cuticle; e, epithelial layer; s, intralamellar septum; h, hemolymph
space. (B) Immunocytochemical identification of
α
-subunit protein
using the
α
5 monoclonal antibody against a highly conserved
cytosolic epitope. Intense fluorescence is evident in the basolateral
membrane region (b) but not the apical membrane region (a) of the
epithelial cells. Scale bars, 100
µ
m.
78 kDa
121 kDa
204 kDa
35
5
35
5
Anterior
Posterior
Salinity (‰)
A
c
e
s
h
B
[image:6.612.47.289.74.234.2]4011
Molecular physiology of crab gill Na
+K
-ATPase
or of greater mRNA stability cannot be assessed by our
experiments. However, the 3
′
untranslated region of the
α
-subunit cDNA contains no adenosine- and uridine-rich
instability elements such as are found in short-lived mRNAs
(Chen and Shyu, 1995), suggesting that this mRNA may be
quite long-lived.
Identification of the Callinectes sapidus Na
+
+K
+
-ATPase
α
-subunit protein with a heterologous antibody against the avian
α
-subunit revealed a single protein band of approximately the
expected size on western blots. Because the monoclonal
α
5
antibody is directed against the highly conserved cytosolic
domain of the
α
-subunit, it is not surprising that it would react
strongly with the crustacean protein. Indeed, the antibody has
been employed to characterize the
α
-subunit of another
arthropod, Drosophila melanogaster (Lebovitz et al., 1989).
We can therefore be confident that the
α
5 antibody is detecting
the appropriate protein in the western blots of C. sapidus gill
cell membranes. The detected band is located at approximately
114 kDa, in close agreement with the size of the
α
-subunit
protein (115.6 kDa) predicted from the cDNA sequence. The
α
5 antibody also identifies a protein that is restricted to the
basolateral membrane of the gill epithelial cells, in agreement
with the previously demonstrated ultracytochemical
localization of the Na
+
+K
+
-ATPase (Towle and Kays, 1986).
A comparson of the relative abundance of
α
-subunit protein
in membranes from posterior and anterior gills showed clearly
that the posterior gills contain a much higher level of
α
-subunit
protein, consistent with the higher expression of
α
-subunit
mRNA in posterior gills. We tentatively conclude on the basis
of these observations that the process of cellular differentiation
leading to the production and maintenance of ion-transporting
cells in the posterior gill epithelium depends at least in part on
transcriptional activation of the
α
-subunit gene in these cells,
yielding high levels of both mRNA and protein. Assembly of
the
α
-subunit protein with the
β
and possibly
γ
subunits (if the
latter exists in the Na
+
+K
+
-ATPase of C. sapidus) and
targeting to the basolateral membrane would yield a functional
pump protein poised to energize Na
+
uptake (and NH
4
+
excretion) across the posterior gill epithelium. Reduced
transcription of the
α
-subunit gene in the anterior gills would
lead to specializations other than ion transport.
Acclimation salinity appears to have little effect on the
expression of
α
-subunit mRNA and protein in the gills of the
blue crab. Although it is difficult to make strictly quantitative
determinations using either the RT-PCR technique or western
blotting, we have shown that a twofold difference in mRNA
abundance is easily detected by our RT-PCR method
(Weihrauch et al., 2001). If variation in
α
-subunit mRNA
availability were the underlying cause of the reported
differences in Na
+
+K
+
-ATPase activity, we should have
detected at least some change in mRNA levels. Under our
experimental conditions, neither
α
-subunit mRNA nor protein
levels appeared to vary very much with salinity.
Because we standardized both the RT-PCR method and the
western blot with equivalent amounts of total RNA
(reverse-transcribed to cDNA) and membrane protein, respectively, it
is conceivable that our approach masked the differentiation of
additional ion-transporting cells in the gill epithelium. Such an
ontogenetic change following a reduction in salinity would
probably recruit a large number of transport and regulatory
proteins involved in transepithelial ion movements, with the
α
-subunit as one component of the recruitment process. Thus, the
relative proportion of the
α
-subunit to total RNA and protein
could remain constant even in the event of substantial
upregulation of transcription and translation.
