Eiszeitalter u. Gegenwart Band 21 Seite 97-107 Öhringen/Württ.,31. Dezember 1970
Versuch einer n e u e n P l e i s t o z ä n - G l i e d e r u n g
Von J U L I U S H E S E M A N N , K r e f e l dMit 4 Tabellen
Z u s a m m e n f a s s u n g . Die Fülle altbekannter und neuerdings gefundener Bildungen so wie physikalischer Messungen des Pleistozäns legt ein intergredierendes Gliederungsschema zur wahlweisen Einordnung der Fakten nahe. Es basiert auf einer durch die Kalt- und Warmzeiten repräsentierten Zyklik und ist auf Mitteleuropa bezogen. Die Einwirkung von mindestens 25 Zyklen auf die Tier- und Pflanzenwelt wird diskutiert.
S u m m a r y . The abundance of old an new sediments and also physical measurements of Pleistocene suggest an integrating schema of organization to classify the facts by choice. This schema is based on a cycle reprented by the cold and warm periods and refers to Mid-Europe. The influence of at least twenty-five cycles on fauna and flora is discussed.
D i e Gliederung des Pleistozäns erscheint in den derzeitigen Aufstellungen unbefriedi gend. Unausgefüllt bleibt vor allem die Lücke zwischen der ältesten K a l t z e i t und dem P l i o z ä n , deren Schließung durch ungewöhnliche Verlängerungen voraufgegangener K a l t zeiten nicht überzeugen kann. Aber auch das klassische, inzwischen a u f sechs (Weichsel-, S a a l e - , Elster-, M e n a p - , Eburon-, B r ü g g e n - ) K a l t - und Warmzeiten erweiterte S y s t e m w i r d durch die Auffindung zwischenzeitlicher B o d e n - und Vegetationsprofile, neuer T e r rassen und glaziärer Sequenzen fraglich. D a s Unbehagen wird durch die Aufteilung v o n K a l t - u n d Warmzeiten auf G r u n d v o n Wärmeschwankungen von ungewisser D a u e r ( G r o ßes Interglazial und Interglazial, Großes Interstadial und Interstadial, Schwankung u n d H o r i z o n t usw.) nicht beseitigt, sondern nur verlängert. Dabei ist der Z u g v o m O l i g o - z u m P o l y g l a z i a l i s m u s unverkennbar, nur vermißt man eine einleuchtende und die neuen Beobachtungen „intergrierende" Pleistozänstratigraphie.
In bisher noch nicht gekannter Vielseitigkeit stehen n e u e F a k t e n zur V e r f ü g u n g : g l a z i ä r e Sequenzen, B ö d e n und D i s k o r d a n z e n , Faunen und Floren, Geschiebe, G e r ö l l -und Schwermineral-Gesellschaften, physikalische Altersdaten -und Paläotemperaturen. Ihre Auswertung und Zusammenfügung w i r d beeinträchtigt durch
1. die fehlende Unterscheidung großer und kleiner Wärmeschwankungen (um eine Potenz, entweder wenige 100 bis 1000 oder ± 10 000 J a h r e ) ,
2. die Ignorierung jener K a l t - und Warmzeiten, welche keine sichtbaren Ä q u i v a l e n t e in Moränen, Terrassen und Hochstränden hinterlassen, weil sie in der Ausdehnung, Eintiefung oder in Hochstränden hinter ihren Vorläufern zurückblieben und sich (wie z. B . in über 40 m hohen Terrassenstufen) nicht abheben,
3. die Unvereinbarkeit der A n n a h m e einer Repräsentation des Pleistozäns durch lediglich etwa 10 Eiszeiten (unter Berücksichtigung von Weichsel I und I I , S a a l e I und I I usw.) mit der viel größeren Z a h l v o n Terrassen und Zeitabschnitten o z e a n i scher Minimal- und M a x i m a l - T e m p e r a t u r e n ,
4 . das Fehlen eines zeitlichen, übergeordneten Maßstabes in F o r m einer Periodizität, welche die Wärmeschwankungen reguliert und die A k k u m u l a t i o n wie die E r o s i o n in einen immer wiederkehrenden A b l a u f zwingt, den F a u n a und F l o r a ebenfalls widerspiegeln ( Z y k l i k ) .
Ich halte einen Versuch, alle bekannten Alters- und Paläotemperatur-Messungen sowie alle bekannten pleistozänen Bildungen einschließlich der F a u n a und F l o r a miteinander in Beziehung zu setzen und in ein zeitliches Schema einzuordnen, trotz aller Lückenhaftigkeit als Zwischenergebnis schon jetzt für k l ä r e n d und anregend. D a s pleistozäne Geschehen
entspricht zweifellos einer P e r i o d i k . Sie tritt uns in verschiedener Ausbildung ent gegen: in der Terrassenbildung (Schotter-Hochflutlehm-Hochflutlehm-Schotter), in den Bodenprofilen ( T u n d r a g l e y - P a r a b r a u n e r d e - B r a u n e r d e - T u n d r a g l e y ) , in der Waldentwick lung (Kiefern/Birken - Zeit - Eichenmischwald - Birken/Kiefern - Zeit), in der Faunenfolge (Lößschnecken/arktische WirbeltiereSteppen u n d WaldtiereLößfauna) oder in den g l a -ziären Sequenzen (Löß-Vorschüttsande-Moräne-Nachschüttsande-Löß).
