1556-6811/08/$08.00⫹0 doi:10.1128/CVI.00008-08
Copyright © 2008, American Society for Microbiology. All Rights Reserved.
Development of a Novel Efficient Fluorescence-Based Plaque Reduction
Microneutralization Assay for Measles Virus Immunity
䌤
Iana H. Haralambieva,
1Inna G. Ovsyannikova,
1Robert A. Vierkant,
2and Gregory A. Poland
1,3*
Mayo Vaccine Research Group, Mayo Clinic College of Medicine, Rochester, Minnesota 559051; Department of Health Sciences Research,
Mayo Clinic College of Medicine, Rochester, Minnesota 559052; and Program in Translational Immunovirology and
Biodefense, Mayo Clinic College of Medicine, Rochester, Minnesota 559053
Received 21 December 2007/Returned for modification 6 February 2008/Accepted 25 April 2008
The measurement of functional measles virus-specific neutralizing antibodies is of considerable interest for vaccine-related research. In this study, we developed and standardized a simple, rapid, highly sensitive, and reproducible fluorescence-based plaque reduction microneutralization (PRMN) assay with visual and auto-mated readout, using a recombinant measles virus engineered to express enhanced green fluorescent protein. The assay is performed in micro format, requires less time to complete (2 versus 4 to 7 days), and is less labor-intensive and less costly than the classical plaque reduction neutralization (PRN) test, widely accepted as the “gold standard” in measles serology. Two available WHO international anti-measles virus standards and one in-house reference serum were used to develop and standardize the new assay. The mean PRMN values from repeated assays were found to be similar to those reported in the literature or assigned to the WHO standards by the classical PRN assay. For validation, we used three groups of low, moderate, and high measles virus vaccine responders’ sera with moderate values of correlation in antibody levels (mIU/ml) between PRMN and the Dade Behring immunoglobulin G enzyme immunoassay (EIA). The PRMN assay was more sensitive at low antibody levels and more informative in terms of protection than this commercial EIA. In conclusion, we have developed and validated a sensitive and high-throughput measles virus-specific PRMN that can be readily used in large population-based measles studies.
Measles is a highly communicable infectious disease, and it remains the leading cause of vaccine-preventable childhood mortality in developing countries and is still a major public health concern in developed countries (15). Measles outbreaks are known to occur even in highly vaccinated populations de-spite the availability of an effective live attenuated measles virus (MV) vaccine (17).
Neutralizing and protective antibodies are directed against the two surface MV glycoproteins, the hemagglutinin (H) and fusion (F) proteins, and are sufficient to provide protection (4). Therefore, diagnostic measures of sufficient levels of functional neutralizing MV-specific antibodies correlate with protection. Neutralizing antibodies are operationally defined by a standard plaque reduction neutralization (PRN) test as antibodies that prevent a cytopathic effect and formation of plaques, using a laboratory MV strain (Edmonston) on Vero cells (4). The PRN assay, an enhanced version of the neutralization test, is widely accepted as the “gold standard” in MV serology and measures the serum dilution capable of preventing 50% of plaque formation by MV (PRN titer; 50% neutralizing dose [ND50]) (1). However, the classical PRN assays are slow (4 to 7 days), labor-intensive, and require large volumes of test com-ponents (since they are conducted in 24-well and 12-well plates using semisolid cell overlay and staining procedures) and are impractical for large numbers of samples. The availability of a
relatively simple, more rapid, reproducible, and sensitive assay for MV neutralizing antibodies, applicable to larger-scale stud-ies, would be of considerable interest for vaccine-related re-search.
For this reason, we have developed and standardized a sim-plified, highly sensitive fluorescence-based plaque reduction microneutralization (PRMN) assay, using recombinant MV based on the Edmonston B strain, engineered to express en-hanced green fluorescent protein, MVeGFP (9, 19). The assay was validated using the second and third World Health Orga-nization (WHO) international anti-MV standards, one in-house reference serum (calibrated against the WHO stan-dard), and three groups of low, moderate, and high MV vaccine responders’ sera with good concordance in antibody titers between PRMN and the Dade Behring immunoglobulin G (IgG) enzyme immunoassay (EIA).
