Turkish Journal of Fisheries and Aquatic Sciences 13: 331-340 (2013)
www.trjfas.org ISSN 1303-2712 DOI: 10.4194/1303-2712-v13_2_15
© Published by Central Fisheries Research Institute (CFRI) Trabzon, Turkey in cooperation with Japan International Cooperation Agency (JICA), Japan
Molecular Cloning and Expression Analysis of Extra Sex Combs Gene
during Development in
Macrobrachium nipponense
Introduction
The polycomb group (PcG) genes are chromatin regulators of the homeotic and other developmental genes found in many different organisms, including fruit fly (Struhl and Akam, 1985; Lewis et al.,1978; Li et al., 2012), mammals (Morey et al., 2010), and even plants (Kohler et al., 2010). These genes have received a lot of attention due to their important roles in the control of tissue and organ development during embryogenesis, tumorigenesis, chromosome X-inactivation, genomic imprinting, and so on (Oktaba et al., 2008; Bracken et al., 2009; Zhang et al., 2012). At least 11 PcG genes have been described and there may be up to 40, including Additional sex combs (JÜrgens, 1985), Enhancer ofzeste(Jones and Gelbart, 1990), extra sex combs (Struhl, 1981), pleiohomeotic (Girton and Jeon, 1994), polycomb (Lewis, 1978), polycomblike (Duncan, 1982), polyhomeotic (Dura et al., 1985), posterior sex combs (JÜrgens, 1985), sex combs extra (Breen and Duncan,1986), sex combs on midleg (JÜrgens, 1985), super sex combs (Ingham, 1984) and enhancer of polycomb (Sato et al., 1983). The extra sex combs (Esc) gene, originally identified in Drosophila melanogaster, was found to be
expressed primarily during the early embryogenesis (Gutjahr et al., 1995; Sathe et al., 1995; Simon et al., 1995) and play important roles in determining segment identity (Struhl et al., 1981) and in hedgehog signaling pathway (Norihisa et al., 2004). Its product is essential for histone H3 K27 methylation and the establishment of PcG silencing of homeotic genes of the Bithorax and Antennapedia complexes (Katsuhito et al., 2008). The Drosophila embryos lacking maternal Esc displayed complete derepression of all homeotic genes and conversion of all segments to the identity of the 14th parasegment (Struhl et al., 1981). In mouse, considerable evidence also suggested mutation of Esc homolog embryonic ectoderm development (EED) resulted in embryonic lethality (Faust et al., 1998). In Medaka, hypomorphic knock-down of oleed (Oryzias latipes embryonic ectoderm development) using morpholino antisense oligonucleotides resulted in the fusion of eyes (Norihisa et al., 2004). Taken together, these observations support Esc as one of the best biomarkers for development of embryogenesis and larvae, which was experimentally demonstrated in the houseflies, butterflies and grasshoppers (Simon et al., 1995; Joyce et al., 1997). Previous study confirmed
Yanping Zhang
1,3, Sufei Jiang
2, Yiwei Xiong
2, Shengming Sun
2, Hui Qiao
2, Shubo Jin
2,
Yongsheng Gong
2, Hongtuo Fu
1,2,*1
Nanjing Agricultural University, Wuxi Fishery College, Wuxi 214081, China.
2
Ministry of Agriculture, Freshwater Fisheries Research Center, Key Laboratory of Freshwater Fisheries and Germplasm Resources Utilization, Chinese Academy of Fishery Sciences,Wuxi, 214081, China.
3
Ministry of Agriculture, Jiangxi Fisheries Research Institute, Scientific Observing and Experimental Station of Fishery Resources and Environment in Poyang Lake, Nanchang, 330039, China.