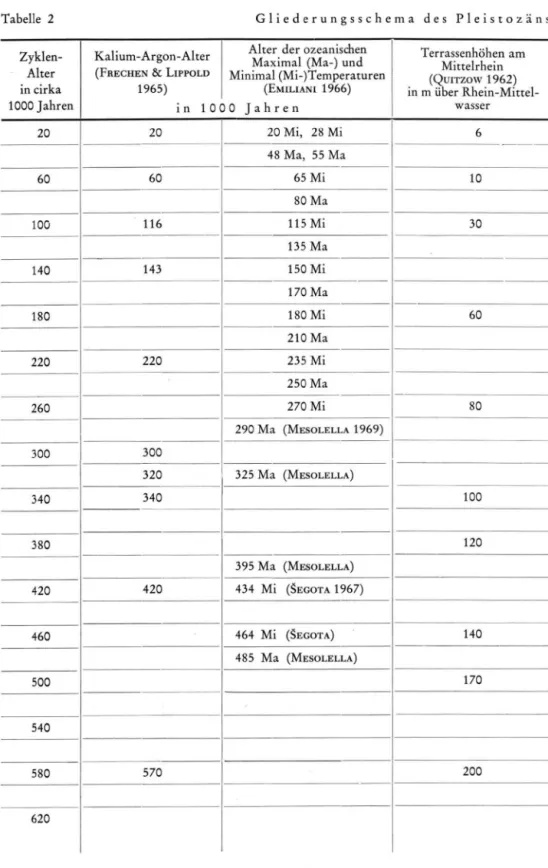
Sucht m a n die Periodik als Phasenfolge zu klassifizieren, so entspricht sie einer Z y k l i k oder Cyclothemen. N i m m t m a n als eine der Voraussetzungen z u r Bildung v o n Sedimen tations-Zyklen eustatische Meeresspiegelschwankungen an, wie sie dem Gletscherhaushalt, Terrassen u n d Hochstränden entsprechen, dann hat m a n den M a ß s t a b für den A b l a u f u n d die D a u e r der pleistozänen K a l t - und Warmzeiten gefunden (Tabellen 2 und 3 ) . Setzt m a n die Cyclotheme in Beziehung zu den ozeanischen Wärmeschwankungen ( E M I L I A N I 1966, M E S O L E L L A U. a. 1969, für Zentraleuropa S E G O T A 1967), z u den Ergebnissen der K / A r M e t h o d e ( F R E C H E N & L I P P O L T 1965) u n d zu den Berechnungen ( T a b . 1) v o n S O E R -GEL (1937) u n d Z E U N E R ( 1 9 3 9 ) , so ergibt sich, soweit überhaupt Bestimmungen v o r genommen wurden, eine gute Übereinstimmung. Differenzen erklären sich aus d e m Bil dungszeitraum v o n Terrassen u n d glaziären Bildungen. Manche Terrassen waren für Z E U N E R u n d S O E R G E L nicht erkennbar, so daß die Zuordnung der beobachteten Terrassen oder Kaltzeiten zu den Eiszeiten eine zu frühe Datierung ergeben mußte ( T a b . 1 ) . D i e N a m e n s g e b u n g ist jedoch eine Sache der K o n v e n t i o n und hat m i t der Einordnung der K a l t - u n d Warmzeiten in den jeweiligen Z y k l u s nichts zu tun. Wichtig ist aber die D a u e r eines Z y k l u s v o n rund 40 000 J a h r e n , eine Zahl, welche sich aus den Altersmessungen er gibt und mit der D a u e r der Cyclotheme des flözführenden Oberkarbons ( J E S S E N 1961) übereinstimmt.
Tabelle 1
D a s A l t e r d e r K a l t z e i t e n n a c h v e r s c h i e d e n e n M e t h o d e n u n d i h r e B e n e n n u n g n a c h S O E R G E L ( 1 9 3 7 ) .
Alter in 1 0 0 0 Jahren nach
Zyklität Kalium/ Argon-Methode FRECHEN SC LIPPOLD 1 9 6 5 Ozeanische Minimal temperaturen (EMILIANI 1 9 6 6 ) ZEUNER 1 9 3 9 SOERGEL 1 9 3 7 Eiszeit (SOERGEL 1 9 3 7 ) 2 0 2 0 2 0 2 5 2 0 Würm 3 6 0 6 0 6 0 7 2 7 0 Würm 2 1 0 0 1 1 6 1 1 0 1 1 5 1 1 0 Würm 1 (Warthe) 140 1 4 0 1 5 0 1 4 5 1 4 0 Riß 3 180
—
—
1 8 7 1 8 5 Riß 2 2 2 0 2 2 0 2 3 5 2 3 0 2 3 0 Riß 1 2 6 0—
2 7 0—
— — 3 0 0 3 0 0 — 3 0 5 3 0 5 Präriß 3 4 0 3 4 0—
—
—
—
3 8 0—
3 9 5 3 9 5 Mindel 3 4 2 0 4 2 0 4 3 4 (SEGOTA 1 9 6 7 ) 4 3 5 4 3 0 Mindel 2 4 6 0 5 0 0 540 — 4 6 4 (SEGOTA) 4 7 6 4 7 5 Mindel 1 4 6 0 5 0 0 540—
5 5 0 5 4 5 Günz 2 5 8 0 5 7 0 — 5 9 0 5 9 0 Günz 1Versuch einer neuen Pleistozän-GIiederung 99
E s h a t aber, wie z . B . in der Weichsel-Eiszeit, etliche dicht aufeinander folgende W ä r meschwankungen gegeben, welche nicht durch einen Temperaturanstieg wie bei Inter glazialen, sondern durch ihre Häufung u n d G e s a m t d a u e r eine (wenn auch in unseren Brei ten abgeschwächte Z y k l i k und) Teilung einer Eiszeit z u m Ausdruck bringen.