MATERIALS AND METHODS
Study subjects.The study cohort of 346 healthy children and young adults from Olmsted County, MN, has been described previously (16). From this co-hort, we included 30 subjects in our assay validation study based on IgG antibody levels as determined by the Dade Behring anti-MV IgG EIA. We selected the 10 subjects with the highest IgG antibody levels and the 10 subjects with the lowest IgG antibody levels. In addition, we drew a simple random sample of subjects in the middle 50% of the MV IgG antibody levels (that is, those with values falling between the 25th and 75th percentiles) and included them as our moderate IgG antibody response subjects. The majority of the study population was white (93%) and 60% was male, with a mean age of 16 years. The mean ages at the first and second immunization were 25.0 months and 11.7 years, respectively, and the time since last measles-mumps-rubella (MMR-II) immunization was 5.0 years.
MV IgG EIA.Quantitative levels of MV-specific IgG antibody titers for all serum specimens were determined using the Dade Behring anti-MV IgG EIA (Dade Behring, Marburg, Germany; sensitivity, 99.6%; specificity, 100%), ac-cording to the manufacturer’s instructions as described previously (8). The assay
* Corresponding author. Mailing address: Mayo Vaccine Research Group, Program in Translational Immunovirology and Biodefense, Mayo Clinic College of Medicine, 611 C Guggenheim Building, 200 First Street SW, Rochester, MN 55905. Phone: (507) 284-4968. Fax: (507) 266-4716. E-mail: [email protected].
䌤Published ahead of print on 7 May 2008.
1054
on August 17, 2020 by guest
http://cvi.asm.org/
in the middle 50% of MV IgG antibody levels [geometric mean⌬A, 0.595; GMT, 1,746 mIU/ml]) were heat inactivated (56°C; 30 min) and assayed 3 times (or up to 20 times for the standards) on separate days for neutralizing antibodies by PRMN. Serum samples were diluted fourfold from 1:4 to 1:4,096 (four replicates for each dilution) in Opti-MEM I (Gibco, Invitrogen Corporation, Carlsbad, CA), except for the second and the third WHO international anti-MV standards (5 IU, National Institute for Biological Standards and Control [NIBSC] code no. 66/202, and 3 IU, NIBSC code no. 97/648; WHO International Laboratory for Biological Standards, NIBSC, Potters Bar, Hertfordshire, United Kingdom), which were diluted fourfold from 1:16 to 1:16,384. The diluted sera were mixed with equal volumes of low-passage challenge virus MVeGFP (9, 19) (final dilu-tions, 1:8 to 1:8,192 for all sera and 1:32 to 1:32,768 for the WHO standards) and incubated for 1 h at 37°C. A standard inoculum of challenge virus was used in Opti-MEM at a dilution adjusted to yield 15 to 30 PFU per well in the control
wells with virus only. Serum-virus mixtures (50l) were transferred to a new
96-well plate, mixed with an equal volume of Vero cell suspension (1.5⫻
104/well; Vero passage level, 20 to 40 passages), and incubated for 2 h at 37°C,
after which 50l of Dulbecco’s modified Eagle’s medium (Gibco, Invitrogen
Corporation, Carlsbad, CA) containing 10% fetal bovine serum (HyClone,
Lo-gan, UT) was added. The plates were incubated for 48 h at 37°C under 5% CO2,
and the numbers of brightly fluorescent green plaques were counted under UV light using a fluorescence microscope (Nikon Eclipse TE300; Nikon Instruments Inc., Melville, NY). For automatic readout, 48 h after assay setup, the cells were lysed with 4% Igepal CA630 (Sigma Aldrich) in Dulbecco’s modified Eagle’s medium-10% fetal bovine serum (incubation, 1 h at 37°C), and GFP fluorescence was measured using a fluorescence reader (Fluoroscan Ascent; Thermo Electron Corporation, Vantaa, Finland) at 485-nm excitation and 527-nm emission wave-lengths. The PRMN assay controls included the second and third WHO inter-national anti-MV antibody standards, containing 5,000 mIU/ml and 3,000 mIU/ ml, respectively, and one in-house reference serum, calibrated against the second
WHO standard. The PRMN values (ND50) were calculated by Karber’s formula
(2) or using nonlinear regression analysis (Microsoft Excel 2004 version 11.3.7) to calculate the titer that gave a 50% reduction in GFP fluorescence compared
to a virus-only control. PRN values of⬍8 were considered to be negative. PRN
values of⬎120 were thought to indicate protection from symptomatic disease
(5), whereas values of⬎800 (20) or 1,052 (5) indicated sterile immunity and
protection from infection with no antibody boosting after MV exposure. The use of the second WHO international anti-MV antibody standard enabled
quantita-tive ND50values to be transformed into mIU/ml, as described previously (6). A
test limit in terms of mIU/ml was determined for each assay (6). Test sera with reactivities greater than the test limit (corresponding to a PRMN value of 8) were considered PRMN positive.