However, most published measurements of Na
+
+K
+
-ATPase activity are specific activity measurements, factoring
in the concentration of total protein in the homogenate or
membrane fraction. Thus, the apparent lack of change in
α
-subunit protein content with salinity suggests that
post-translational processes may play an important role in the
response of Na
+
+K
+
-ATPase activity to salinity change. In
mammalian tissues, a wide variety of mechanisms regulating
the Na
+
+K
+
-ATPase have been studied. These include
interaction with the
γ
-subunit and cytoskeletal elements,
endogenous ouabain-like inhibitors and hormones that may act
through protein kinases and phosphatases (for a review, see
Therien and Blostein, 2000).
Studies of crustacean gills have revealed several candidate
regulatory molecules that may participate in controlling the
function of the Na
+
+K
+
-ATPase. Dopamine and cyclic AMP
have been implicated by several authors (Bianchini and Gilles,
1990; Lucu and Flik, 1999; Mo et al., 1998; Sommer and
Mantel, 1988, 1991). A sinus gland component, later identified
as crustacean hyperglycemic hormone, has been shown to
enhance Na
+
transport and Na
+
+K
+
-ATPase activity in
perfused gills of the intertidal crab Pachygrapsus marmoratus
(Eckhardt et al., 1995; Spanings-Pierrot et al., 2000). Methyl
farnesoate levels increased in the hemolymph of Carcinus
maenas following transfer from sea water to dilute salinity
(Lovett et al., 2001); whether levels of Na
+
+K
+
-ATPase
activity are affected by this molecule is unknown. Future
studies are therefore necessary to identify the
transport-regulating factors and their mode of action on the Na
+
+K
+
-ATPase as well as other ion transporters.
This research was supported by a National Science
Foundation grant to D.W.T. (IBN-9807539) and an NSF
Research Experiences for Undergraduates grant to MDIBL
(DBI-9820400). The authors wish to thank Christine Smith,
supervisor of the Marine DNA Sequencing Center at MDIBL,
for her excellent and timely assistance. The Marine DNA
Sequencing Center is supported by a grant from the Maine
Science and Technology Foundation.
References
Aldridge, J. B. and Cameron, J. N. (1982). Gill morphometry in the blue
crab, Callinectes sapidus Rathbun (Decapoda Brachyura). Crustaceana 43,
297–305.
Altschul, S. F., Madden, T. L., Schaffer, A. A., Zhang, J., Zhang, Z.,
Miller, W. and Lipman, D. J. (1997). Gapped BLAST and PSI-BLAST:
a new generation of protein database search programs. Nucleic Acids Res.
Barra, J. A., Péqueux, A. and Humbert, W. (1983). A morphological study
on gills of a crab acclimated to fresh water. Tissue Cell 15, 583–596.
Baxter-Lowe, L. A., Guo, J. Z., Bergstrom, E. E. and Hokin, L. E. (1989).
Molecular cloning of the Na,K-ATPase
α
-subunit in developing brine
shrimp and sequence comparison with higher organisms. FEBS Lett. 257,
181–187.
Bianchini, A. and Gilles, R. (1990). Cyclic AMP as a modulator of NaCl
transport in gills of the euryhaline Chinese crab Eriocheir sinensis. Mar.
Biol. 104, 191–196.
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein–dye
binding. Anal. Biochem. 72, 248–254.
Chen, C. Y. and Shyu, A. B. (1995). AU-rich elements: characterization and
importance in mRNA degradation. Trends Biochem. Sci. 20, 465–470.
Chomczynski, P. and Sacchi, N. (1987). Single-step method of RNA
isolation by acid guanidinium thiocyanate–phenol–chloroform extraction.
Anal. Biochem. 162, 156–159.
Compère, P., Wanson, S., Péqueux, A., Gilles, R. and Goffinet, G. (1989).
Ultrastructural changes in the gill epithelium of the green crab Carcinus
maenas in relation to the external salinity. Tissue Cell 21, 299–318.
Copeland, D. E. and Fitzjarrell, A. T. (1968). The salt absorbing cells in the
gills of the blue crab (Callinectes sapidus Rathbun) with notes on modified
mitochondria. Z. Zellforsch. 92, 1–22.
Cutler, C. P., Sanders, I. L., Hazon, N. and Cramb, G. (1995). Primary
sequence, tissue specificity and expression of the Na
+
,K
+
-ATPase alpha 1
subunit in the European eel (Anguilla anguilla). Comp. Biochem. Physiol.
111B, 567–573.