T ü r die rückwärtige Verfolgung der K a l t z e i t e n sind hier drei Kriterien benutzt w o r den: Rhein-Terrassen, K / A r - D a t e n u n d d a s Villafranc mit seinen Lößprofilen ( B R U N N A C K E R 1967). Terrassen lassen sich a m Mittelrhein ( Q U I T Z O W 1962) u n d a n der R u h r bis zu den Oberen H ö h e n (Homberg-, M e t t m a n n ) - T e r r a s s e n x) verfolgen ( T a b . 3 ) . Stellt m a n
die teilweise (bis 60 m ) großen A b s t ä n d e in Rechnung, lassen sich diese Terrassen nach Z y k l e n bis zum Abschnitt der Eintiefung der Terrassentreppen in die vorhergegangene T r o g r e g i o n etwa u m die Zeit von 0,8 M i o J a h r e n aufteilen. Veranschlagt m a n für die Zeit der Eintiefung der Trogregion zum H o c h t a l um 50 m die Dauer v o n vier Cyclothemen, so gelangt man bis z u r Zeitmarke 0,94 M i o J a h r e u n d damit nahe an das älteste p h y s i k a lisch datierte G l a z i a l (in Kalifornien) v o r 0,98 Mio J a h r e n ( T a b . 3).
A u f diese Weise ergibt sich ein mindestens 25 Eiszeiten umfassender P o l y g l a z i a l i s m u s . Es bleibt nur übrig, diese theoretisch gefolgerten K a l t z e i t e n auch durch Sedimente zu b e legen. Einen Weg d a z u h a t B R U N N A C K E R mit seinen Schülern durch die planmäßige B e s t a n d s a u f n a h m e weit zurückreichender Quartärprofile am Mittelrhein mit Kärlich
( B R U N N A C K E R , S T R E I T & S C H I R M E R 1 9 6 9 ) und a m Niederrhein mit B r ü g g e n ( B O E N I G K
1969) und Frechen ( K O W A L Z I K 1970) beschritten. D a b e i sind mehrere L ö ß und T e r a s -sensysteme oberhalb der Hauptterrasse aufgefunden worden, welche die große Lücke z w i schen Villafranc u n d Tegelen ausfüllen können u n d über ein einziges „ P r ä t i g l i u m " hinaus gehen dürften. Versucht man das Villafranc v o n S t . Vallier als B a s i s d e s P l e i s t o z ä n s mit seinen drei Bodenprofilen u n d dem liegenden Periglazialschotter v o n C h a m -b a r a n a n die älteren Rheinterrassen anzuschließen, gelangt man für das hier fixierte älteste G l a z i a l ebenfalls z u einem Alter von 0,9 Mio J a h r e n . Eine D i s k o r d a n z trennt das V i l l a franc v o n St. Vallier v o n ariden Liegendschottern, welche wohl ohne Bedenken dem P l i o z ä n zugeordnet werden können (Tab. 3 ) . D i e Basis des Villafranc ist altersmäßig mit rund 1,6 M i o Jahren bestimmt. E s bleibt also eine Lücke v o n 0,6 Mio Jahren v o n der Basis des Villafranc bis zum ersten belegten G l a z i a l . Ein solcher Zeitraum dürfte für die Ablösung der pliopleistozänen Wirbeltier-Mischfauna v o n Beginn bis Ende des Villafranc nötig sein. Die ersten Terrassentreppen (Höhenterrassen) bedeuten nicht nur morphogenetisch eine Wende, sondern dürften auch unschwer eine Tieferlegung der Erosionsbasis durch das jetzt auf Norddeutschland übergreifende Inlandeis anzeigen. Fortan gab es enge Wechselwir kungen zwischen Terrassen-Einschneidung u n d L a g e der Nordseeküste, zwischen zeitwei liger Blockierung der N o r d s e e und Auf schotterung.
Die T i e r w e l t reagierte auf den mindestens 25maligen Wechsel v o n Warm- und K a l t z e i t e n schließlich ( v o n der Gegenwart her betrachtet) mit dem Verschwinden w ä r m e liebender Elemente (Elefant, Nashorn, A n t i l o p e , Flußpferd, Wasserbüffel, L ö w e , Panther, H y ä n e usw.). Lediglich die Vögel kehrten d a n k ihrer größeren Beweglichkeit jahreszeitlich als Z u g v ö g e l zurück. F ü r das Verhalten der übrigen Fauna leuchtet die Vorstellung ( K R E T Z O I 1961 nach T O E P F E R 1963) ein, d a ß in günstigen Zeiten neue Formen in Wellen oder „Fronten" in Mitteleuropa einwanderten, sich in den Warm- (mediterrane Steppen u n d Wälder) und K a l t z e i t e n (Lößsteppen u n d Tundra-Gebiete) mit verschiedenem E r f o l g (bei der Nichtumkehrbarkeit des Evolutionsgeschehens) anpaßten u n d in Warmzeiten durch neue Faunenwellen verdrängt w u r d e n . Die einzelnen Zyklen waren indessen für die Setzung umfassender Zäsuren in der V e r ä n d e r u n g der Tierwelt zu k u r z . Abgesehen v o n
1) Die Zuordnung der Höhen (Mettmann-, Hösel-, Drüfel)-Terrassen zum Pliozän, der ich noch kürzlich (HESEMANN 1969) gefolgt war, ist demnach durch die Verweisung ins Altpleistozän zu berichtigen.