Statistical methods.Data were descriptively summarized using frequencies and percents for categorical variables and means and ranges for continuous
variables. Plots of antibody levels (in mIU/ml or both ND50and mIU/ml)
sug-gested moderate to high levels of data skewness, so all calculations of summary statistics and all formal statistical analyses for these measures were carried out using log-transformed values. The resulting mean values were then back trans-formed and thus expressed in their original sampling units, effectively yielding GMT values. We determined the reproducibility of the PRMN assay using Shewhart control charts, plotting replicate values of the second WHO standard
and third WHO standard over time. Control limits, depicting the means⫾2
standard deviations (SD), were superimposed on each of the charts. Since EIAs and PRMN (PRN) assays are not expected to give equivalent quantitative values (6, 18), we did not statistically compare the EIA and PRMN values but rather
RESULTS
MV-induced antibody levels, measured by enzyme-linked immunosorbent assay (ELISA). Three hundred forty-six healthy vaccinees were screened for MV-specific antibodies with the Dade Behring EIA kit. The GMT for the MV-specific IgG was 1,390 mIU/ml (range, 28 to 14,999). The proportion of individuals with a positive response for MV-specific antibody was 86% (⌬A⬎0.2), while the remaining 14% were negative or equivocal for MV-specific antibody (⌬A ⱕ 0.2). For the PRMN validation studies, we used three EIA sampling groups, defined by the EIA antibody titer in mIU/ml: 10 sera with the lowest IgG antibody level comprised the low-antibody-level group (EIA GMT, 150 mIU/ml; geometric mean⌬A, 0.096), 10 sera with the highest IgG antibody levels comprised the high-antibody-level group (EIA GMT, 8,610 mIU/ml; geometric mean ⌬A, 1.403), and a random sample of 10 sera in the middle 50% of MV IgG antibody levels comprised the mod-erate-antibody-level group (EIA GMT, 1,746 mIU/ml; geo-metric mean⌬A, 0.595). The group with the lowest IgG anti-body levels included three EIA-negative sera (⌬A⬍0.1) and seven EIA-equivocal sera (⌬Aⱕ0.2).