Eckhardt, E., Pierrot, C., Thuet, P., Van Herp, F., Charmantier-Daures,
M., Trilles, J. P. and Charmantier, G. (1995). Stimulation of
osmoregulating processes in the perfused gill of the crab Pachygrapsus
marmoratus (Crustacea, Decapoda) by a sinus gland peptide. Gen. Comp.
Endocrinol. 99, 169–177.
Harris, R. R. and Bayliss, D. (1988). Gill (Na
+
+K
+
)-ATPases in decapod
crustaceans: Distribution and characteristics in relation to Na
+
regulation.
Comp. Biochem. Physiol. 90A, 303–308.
Horisberger, J.-D., Lemas, V., Kraehenbühl, J.-P. and Rossier, B. C.
(1991). Structure–function relationship of Na,K-ATPase. Annu. Rev.
Physiol. 53, 565–584.
Kawakami, K., Noguchi, S., Noda, M., Takahashi, H., Ohta, T.,
Kawamura, M., Nojima, H., Nagano, K., Hirose, T., Inayama, S. et al.
(1985). Primary structure of the alpha-subunit of Torpedo californica
(Na
+
+K
+
)ATPase deduced from cDNA sequence. Nature 316, 733–736.
Kotlyar, S., Weihrauch, D., Paulsen, R. S. and Towle, D. W. (2000).
Expression of arginine kinase enzymatic activity and mRNA in gills of the
euryhaline crabs Carcinus maenas and Callinectes sapidus. J. Exp. Biol.
203, 2395–2404.
Kozak, M. (1991). Structural features in eukaryotic mRNAs that modulate the
initiation of translation. J. Biol. Chem. 266, 19867–19870.
Lebovitz, R. M., Takeyasu, K. and Fambrough, D. M. (1989). Molecular
characterization and expression of the (Na
+
+K
+
)-ATPase alpha-subunit in
Drosophila melanogaster. EMBO J. 8, 193–202.
Lignot, J. H., Charmantier-Daures, M. and Charmantier, G. (1999).
Immunolocalization of Na
+
,K
+
-ATPase in the organs of the branchial cavity
of the European lobster Homarus gammarus (Crustacea, Decapoda). Cell
Tissue Res. 296, 417–426.
Lovett, D. L., Verzi, M. P., Clifford, P. D. and Borst, D. W. (2001).
Hemolymph levels of methyl farnesoate increase in response to osmotic
stress in the green crab, Carcinus maenas. Comp. Biochem. Physiol. 128A,
299–306.
Lucu, C
ˇ . (1993). Ion transport in gill epithelium of aquatic Crustacea. J. Exp.
Zool. 265, 378–386.
Lucu, C
ˇ . and Flik, G. (1999). Na
+
–K
+
-ATPase and Na
+
/Ca
2+
exchange
activities in gills of hyperregulating Carcinus maenas. Am. J. Physiol. 276,
R490–R499.
Macías, M.-T., Palmero, I. and Sastre, L. (1991). Cloning of a cDNA
encoding an Artemia franciscana Na/K ATPase
α
-subunit. Gene 105,
197–204.
Mo, J. L., Devos, P. and Trausch, G. (1998). Dopamine as a modulator of
ionic transport and Na
+
/K
+
-ATPase activity in the gills of the Chinese crab
Eriocheir sinensis. J. Crust. Biol. 18, 442–448.
Neufeld, G. J., Holliday, C. W. and Pritchard, J. B. (1980). Salinity
adaption of gill Na,K-ATPase in the blue crab, Callinectes sapidus. J. Exp.
Zool. 211, 215–224.
Ovchinnikov, Y. A., Monastyrskaya, G. S., Broude, N. E., Ushkaryov, Y.
A., Melkov, A. M., Smirnov, Y. V., Malyshev, I. V., Allikmets, R. L.,
Kostina, M. B., Dulubova, I. E. et al. (1988). Family of human Na
+
,K
+
-ATPase genes. Structure of the gene for the catalytic subunit (alpha
III-form) and its relationship with structural features of the protein. FEBS Lett
233, 87–94.
Paulsen, R. S., Weihrauch, D., Lignot, J.-H. and Towle, D. W. (2000).
Na,K-ATPase in gills of the blue crab Callinectes sapidus: cDNA
sequencing and salinity-related expression of alpha subunit mRNA and
protein. Am. Zool. 40, 1166–1167.
Péqueux, A. (1995). Osmotic regulation in crustaceans. J. Crust. Biol. 15,
1–60.