Tabelle 2 G l i e d e r u n g s s c h e m a d e s P l e i s t o z ä n s Zyklen-Alter in cirka 1 0 0 0 Jahren Kalium-Argon-Alter
(FRECHEN & LIPPOLD 1 9 6 5 )
i n 1 0
Alter der ozeanischen Maximal (Ma-) und Minimal (Mi-)Temperaturen (EMILIANI 1 9 6 6 ) 3 0 J a h r e n Terrassenhöhen am Mittelrhein (QUITZOW 1 9 6 2 ) in m über Rhein-Mittel wasser 2 0 2 0 2 0 Mi, 2 8 Mi 6 4 8 Ma, 5 5 Ma 6 0 6 0 6 5 Mi 1 0 8 0 Ma 1 0 0 1 1 6 1 1 5 Mi 3 0 1 3 5 Ma 1 4 0 1 4 3 1 5 0 Mi 1 7 0 Ma 1 8 0 1 8 0 Mi 6 0 2 1 0 Ma 2 2 0 2 2 0 2 3 5 Mi 2 5 0 Ma 2 6 0 2 7 0 Mi 8 0 2 9 0 Ma (MESOLELLA 1 9 6 9 ) 3 0 0 3 C 0 3 2 0 3 2 5 Ma (MESOLELLA) 3 4 0 3 4 0 1 0 0 3 8 0 1 2 0 3 9 5 Ma (MESOLELLA) 4 2 0 4 2 0 4 3 4 Mi (SEGOTA 1 9 6 7 ) 4 6 0 4 6 4 Mi (SEGOTA) 1 4 0 4 8 5 Ma (MESOLELLA) 5 0 0 1 7 0 5 4 0 5 8 0 5 7 0 2 0 0 6 2 0
Versuch einer neuen Pleistozän-Gliederung 101
s e i t 0 , 6 M i o J a h r e n n a c h Z y k l e n
Rheinterrassen, () vermutlich in der gleichen Terrassenstufe
enthalten Kalt- u. Warmzeiten, () theoretisch Mögliche Einordnung (Eu) Homininen ( H E B E L E R & KURTH 1966)
Untere Niederterrasse Kaltzeit 1 Weichsel 2 Homo sapiens sapiens Warmzeit P o p e r i n g e - T o r f e Moersrioofd /
Obere Niederterrasse Kaltzeit 2 Weichsel 1 H. sapiens neanderthalensis Warmzeit Eem H. sapiens praeneanderthalensis Unt.Mittelterr.,Krefeld,Mittelterr. Kaltzeit 3 Warthe
Warmzeit Ohe ? Untere Mittelterr. (Talweg-Terr.) Kaltzeit 4 Drenthe
Warmzeit Holstein H. sapiens praesapiens Mittl. Mittelterr. (Rinnenschotter) Kaltzeit 5
Homo erectus heidelbergensis Warmzeit
Homo erectus heidelbergensis (Oberer Mittl. Mittelterrasse) Kaltzeit 6
Homo erectus heidelbergensis Warmzeit Homo erectus heidelbergensis Obere Mittelterrasse Kaltzeit 7 Elster
Homo erectus heidelbergensis
Warmzeit
Homo erectus heidelbergensis
Oberste Mittelterrasse Kaltzeit 8
Homo erectus Warmzeit
Homo erectus Untere jung. Hauptterrasse Kaltzeit 9
Homo erectus Warmzeit Cromer 2 Homo erectus Obere jung. Hauptterrasse Kaltzeit 10
Homo erectus
Warmzeit Cromer 1
Homo erectus
(Ob. jung. Hauptterrasse) Kaltzeit 11 Menap Warmzeit Waal Mittlere Hauptterrasse Kaltzeit 12 Eburon
Warmzeit Tegelen 2 Obere Hauptterrasse Kaltzeit 13
Warmzeit Tegelen 1 (Obere Hauptterrasse) Kaltzeit 14 Brüggen
Warmzeit Untere (Drüfel) Höhenterrasse Kaltzeit 15
Warmzeit Untere (Hösel) Höhenterrasse Kaltzeit 16
Tabelle 3 G l i e d e r u n g s s c h e m a d e s P l e i s t o z ä n s v o m P l i o z ä n b i s v o r 0 , 6 M i o J a h r e n n a c h Z y k l e n Zyklen-Alter in ca. 1 0 0 0 Jahren Kalt- und Warmzeiten
Mögliche Zuordnung von Terrassen am Mittelrhein
(QUITZOW 1 9 6 2 )
Villafranc-Profil
(BRUNNACKER 1 9 6 7 )
Einschnitte in Morphologie, Fauna und Flora
Warmzeit
6 6 0 Kaltzeit 1 7 (Obere Höhenterrasse 1 ) ( 2 6 0 m üb. Rhein-Mittelwass.)