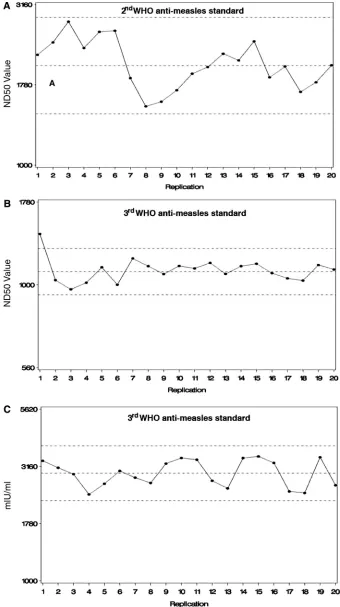
Establishment of PRMN.To establish and characterize the performance of the PRMN assay, we initially used the two available WHO international anti-MV standards with assigned values (second standard 66/202 to 5 IU and third standard 97/648 to 3 IU) and known performance in the classical PRN assay. In addition, we also used a diluted WHO standard (66/ 202) working stock of 200 mIU/ml as described by Ratnam et al. (18) and one in-house high-titer reference serum. All sera were tested against the second WHO anti-MV serum 66/202 as a standard, which allowed us to transform all ND50values into mIU/ml values. After identifying the dynamic range for these products, we conducted multiple PRMN assays using a stan-dard operating procedure over a period of several months. The reproducibility and variability of the PRMN assay were deter-mined by the generation of quality control charts in which the mean⫾2 SD limits for the standards were determined (Fig. 1). The reproducibility of our assay was found to be lower than or comparable to that shown for the classical PRN assay (7) and for other neutralization assays, based on reporter gene expres-sion (14). The geometric mean PRMN titer (ND50) of the second WHO standard 66/202 identified in 20 repeated assays was 2,061 versus a mean ND50value of 2,350, found for the
same standard by the classical PRN assay (7). The perfor-mance of the PRMN assay on low (200-mIU/ml standard by
on August 17, 2020 by guest
FIG. 1. WHO anti-MV standard control charts in PRMN. (A) Second WHO anti-MV standard 66/202 control chart showing PRMN ND50 values in 20 repeated assays. (B and C) Third WHO anti-MV standard 97/648 control charts showing PRMN ND50values (B) and PRMN mIU/ml values (C) in 20 repeated assays. The values are plotted on a log scale. The dashed horizontal lines depict the sample mean⫾2 SD limits.
on August 17, 2020 by guest
http://cvi.asm.org/
and high (3,000-mIU/ml third WHO standard by PRN)-antibody-level standards was also examined. The geometric mean PRMN titer of the third WHO standard 97/648 for 20 repeated tests was 1,107, and the geometric mean calculated value was 2,948 mIU/ml (the assigned value for this standard is 3,000 mIU/ml as determined by the classical PRN assay). The second WHO diluted working stock of 200 mIU/ml was tested 10 times, and the geometric mean PRMN titer (ND50) was 82
versus a PRN titer of 94, found for the same standard by Ratnam et al. (18), while the geometric mean calculated value in mIU/ml was 211. In a set of repeated assays, we were also able to assess the performance of our in-house reference se-rum using our PRMN assay with respect to its future use in this assay as a standard. The geometric mean PRMN titer (ND50)
of this serum was 1,207, and the calculated GMT value was 2,999 mIU/ml. Thus, the in-house reference serum was as-signed a value of 3,000 mIU/ml for future studies. In 10 re-peated assays of the third WHO standard 97/648, we compared the visual and the automatic readouts. The geometric mean ND50value based on visual plaque counting was 927, while the
geometric mean ND50 value calculated from the measured GFP fluorescence was 980, and there was no significant differ-ence between the mean values from visual and automatic read-outs (P⫽0.45;ttest on log-transformed values).
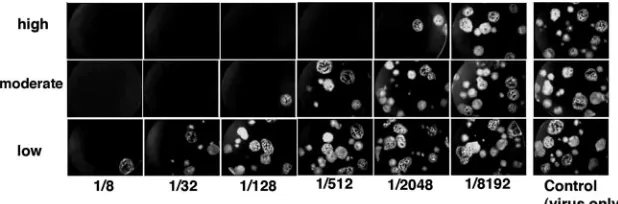
Correlation between PRMN and Dade Behring IgG EIA in MV immunity testing.To further characterize the new PRMN assay, we explored its performance on human samples using low, moderate, and high MV vaccine responders’ sera. GFP expression and plaque visualization only 48 h after assay setup are shown in Fig. 2, which also depicts the performances of representative low-, moderate-, and high-titer sera in PRMN. Summary Dade Behring EIA results and PRMN assay results for the three groups are presented in Table 1. The EIA GMT for the low-antibody-level group was 150 mIU/ml (range, 28 to 249) versus 104 mIU/ml (range, 21 to 257) in PRMN. The GMT in EIA for the moderate-antibody-level group was 1,746
mIU/ml (range, 1,196 to 2,633) versus 1,086 mIU/ml (range, 263 to 2,732) using the PRMN assay, and the GMT in EIA for the high-antibody-level group was 8,610 mIU/ml (range, 6,481 to 14,999) versus 3,319 mIU/ml (range, 876 to 10,246) using the PRMN assay (Table 1). All of the samples from the low-antibody-level group were found to be negative (three sera) or equivocal (seven sera) using the EIA, while the same sera were positive using the PRMN assay, although the neutralizing an-tibodies were well below the protective level (PRN titer ⬎ 120). The rest of the sera were positive in both assays.