* Corresponding Author: Tel.: +86.510 85558835; Fax: +86.510 85558835; E-mail: [email protected]
Received 31 December 2012 Accepted 25 February 2013
Abstract
The extra sex combs (Esc) gene first identified in Drosophia encodes a transcriptional repressor of homeotic genes belonging to the polycomb group (PcG) gene and is primarily expressed in the early embryos. In this study, we have isolated the full-length cDNA of an Esc gene from the testis of oriental river prawn (Macmbrachium nipponense) according to the established expressed sequence tags (ESTs) information using Rapid Amplification of the cDNA Ends (RACE) technique, designated as MnEsc. The full-length cDNA of MnEsc is 1,461bp in size and has an open reading frame (ORF) of 1,068bp, encoding a 355-amino acid protein. Real-time quantitative PCR analyses revealed that the expression level of MnEsc varied significantly in the developing embryo, postembryonic larval and adult tissues. Real-time quantitative PCR showed the MnEsc gene was expressed in testis, liver, ovary, brain, abdominal ganglion, heart, intestine and muscle with the highest level of expression in ovary and brain. In vivo silencing of the gene, the dsRNA of MnEsccaused a significantly decrease in target gene expression level in brain and ovary tissues, but no exterior appearance change of experimental shrimp was observed.
that the crustaceans have dramatically close evolution relationship with insects (Budd et al., 2009), we hypothesize that MnEscis involved in the embryonic development and phenotypic differentiation in crustaceans.
The oriental river prawn, Macrobrachium nipponense (Crustacea; Decapoda; Palaemonidae) is a commercial freshwater prawn. It is considered as an important fishery resource in China with an annual production of 205,010 tons (Bureau of Fishery, 2009). The identification of a germ cell specific marker in mitten crab will aid in the study of the molecular event involved in the determination of the development of these cells (Wang et al., 2012). In recent years, our laboratory has performed some molecular research on the M. nipponense development system. In this study, we cloned a full length Esc cDNA from M. nipponense and analyzed its expression pattern throughout developmental stages, which could help to understand the regulatory mechanism of early embryonic development, postembryonic development and phenotypic differentiation in the oriental river prawn. In addition, we also investigated the ability of dsRNA to inhibit MnEsc mRNA expression in M. nipponense in vivo.
Materials and Methods
Embryo, Larvae and Tissue Collections
Several female and male healthy adult shrimps with wet weight of 1.26~4.25g were obtained from Tai Lake in Wuxi, China (120°13'44"E, 31°28'22"N). All of these samples were transferred to lab breeding conditions and maintained in a 500-liter tank with aerated freshwater for 72 h before tissue collection. The different developmental stages of eggs and larval were obtained from our breeding room. After prawn spawning, each developmental stage of embryos (cleavage stage; blastula stage; gastrula stage; nauplius stage; protozoea stage; zoea stage) was collected according to morphological methods following the criteria by Chen et al. (2012). Larvae were collected every 3 days between 1 day post-hatching larvae (L1) and L13. Post-larvae were collected every 4 days from 1 to 20 day after the metamorphosis (P1~P20), and every 10 days from 21 to 30 day. A variety of tissues, including ovary, testis, muscle, heart, abdominal ganglion, brain, liver and intestine were also collected. The ovary was collected from mature female shrimps, but testis, muscle, heart, abdominal ganglion, brain and intestine were collected from mature male shrimps. The samples were washed with 1 × PBS (phosphate-buffered saline, 0.01 M), frozen directly in liquid nitrogen and stored at -80 °C until processed.
RNA Isolation and Reverse Transcription (RT)
Total RNA was extracted from embryos and
larvae at different stages of development and was also from other tissues in mature shrimps with RNAiso Plus Reagent (TaKaRa Bio Inc., Japan) in accordance with the protocol of the manufacturer. The isolated RNA was treated with RNase-free DNase I (Sangon, shanghai, China) to eliminate possible genomic DNA contamination. The concentration of each total RNA sample was then measured by BioPhotometer (Eppendorf, Hamburg, Germany), and 2μL was analyzed on a 1% agarose gel to check the integrity. The first-strand cDNA synthesis for real-time quantitative PCR was performed for each RNA using 1μg of total RNA, 4μL 5× iScript reaction mix (BIO-RAD, CA, USA) and 1μL iScript reverse transcriptase in a final volume of 20 μL. The reaction was incubated at 25℃ for 5min, then 42℃ for 30min followed by 85℃ for 5min. The cDNA of real-time quantitative PCR was kept at -20℃.