Warmzeit
7 0 0 Kaltzeit 1 8 Obere Höhenterrasse ( 2 ) (Homberg-, Mettmann-T.) 0,7 Mio Jahre: Erscheinen insta
biler Schwerminerale Warmzeit
7 4 0 Kaltzeit 1 9 (Obere Höhenterrasse 3 )
Warmzeit
7 8 0 Kaltzeit 2 0 (Obere Höhenterrasse 4 )
Warmzeit
0,8 — 1 Mio Jahre: Eintiefung
8 2 0 Kaltzeit 2 1 Villafranc: Löß 3 der Trogregion am Mittelrhein um 5 0 m zum Hochtal
( 3 1 0 — 2 6 0 m über Rhein-Mittelwasser)
Bodenprofil
der Trogregion am Mittel rhein um 5 0 m zum Hochtal ( 3 1 0 — 2 6 0 m über Rhein-Mittelwasser) 8 6 0 Kaltzeit 2 2 Villafranc: Löß 2 Bodenprofil 9 0 0 Kaltzeit 2 3 Villafranc: Löß 1 1 0 m mächtiges Bodenprofil
9 4 0 Kaltzeit 2 4 Villafranc: Periglaziale Schotter von Chambaran Hipparion, Antilopen und die
meisten altweltlichen Affen ver schwunden (DIETRICH 1 9 5 3 ) Tertiäre Floren-Elemente unter 1 0 %
9 8 0 Älteste datierte
Kaltzeit D i s k o r d a n z
Hipparion, Antilopen und die meisten altweltlichen Affen ver schwunden (DIETRICH 1 9 5 3 ) Tertiäre Floren-Elemente unter 1 0 %
1 4 0 0 Villafranc „Neue Wirbeltier-Gattungen in
altertümlichen Assoziationen" (KAHLKE 1 9 6 8 ) Australopithecus (älteste humane Hominiden) 1 6 0 0 Früh-Villafranc „Neue Wirbeltier-Gattungen in altertümlichen Assoziationen" (KAHLKE 1 9 6 8 ) Australopithecus (älteste humane Hominiden) Pliozän Liegendschotter ariden Charak
ters mit roten Einlagerungen
Tertiäre Floren-Elemente bis 3 2 %>
Versuch einer neuen Pleistozän-Gliederung 103
Dauerformen ( z . B . Dicerorhinus etruscus, D. kirchbergensis) lösen (besonders unter den Kleinsäugern, T O E P F E R 1 9 6 3 ) einzelne G a t t u n g e n einander ab oder (bei den G r o ß s ä u g e r n ) Stammreihen entwickeln sich. D i e Lebensdauer v o n Arten u n d Gattungen liegt dabei viel fach zwischen 0 , 1 — 0 , 3 M i o J a h r e n .
M a n hat deshalb die Glieder der E l e f a n t e n - oder Pferde-Stammreihen als M a ß s t ä b e für die P l e i s t o z ä n - G l i e d e r u n g genommen u n d so Alt-, (vorher auch noch Ä l t e s t ) - , Mittel-und J u n g p l e i s t o z ä n unterschieden, ohne a b e r die Ungleichheit der Zeitabschnitte zu beto nen. V o n den (im Verhältnis zur theoretischen Vielzahl) überlieferten wenigen Faunen zeichnen sich jedoch einige dadurch aus, d a ß besonders viele alte Formen erloschen sind und besonders viele neue F o r m e n erscheinen. Sechsmal fällt ein solcher Wechsel a u f :
1. A m E n d e des Früh-Villafranc, vor e t w a 1,4 Mio J a h r e n . Nicht mehr v o r h a n d e n : Zyglodon borsoni, Ursus etruscus,
neu: Desmana vehringi, Mimomys pliocaenicus, Sus arvernensis, Leptobos elatus, Hystrix refossa.
2. A m E n d e des V i l l a f r a n c , vor 1 M i o J a h r e n . Nicht mehr v o r h a n d e n : Soriculus, Prospalax,
neu: Equus stenonis, Allohippus robustus, Dicerorhinus etruscus, Die. kirchbergen-sis, Archidiskon meridionalis, Elepbas planifrons, Crocuta perrieri.
3. A m E n d e der Tegelen-Warmzeit, v o r 0 , 5 M i o J a h r e n .
Nicht mehr v o r h a n d e n : Leptobos elatus, Macaca florentina, Talpa pr'dglacialis, Ursus etruscus,
neu: Desmana tegelensis, Sus strocci, Desmana tegelensis, Hystrix etrusca, Dolomys milleri, Trogontherium cuvieri.
4 . A m E n d e der Cromer-Warmzeit, v o r 0 , 3 2 M i o J a h r e n .
Nicht mehr v o r h a n d e n : Equus stenonis, Archidiskon meridionalis, Trogontherium, neu: Ursus spelaeus, Sorex, Discrostonyx Microtus, Desmana mosebata mosbachen-sis, Ovibos, Bison schoetensacki, Caprea, Megaloceras, Orthogonoceras, Capreolus, Rangifer, Palaeoxolon antiquus, Mammontheus trogontherium, Hippopotamos, Machairodus, Equus süssenhornensis, E. mosbachensis, Sus scrofa.
( E r s a t z v o n L e p t o b o s durch Bos u n d Bison, der „ Q u a g g a s " durch caballische Pferde und v o n Archidiskon meridionalis durch Mammontheus trogontheri, D I E T R I C H
1 9 5 3 ) .
5 . A m E n d e der E l s t e r - K a l t z e i t vor 0 , 1 8 M i o Jahren.
Nicht mehr v o r h a n d e n : Archidiskon meridionalis, Hystrix refossa, Orthogonoceras, Dolicbodoryceras, Hippopotamus, Equus mosbachensis,
neu: Ursus aretos, Gulo.
6. A m E n d e der Eem-Warmzeit, vor e t w a 0 , 0 8 Mio J a h r e n .
Nicht mehr v o r h a n d e n : Megaloceras, Bison priscus, Equus caballus, Dicerorhinus etruscus, Die. kirchbergensis, Bos primigenius, Buffelus, Saiga tatarica, Palaeoloxo-don antiquus,
neu: Mammontheus primigenius, Equus caballus, E. germanicus, Coelodonta anti-quitatis, Buffelus, Saiga tatarica.
Aus der allmählich dichteren Aufeinanderfolge (wohl kein Trugschluß wegen der zu nächst dürftigeren Belege) der „Faunenwellen" ergibt sich für das Altpleistozän ein lang sameres, für d a s J u n g p l e i s t o z ä n ein sich steigerndes T e m p o der Evolution. A b e r so wenig der vorgeschichtliche Mensch durch die J a g d die Wirbeltierwelt dezimiert hat ( S O E R G E L 1 9 2 2 ) , so gewiß ist dem H o m o sapiens sapiens die derzeitige Verarmung der Wirbeltier welt zur L a s t zu legen.