The associations between MV neutralizing-antibody levels and EIA levels are visually displayed in Fig. 3. Most values fall to the right of the 45° line of equivalence, indicating that on average EIA values were higher than neutralizing-antibody values. Group-specific and overall correlations between the EIA and PRMN antibody titers were moderate to high (Fig. 3). Intraclass correlations ranged between 0.58 (subset to the group with moderate EIA values) to 0.68 (in the group with low EIA values). Correlations tended to be greater in the low and high EIA groups than in the moderate group, perhaps reflecting the decreased variability in the extreme groups due to the sampling scheme. Group-specific Pearson correlations, which are more sensitive to the sampling design, ranged from 0.21 in the moderate EIA group to 0.82 in the high group. The distribution of PRMN titers in terms of protection (Fig. 4) revealed that this specific broad-range set of sera represented all three main categories relevant to MV protective antibody levels: below the level of protection, protection from symptom-atic disease, and protection from infection.
DISCUSSION
A battery of immunological assays has been used to evaluate MV humoral immunity, among which the classical PRN assay correlates best with protection in vivo (4, 5, 20). In this study, we developed and standardized a practical alternative to the
TABLE 1. Antibody titers and 50% neutralizing doses of test sera by EIA and PRMN
MV antibody
response category No. of observations per category
ELISA GMT (mIU/ml)
(rangea
)
PRMN GMT (mIU/ml) (range)
PRMN GMT (ND50)
(range)
Low 10 for ELISA, 30 for PRMN 150 (28–249) 104 (21–257) 50 (12–130)
Moderate 10 for ELISA, 30 for PRMN 1,746 (1,196–2,633) 1,086 (263–2,732) 433 (127–1,205) High 10 for ELISA, 30 for PRMN 8,610 (6,481–14,999) 3,319 (876–10,246) 1,702 (490–5,328)
a
The range is defined as the minimum and maximum values found for each assay in each of the respective sampling groups.
on August 17, 2020 by guest
classical PRN—a rapid, simple, and reproducible PRMN test based on recombinant MV expressing GFP.
The performance of the two WHO international anti-MV standards in PRMN gave us the opportunity to develop and validate the test, to transform all values into mIU/ml, and to compare our results with those of the classical PRN assay, since the WHO standards’ values were established and de-signed for use mainly in the PRN assay. The PRMN assay requires substantially less incubation time (2 days versus 4 to 7 days) and fewer test components (no overlay medium and no staining and fixation reagents) than the classical PRN assay and eliminates the complicated steps of semisolid cell overlays, fixation, and staining. Moreover, the GFP expression greatly facilitates and improves the test readout. We have demon-strated that the GFP-based PRMN assay is rapid, reproduc-ible, and sensitive and produces results similar to those ob-tained by the classical PRN assay. The PRMN assay values from 20 repeated assays were remarkably similar to those re-ported in the literature (7, 18) or assigned to the WHO stan-dards by the classical PRN assay. The new assay is basically a PRN assay with visual (plaque enumeration) and/or automatic
readout, which renders results consistent with those in the classical PRN assay. This is of utmost importance, since the reported PRN antibody levels relevant to protection could be directly applied to the interpretation of PRMN assay results. However, an important limitation of a PRMN assay with visual readout is human subjectivity. While the fluorescence reader-based assay offers clear advantages in time and human re-sources, it does require specific instrumentation. The potential sources of error with the latter method, such as high back-ground fluorescence, edge effects, and sample fluorescence/ quenching, should be addressed prior to implementation.