5′- and 3′-RACE Amplification
Total RNA was extracted from testis of mature shrimps. Concentrations were measured by BioPhotometer (Eppendorf, Hamburg, Germany) and 1μL was analysed on a 1% agarose gel to check the integrity. The full length of the MnEsc gene was obtained by using the 5′ RACE cDNA and 3′ RACE cDNA amplification kit (TaKaRa Bio Inc., Japan). 3′ and 5′-RACE cDNAs were then used for the 3′/5′-RACE PCR with 3′ gene-specific primer (3GSP1, 3GSP2) and 5′ gene-specific primer (5GSP1, 5GSP2) that were designed on the basis of EST sequence of MnEsc (GenBank accession no. JK526490) obtained from the M. nipponense testis cDNA library (Qiao et al., 2012). All primers used in this study were shown in Table 1. The two primers of Esc-F and Esc-R validated the MnEsc fragments from the EST cDNA library. The 3′-RACE and 5′-RACE were performed with 3′-full RACE Core Set Ver.2.0 Kit and 5′-full RACE Kit (TaKaRa Bio Inc., Japan) according to the manufacturer’s instructions.
The PCR fragments were subjected to electrophoresis on a 1.5% agarose gel to compare the length difference. Amplified cDNA fragments were cloned into the pMD18-T vector (TaKaRa Bio Inc., Japan) following the instructions provided by the manufacturer. The recombination was then transformed into the ESCherichia coli DH5α (Qiagen, Germany) competent cells which were identified by blue/white screening and confirmed by PCR. At least five positive clones were sequenced in both directions using an automatic DNA sequencer (Applied Biosystems ABI-3730, USA) and these resulting sequences were verified and subjected to cluster analysis in NCBI.
Bioinformatics Analyses
and BLASTN program (Altschul et al., 1990). The protein prediction was performed using the ORF Finder tool (http://www.ncbi.nlm.nih.gov /gorf/gorf.html). The motif was performed with the motif scan program (http://hits.isb-sib.ch/cgi-bin/motif scan/). Esc deduced amino acid sequences from M. nipponense and representative invertebrates were compared by multiple sequence alignment using ClustalX. A neighbor-joining (NJ) phylogenetic tree was constructed using MEGA4.0 (Tamura et al., 2007).
Real-Time Quantitative PCR Expression Analysis
The MnEscmRNA expression at different stages from embryo to post-larva, and various adult tissues was measured by aSsoFastTM EvaGreen® (BIO-RAD, CA, USA) real-time quantitative PCR analysis in Bio-Rad iCycler iQ5 Real-Time System (BIO-RAD, CA, USA). Gene-specific primers (Table 1) were used to amplify the Esc transcript, and the PCR products were sequenced to verify the specificity of the PCR primers. Amplifications were performed on a 96-well plate with a 25μL reaction volume containing 1μL cDNA (50ng), 10μL SsoFastTM EvaGreen® Supermix (BIO-RAD, CA, USA), 0.5μL 10μM of gene specific forward and reverse primers (Table 1), and 13 µL of DEPC-water. The PCR temperature profile was 95°C for 30 s followed by 40 cycles of 94°C for 15 s, 58°C for 20 s and 72°C for 20 s , with a 0.5°C/5s incremental increase from 60°C to 95°C. Each sample was run in triplicate along with the internal control gene. To ensure that only one PCR product was amplified and detected, the dissociation curve analysis of amplification products was performed at the end of each PCR reaction. Amplification of β-actin as an internal reference was also carried out in the same sample (Table 1). The relative copy number of MnEsc mRNA was calculated according to the 2-⊿⊿CT
comparative CT method (Livak et al., 2001).