Auch in der Entwicklung der Hominiden scheinen Haupteinschnitte an der Grenze Plio/Pleistozän u n d um die C r o m e r - W a r m z e i t zu liegen. Die Plio-Pleistozän-Grenze b e deutet das U b e r g a n g s f e l d von der „subhumanen P h a s e " der Hominiden über d a s Tier hin aus zur humanen P h a s e in Gestalt der Australopithecinen, welche als „ p y g m o i d e klein hirnige Aufrechtgänger mit typischen Standfüßen und grundsätzlich menschlichem G e b i ß " ( H E B E R E R ) sich schon mancher Werkzeuge und Geräte bedienten. Ihnen folgten die F r ü h menschen mit (Pithecanthropus) H o m o erectus (heidelbergensis als ältester Fund in der Elster-Kaltzeit) u n d diesen verhältnismäßig rasch aufeinander die A l t - ( H o m o sapiens neanderthalensis) u n d Jetztmenschen ( H o m o sapiens praesapiens u. H . sapiens sapiens) in der letzten W a r m - u n d Kaltzeit ( H E B E R E R & K U R T H 1 9 6 6 ) . Also anscheinend wie bei den Wirbeltieren allgemein eine beschleunigte Evolution.
Bei den marinen M o l l u s k e n zeichnen sich der Einbruch der K a l t z e i t e n und der seitdem waltende Z y k l u s von K a l t - W a r m z e i t e n mit der Mischung und A b l ö s u n g lusitani-scher durch boreale Formen ab (vgl. Verzeichnis der Schnecken und Muscheln bei VAN D E R K L E R K & F L O R S C H Ü T Z 1953). Mit dem Aufhören besonders vieler alter und dem Erschei nen neuer Arten hebt sich die C r o m e r - W a r m z e i t als Wende ab, während Holstein-See und E e m - M e e r sich durch das Ausscheiden weniger Formen (Corbula fluminalis, Vivipara dilu-viana, Valvata naticina) voneinander unterscheiden.
Eine gegensätzliche Entwicklung lehrt die Geschichte der pleistozänen Vegetation (so weit bei der Verschiedenheit pflanzlicher und tierischer B a u p l ä n e überhaupt ein Vergleich möglich ist). N a c h d e m bereits im U n t e r o l i g o z ä n Mitteleuropas die ersten „ a r k t o t e r t i ä r e n " Floren-Elemente erschienen waren ( A H R E N S U. a. 1 9 6 8 ) , kommen sie in der Folgezeit in einem oftmaligen Wechsel zum Ausdruck. Im Pliozän aber kam es zu erheblichen Schwan kungen in der Beteiligung typischer Tertiärformen ( T a b . 4 ) . Noch bleiben einschneidende Absenkungen der T e m p e r a t u r aus, aber im Reuver-Abschnitt partizipieren als Vorboten einer erheblichen A b k ü h l u n g Betula und Pinns silvestris zeitweilig mit 50 °/o an der Z u sammensetzung der Baumpollen, w ä h r e n d die tertiären Formen auf den bis dahin noch nicht erreichten T i e f s t a n d von wenig über 10 °/o zurückgehen (Tab. 4 ) . Diese Tendenz setzt sich fort, bis in der Cromer-Warmzeit der U m b a u der tertiären Waldvegetation in die pleistozäne warmzeitliche Waldvegetation vollzogen ist ( F R E N Z E L 1968). V o n nun an läuft in jeder Warmzeit eine schnelle Folge scharf getrennter Waldtypen ( B i r k e / K i e f e r , Kiefer/ Birke/Fichte/Ulme, Eichenmischwald, Hainbuchen/Tanne oder Carpinus/Fichte, Kiefer/ Fichte/Birke/Kiefer) a b , nur in der H o l s t e i n - W a r m z e i t durch eine starke Vorherrschaft v o n N a d e l h ö l z e r n verschleiert.