Several EIA kits for anti-MV antibodies are available com-mercially, among which the Dade Behring EIA kit was previ-ously found to perform better in comparison with other com-mercial EIA kits based on a PRN titer of 120 as a cutoff (12, 18). While these kits are easy to use, they are generally not sufficiently sensitive to detect low levels of antibodies and are therefore not optimal for seroprevalence surveys (11). To val-idate our new assay, we used low, moderate, and high MV vaccine responders’ sera as determined by the Dade Behring EIA. The new assay was sensitive enough to detect both low-titer and high-low-titer neutralizing antibodies in clinical serum samples. Similar to previous reports for PRN (6, 11), we found that the PRMN assay showed much better sensitivity than the Dade Behring EIA kits and was able to detect low levels of anti-MV antibodies in samples that gave negative/equivocal results using the EIA. Analyzing the associations between MV PRMN antibody levels in mIU/ml and EIA antibody levels, we found moderate values of correlation, and on average, EIA values were higher than neutralizing antibody values. Others have also reported similar results for the classical PRN assay and the Dade Behring EIA (6). The discrepancies between the enzyme immunoassays and PRMN (PRN) assays may be largely due to the differing formats and MV antigens used in the two tests. Most of the commercial enzyme immunoassays and the Dade Behring EIA kit, in particular, detect antibodies
FIG. 3. Association between MV neutralizing-antibody and EIA levels. (A) Values are plotted on a log scale. Each individual is rep-resented three times, once per measure of neutralizing antibodies. The dashed lines separate the three sampling strata as defined by EIA values. The solid line is the 45° line of equivalence. Most values fall to the right of the line, indicating that on average EIA values are larger than neutralizing-antibody values. (B) Correlations between EIA and neutralizing-antibody response measures, overall and by EIA-defined subgroups. Interaction correlations are for three neutralizing-antibody measures per individual. Pearson correlations are based on one obser-vation per individual, using the mean of the three neutralizing-anti-body measures. Overall results for the Pearson correlation should be interpreted with caution, as they are more heavily affected by the sampling design than the intraclass correlations.
FIG. 4. Relevance of PRMN ND50values to protective antibody levels. PRMN values of low-, moderate-, and high-antibody-titer sera (30 observations per category) were plotted on a log scale, and the antibody levels relevant to protection are shown as dotted lines: 120 (5), threshold for protection against symptomatic disease; 800 (20) or 1,052 (5), threshold for protection against infection and antibody boosting after MV exposure.
on August 17, 2020 by guest
http://cvi.asm.org/
izing antibodies (14). For MV, alternatives to the classical PRN assay include H protein-based EIA (3), neutralization EIA, (13) and a neutralization test, based on a recombinant GFP-expressing MV (10). The last study applied a different strain of MV (AIK-C) and a longer and more complicated assay protocol, including a semisolid agarose overlay. This as-say did not apply the WHO international anti-MV standards for validation and transformation of the values into mIU/ml, and therefore, it is difficult to compare the two studies or correlate the neutralization values to protection.
In conclusion, this new reporter gene-based PRMN assay provides an expedited and high-throughput alternative to the classical PRN assay. With its delineated protective measures, the validated PRMN assay can be readily transferred to other laboratories and applied to MV immunity testing in larger population-based studies.
ACKNOWLEDGMENTS
This work was partially supported by NIH grants AI33144 and AI48793.
We thank Roberto Cattaneo (Mayo Clinic, Rochester, MN) for providing the GFP-expressing MV. We thank Neelam Dhiman and Rick Kennedy from the Mayo Vaccine Research Group for helpful suggestions and technical help. We also thank Cheryl Hart for techni-cal assistance in preparing the manuscript.