Piller, S. C., Henry, R. P., Doeller, J. E. and Kraus, D. W. (1995). A
comparison of the gill physiology of two euryhaline crab species,
Callinectes sapidus and Callinectes similis: energy production,
transport-related enzymes and osmoregulation as a function of acclimation salinity.
J. Exp. Biol. 198, 349–358.
Shull, G. E., Lane, L. K. and Lingrel, J. B. (1986). Amino-acid sequence of
the beta-subunit of the (Na
+
+K
+
)ATPase deduced from a cDNA. Nature
321, 429–431.
Shull, G. E., Schwartz, A. and Lingrel, J. B. (1985). Amino-acid sequence
of the catalytic subunit of the (Na
+
+K
+
)ATPase deduced from a
complementary DNA. Nature 316, 691–695.
Skou, J. C. (1957). The influence of some cations on an adenosine
triphosphatase from peripheral nerves. Biochim. Biophys. Acta 23, 394–401.
Sommer, M. J. and Mantel, L. H. (1988). Effect of dopamine, cyclic AMP
and pericardial organs on sodium uptake and Na+K-ATPase activity in gills
of the green crab Carcinus maenas (L). J. Exp. Zool. 248, 272–277.
Sommer, M. J. and Mantel, L. H. (1991). Effects of dopamine and
acclimation to reduced salinity on the concentration of cyclic AMP in the
gills of the green crab, Carcinus maenas (L). Gen. Comp. Endocrinol. 82,
364–368.
Spanings-Pierrot, C., Soyez, D., Van Herp, F., Gompel, M., Skaret, G.,
Grousset, E. and Charmantier, G. (2000). Involvement of crustacean
hyperglycemic hormone in the control of gill ion transport in the crab
Pachygrapsus marmoratus. Gen. Comp. Endocrinol. 119, 340–350.
Takeyasu, K., Lemas, V. and Fambrough, D. M. (1990). Stability of
Na
+
–K
+
-ATPase
α
-subunit isoforms in evolution. Am. J. Physiol. 259,
C619–C630.
Taylor, H. H. and Taylor, E. W. (1992). Gills and lungs: The exchange of
gases and ions. In Microscopic Anatomy of Invertebrates, vol. 10. Decapod
Crustacea (ed. F. W. Harrison and A. G. Humes), pp. 203–293. New York:
Wiley-Liss.
Therien, A. G. and Blostein, R. (2000). Mechanisms of sodium pump
regulation. Am. J. Physiol. 279, C541–C566.
Towle, D. W. (1990). Sodium transport systems in gills. In Comparative
Aspects of Sodium Cotransport Systems (ed. R. K. H. Kinne), pp. 241–263.
Basel: Karger Publishing.
Towle, D. W. and Burnett, L. E. (2001). Osmoregulatory, digestive and
respiratory physiology. In The Blue Crab Callinectes sapidus (ed. V.
Kennedy). Cambridge, MD: Maryland Sea Grant (in press).
Towle, D. W. and Kays, W. T. (1986). Basolateral localization of Na
+
+K
+
-ATPase in gill epithelium of two osmoregulating crabs, Callinectes sapidus
and Carcinus maenas. J. Exp. Zool. 239, 311–318.
Towle, D. W., Palmer, G. E. and Harris III, J. L. (1976). Role of gill
Na
+
+K
+
-dependent ATPase in acclimation of blue crabs (Callinectes
sapidus) to low salinity. J. Exp. Zool. 196, 315–322.
Towle, D. W., Rushton, M. E., Heidysch, D., Magnani, J. J., Rose, M. J.,
Amstutz, A., Jordan, M. K., Shearer, D. W. and Wu, W. S. (1997).
Sodium–proton antiporter in the euryhaline crab Carcinus maenas:
Molecular cloning, expression and tissue distribution. J. Exp. Biol. 200,
1003–1014.
Verrey, F., Kairouz, P., Schaerer, E., Fuentes, P., Geering, K., Rossier, B.
C. and Kraehenbuhl, J.-P. (1989). Primary sequence of Xenopus laevis
Na
+
–K
+
-ATPase and its localization in A6 kidney cells. Am. J. Physiol. 256,
F1034–F1043.
Weihrauch, D., Ziegler, A., Siebers, D. and Towle, D. W. (2001). Molecular
characterization of V-type H
+
-ATPase (B-subunit) in gills of euryhaline
crabs and its physiological role in osmore