Es bedurfte mithin der Einwirkung von K a l t - und Warmzeiten über 1,3 Mio J a h r e hindurch, um die tertiären Baumpollen in Mitteleuropa auszuschalten. A b e r während die Wirbeltiere auf diese periodischen I m p u l s e einer veränderten Umwelt mit der Schöpfung ganzer Ketten v o n Stammesreihen reagierten, verhielten sich die Pflanzensippen physiolo gisch konservativ u n d wenig variabel. Abgesehen von der fortschreitenden V e r a r m u n g an tertiären Elementen konsolidierte sich die pleistozäne Vegetation in der B i l d u n g zweier S t a n d a r d t y p e n , die sich als Siedlungsgemeinschaften von K ä l t e s t e p p e n mit nur (für Mitteleuropa) k u r z z e i t i g und schwach ausgeprägter T u n d r a und als Eichenmischwald mit einer N a d e l w a l d - P h a s e am Anfang und am Ende präsentierten. Beide besetzten immer wieder je K a l t - oder Warmzeit die gleichen Areale ( F R E N Z E L 1968), wobei sie sich aus den Refugien des Mittelmeerraumes oder den nordeuropäischen Tundren ergänzten. D a b e i er wiesen sich Sumpf- u n d Wasserpflanzen als besonders konservativ. D i e großen Einschnitte in der Entwicklung während des Pleistozäns liegen — wie bei den Wirbeltieren — im Früh-Abschnitt des Villafranc und in der C r o m e r - W a r m z e i t , so daß eine Wechselwirkung naheliegt. Wie die Evolution der Tierwelt verlangt auch die Vegetationsgeschichte des Pleistozäns die 1,3 M i o J a h r e als Frist für die Reifung. Diese große und entscheidende Zeit
Versuch einer neuen Pleistozän-Gliederung 105
Tabelle 4
D e r A n t e i l t y p i s c h e r B a u m p o l l e n a n d e r V e g e t a t i o n v o n O b e r p l i o z ä n u n d P l e i s t o z ä n ( i m w e s e n t l i c h e n n a c h Z A G W I J N
a u s F R E N Z E L 1 9 6 8) i n N W - E u r o p a
Z eitweiliger maximaler Anteil an Zeit-Abschnitte Typische
Tertiärformen
Eichenmisch wald
Pinus
silvestris Betula Picea i n P r o z e n t e n Eem-Warmzeit 0 80 90 90 12 Holstein-Warmzeit 0 10 70 10 20 Cromer-Warmzeit 1—0 80 85 70 15 Waal-Warmzeit 4 45 Tegelen-Warmzeit 8 12 80 6 55 Prätegelen 2 3 Reuver (Asti) 12—38 40 50 35 4 Brunssumian C 20—70 B 20—72 A 30—70 (Piacentin) Susterian (Pont) 20—30
zwischen P l i o z ä n und Cromer-Warmzeit ist geologisch und biologisch (Gleichsetzung von Villafranc u n d Tegelen, A n n a h m e nur weniger Kaltzeiten v o r der Tegelen-Warmzeit) nicht genügend gewürdigt. In den großen Einschnitten der L e b e w e l t vor 1,6 und 0 , 3 Mio J a h r e n d a r f m a n außer einer Verzahnung biologischer Zeitenwenden ( D I E T R I C H 1 9 5 3 ) auch eine, im großen gesehen ständige, noch nicht beendete A b k ü h l u n g seit dem Pliozän erblicken. D a s wäre auch insofern nicht unwahrscheinlich, als die Gegenwart erst beim 2 5 . Zyklus der Kaltzeiten angelangt ist und (wie im P e r m o k a r b o n ) ein Vielfaches noch bevorsteht.
Überblickt man die Entwicklung von F a u n a und Flora im Q u a r t ä r , die D o k u m e n t i e rung von K a l t - und Warmzeiten durch M o r ä n e n , Lösse, Terrassen und Strandstände einer seits und d a s Ergebnis verschiedener Untersuchungsmethoden zu ihrer zeitlichen, räum lichen und klimatischen E r f a s s u n g andererseits, so ergibt sich ein im großen gerichteter und zyklisch gearteter Ablauf. D i e jeweils fixierte organische und anorganische Materie ent spricht dabei in ihrer Ausbildung sozusagen Variationen z u m gleichen T h e m a , nämlich dem Arrangieren mit den durch K a l t - und Warmzeiten gebotenen Verhältnissen. Diese begrenzte V a r i a b i l i t ä t innerhalb der Z y k l i k ist durch den komplexen C h a r a k t e r der steuernden F a k t o r e n ( P a l ä o t o p o g r a p h i e , sinkende Temperaturen, weichende N o r d s e e , extratellurische Ursachen usw.) begreiflich.
Die Bildungen des Pleistozäns müssen sich jedesmal ihren R a u m und ihre S u b s t a n z größtenteils a u f Kosten vorangegangener A b l a g e r u n g e n selbst schaffen. Infolgedessen sind Moränen, Terrassen und L ö s s e wegen ihrer lockeren Beschaffenheit und exponierten L a g e häufig verschwunden oder aber nach M o r p h o l o g i e und Zusammensetzung oft nicht von ihren V o r g ä n g e r n zu unterscheiden. D i e g e o l o g i s c h e D i a g n o s e steht deshalb vor einer schwierigen A u f g a b e . Sie kann sich aber der mannigfachen (von der B o d e n k u n d e , Paläontologie, Petrographie, Morphologie, den physikalischen A l t e r s und T e m p e r a t u r -Bestimmungen gebotenen) Indizien in gemeinsamer A b s t i m m u n g bedienen. S i e bleiben jedoch so l a n g e erdgeschichtlich beziehungslos, bis für ihre E i n o r d n u n g die spezifisch
plei-stozäne zyklische Periodik zur V e r f ü g u n g steht. D a ß diese einer Z y k l i k entspricht, b e d a r f angesichts der kalt-warmzeitlichen, durch F a u n a u n d F l o r a widergespiegelten und durch physikalische D a t e n zeitlich bestätigten Sedimentations-Folge keiner weiteren Beweisfüh rung, wenn auch die letzten Ursachen dafür unbekannt sind. A u f der Grundlage der Z y k l i k ist die vorstehende Gliederung des Pleistozäns entworfen, eine in sich konsequente Aufteilung, z w a r immer noch unvollkommen, aber m i t weniger Lücken als bisher, in einer natürlichen O r d n u n g und mit der Möglichkeit, alte sowie neue Beobachtungen und D a t e n erdgeschichtlich einzuordnen.