Gregory A. Poland is the chair of a DMSB for non-MV-based vaccines in development by Merck Research Laboratories.
REFERENCES
1.Albrecht, P., K. Herrmann, and G. R. Burns.1981. Role of virus strain in conventional and enhanced measles plaque neutralization test. J. Virol.
Methods3:251–260.
2.Ballew, H. C.1992. Neutralization, p. 229–241.InS. Specter and G. Lancz (ed.), Clinical virology manual. Elsevier, New York, NY.
3.Bouche, F., W. Ammerlaan, F. Berthet, S. Houard, F. Schneider, and C. P.
9.Duprex, W. P., S. McQuaid, L. Hangartner, M. A. Billeter, and B. K. Rima.
1999. Observation of measles virus cell-to-cell spread in astrocytoma cells by using a green fluorescent protein-expressing recombinant virus. J. Virol.
73:9568–9575.
10.Fujino, M., N. Yoshida, K. Kimura, J. Zhou, Y. Motegi, K. Komase, and T. Nakayama.2007. Development of a new neutralization test for measles virus.
J. Virol. Methods142:15–20.
11.Goncalves, G., F. Cutts, T. Forsey, and H. R. Andrade.1999. Comparison of a commercial enzyme immunoassay with plaque reduction neutralization for maternal and infant measles antibody measurement. Rev. Inst. Med. Trop.
Sao Paulo41:21–26.
12.Hesketh, L., A. Charlett, P. Farrington, E. Miller, T. Forsey, and P. Morgan-Capner.1997. An evaluation of nine commercial EIA kits for the detection
of measles specific IgG. J. Virol. Methods66:51–59.
13.Lee, M. S., B. Cohen, J. Hand, and D. J. Nokes.1999. A simplified and standardized neutralization enzyme immunoassay for the quantification of
measles neutralizing antibody. J. Virol. Methods78:209–217.
14.Manischewitz, J., L. R. King, N. A. Bleckwenn, J. Shiloach, R. Taffs, M. Merchlinsky, N. Eller, M. G. Mokplajczyk, D. J. Clanton, T. Monath, R. A. Weltzin, D. E. Scott, and H. Golding.2003. Development of a novel
vaccinia-neutralization assay based on reporter-gene expression. J. Infect. Dis.188:
440–448.
15.Moss, W. J., and D. E. Griffin.2006. Global measles elimination. Nat. Rev.
Microbiol.4:900–908.
16.Ovsyannikova, I. G., R. M. Jacobson, R. A. Vierkant, S. J. Jacobsen, V. S. Pankratz, and G. A. Poland.2004. The contribution of HLA class I antigens in immune status following two doses of rubella vaccination. Hum. Immunol.
65:1506–1515.
17.Poland, G. A., and R. M. Jacobson.1994. Failure to reach the goal of measles elimination. Apparent paradox of measles infections in immunized persons.
Arch. Intern. Med.154:1815–1820.
18.Ratnam, S., V. Gadag, R. West, J. Burris, E. Oates, F. Stead, and N. Boulianne.1995. Comparison of commercial enzyme immunoassay kits with plaque reduction neutralization test for detection of measles virus antibody.
J. Clin. Microbiol.33:811–815.
19.Vongpunsawad, S., N. Oezgun, W. Braun, and R. Cattaneo.2004. Selectively receptor-blind measles viruses: identification of residues necessary for SLAM- or CD46-induced fusion and their localization on a new
hemagglu-tinin structural model. J. Virol.78:302–313.
20.Ward, B. J., S. Aouchiche, N. Martel, F. M. Bertley, N. Bautista-Lopez, B. Serhir, and S. Ratnam.1999. Measurement of measles virus-specific neu-tralizing antibodies: evaluation of the syncytium inhibition assay in compar-ison with the plaque reduction neutralization test. Diagn. Microbiol. Infect.
Dis.33:147–152.