Mag m a n diesen Versuch als weitere Variation der bereits vorhandenen zahlreichen Hypothesen skeptisch aufnehmen u n d als verfrüht ansehen, so kann er gleichwohl bei der Reformbedürftigkeit des klassischen Eiszeiten-Systems wegen seines unleugbaren Zerfalls in mehrteilige K a l t und Warmzeiten als Lückenbüßer dienen. D a s Verharren im O l i g o -glazialismus engt den Blick für stratigraphische Vergleiche sowie für die Geschichte der T i e r - und Pflanzenwelt angesichts der vermehrten F a k t e n ein. D a s hier über die bisherigen Aufteilungen hinausgehende S c h e m a soll lediglich eine Möglichkeit z u r w a h l f r e i e n E i n o r d n u n g alter u n d neuer Pleistozän-Bildungen bieten und für die B e urteilung der tierischen und pflanzlichen Entwicklung mehr Stationen, v o r allem im A l t pleistozän, zur Verfügung stellen. W a s bei dieser Gelegenheit über E v o l u t i o n gesagt ist, hat mehr die Bedeutung einer Fragestellung zur Wechselwirkung v o n geologischem u n d biologischem Geschehen als einer abschließenden Feststellung, weil die K a u s a l z u s a m m e n h ä n g e zwischen Evolution und U m w e l t ä n d e r u n g e n noch nicht geklärt sind.
L i t e r a t u r h i n w e i s e
AHRENS, H . U. a.: Zur Plio/Pleistozän-Grenze in der Deutschen Demokratischen Republik. — 23. intern, geol. Congress, 10, 65—77, 5 Abb., Prag 1968.
BOENICK, W.: Zur Kenntnis des Altquartärs bei Brüggen (westlicher Niederrhein). — Sonderveröff. geol. Inst. Köln, 1 7 , 138 S., 20 Abb., 7 Tab., 3 T a f , Köln 1969.
BRUNNACKER, K . : Der Villafranchium-Löß bei St. Vallier. — N . Jb. Geol. Paläont., J g . 1967, 257—267, 1 Abb., 1 Tab., Stuttgart 1967.
BRUNNACKER, K., STREIT, R. & SCHIRMER, W.: Der Aufbau des Quartär-Profils von Kärlich/Neu
wieder Becken (Mittelrhein). — Mz. naturw. Arch., 8 , 102—133, 8 Abb., 3 Tab., 1 Taf., Mainz 1969.
DIETRICH, W. O.: Neue Funde des etruskischen Nashorns in Deutschland und die Frage der Villa-franchium-Faunen. — Z. Geol., 2, 417—430, 1 Abb., Berlin 1953.
EMILIANI, C . : Palaeotemperature Analysis of Core 280 and pleistocene Correlations. — Journ. of Geol., 6 6 , 265—275, 5 Abb., 3 Tab., Chicago 1958.
FRECHEN, J . & LIPPOLT, H. J . : Kalium-Argon-Daten zum Alter des Laacher Vulkanismus, der Rheinterrassen und der Eiszeiten. — Eiszeitalter u. Gegenwart, 1 6 , 5—30, 8 Abb., Öhringen 1965.
FRENZEL, B . : Grundzüge der pleistozänen Vegetationsgeschichte Nord-Eurasiens. — Erdwiss. Forsch., 1 , 326 S., 17 Taf., 67 Abb., Wiesbaden 1968.
HEBERER, G. & KURTH, G.: Die (Eu)Hominiden vom Mittelpleistozän bis ins mittlere Jung pleistozän. — Handbuch der Urgeschichte, I, 209—233, 1 Taf., 5 Abb., 1 Tab., Bern 1966. HESEMANN, J . : Das Münsterland als pliozäne Erosionslandschaft. — N . J b . Geol. Paläont. Mh.,
Jg. 1969, 530—534, 3 Tab., Stuttgart 1969.
JESSEN, W.: Zur Sedimentologie des Karbons. — 4. Congr. Avancement Etudes Strat. Geol. Car-boniferes, C. R., 307—322, Maastricht 1961.
KAHLKE, H . - D . : Vertebratenstratigraphie zur Plio/Pleistozän-Grenze. — Report 23. Intern, geol. Congress, Section 10, 27—39, Prag 1968.
KRETZOI, M.: Stratigraphie und Chronologie. Czwartorzed europy Srodkoweji Wschodniej. — Inqua, 1961, 313—329, Lodz 1964.
MESOLELLA, K. M., MATTHEWS, R., BROECKER, W. S. & THURBER, D . L . : The astronomic Theory
of climatic Change.: Barbados Data. — J . Geol., 7 7 , 250—274, 13 Abb., Chicago 1969. QUITZOW, H . W.: Mittelrhein und Niederrhein. — Beitr. z. Rheinkde., 1 3 , 27—39, 4 Abb.,
Koblenz 1962.
SEGOTA, T.: Paläotemperature Changes in the Upper and Middle Pleistocene. — Eiszeitalter u. Gegenwart, 1 8 , 127—141, 3 Abb., Öhringen 1967.
Versuch einer neuen Pleistozän-Gliederung 107
SOERGEL, W.: Die Jagd der Vorzeit. — 149 S., 28 Abb., 1 Tab., Jena 1922. — : Die Vereisungskurve. — 87 S., 1 Taf., 3 Tab., Berlin 1937.
TOEPFER, V . : Tierwelt des Eiszeitalters. — 46 Abb., 20 Taf., 2 Tab., Leipzig 1963.
VLERK, I . M. VAN DER & FLORSCHÜTZ, F . : The palaeontological Base of the Subdivision of the Pleistocene in the Nederlands. — Verhandl. Koninkl. nederl. Akad. van Wetensch. afd. N a -tuurkunde, 1, reeks, 20, N r . 2, 58 S., 29 Tab., 4 Taf., Amsterdam 1953.
ZEUNER, F . E . : Schwankungen der Sonnenstrahlung und des Klimas im Mittelmeergebiet während des Quartärs. — Geol. Rdsch., 30, 650—656, 1 Tab., Stuttgart 1939.
Manuskr. eingeg. 13. 7. 1970. Anschrift des Verf.: Prof. Dr. J . Hesemann, 415 Krefeld, Von-Steuben-Straße 17.