ds RNA Preparation
PCR fragments containing M. nipponense Esc or green fluorescent protein (GFP) open-reading frame were amplified using gene-specific primers containing T7 promoter site at the 5′ ends of the gene-specific primers. The MnEsc dsRNA and GFP dsRNA synthesis primers are shown in Table 1. The PCR products were purified with a gel extraction kit (Sangon, shanghai, China). dsRNA was synthesized in vitro using Transcript AidTM T7 High Yield Transcription kit (Fermentas, Inc., USA) according to the manufacturer’s instructions. The dsRNA was purified by ethanol precipitation and dissolved in RNase-free water. Purity and integrity of the dsRNA were examined by standard agarose gel electrophoresis. Concentration of the dsRNA was measured at 260 nm by using a BioPhotometer (Eppendorf, Hamburg, Germany), and then kept at -20℃ until used.
For the short-term in vivo dsRNA injection experiment, 30 health mature female M. nipponenses (each weighing 1.6-2.3g) were selected to inject into heart. The female shrimps were assigned to three treatment groups: MnEsc-dsRNA injected (n=10), GFP-dsRNA injected (n=10) and vehicle injected (n=10). Each shrimp was injected with 4μg MnEsc-dsRNA or 4μg GFP-MnEsc-dsRNA, or a similar volume vehicle. The MnEsc mRNA expression of the brain and ovaries were investigated to detect the interference efficiency by real-time quantitative PCR after injection for 4 days and 1 week.
Statistical Analysis
All data were presented as mean ± SE (standard error of the mean). Statistical analysis was performed
Table 1. Primers used in this study
Name Sequence (5′→3′) Primer description
Esc-F Esc-R Esc-3GSP1 Esc-3GSP2 Esc-5GSP1 Esc-5GSP2 3′RACE OUT 3′RACE IN 5′RACE OUT 5′RACE IN
TCACTTGCTACCATGCAGTGCG TCCCAACAGCTAGCACTTTTGC CCTTGTGGCTACCTTTGG GTTGGGAACCAGTTAGGC
AGGTAGCCACAAGGATATCTGTCCTG TGCTGCTAAGATTGGCCTTCCTG TACCGTCGTTCCACTAGTGATTT
CGCGGATCCTCCACTAGTGATTTCACTATAGG CATGGCTACATGCTGACAGCCTA
CGCGGATCCACAGCCTACTGATGATCAGTCGATG
Esc fragment
FWD first primer for 3′ RACE FWD second primer for 3′ RACE RVS first primer for 5′ RACE RVS first primer for 5′ RACE RVS first primer for 3′ RACE RVS second primer for 3′ RACE FWD first primer for 5′ RACE FWD second primer for 5′ RACE Esc-RTF
Esc-RTR β-actinF β-actinR Esc-dsF Esc-dsR GFP-F GFP-R
TGGCTACCTTTGGTGGAGTAGAG TGCAGGTGCCCTCAATGTC TATGCACTTCCTCATGCCATC AGGAGGCGGCAGTGGTCAT GAAGACCATGGACAGCCAAT GAATGATCCATGCCACATGA GTCAGTGGAGAGGGTGAAGG AAAGGGCAGATTGTGTGGAC
FWD primer for Esc expression RVS primer for Esc expression
FWD primer for β-actin expression RVS primer for β-actin expression For MnEsc dsRNA synthesis
using SPSS13.0. Statistical significance was determined using one-way ANOVA and post hoc Duncan multiple range tests. Significance was set at P<0.05.
Results
Cloning and Characterization of the MnEsc Gene
The nucleotide sequence reported in this study has been deposited in the GenBank DNA database under Accession No. JX221051. The full-length cDNA sequence of MnEsc was comprised of 1,461bp with a 1,068bp open reading frame (ORF) encoding 356 amino acid residues which display an estimated molecular mass of 40.201kDa and the isoelectric point of 7.09. The 5’ and 3’ untranslated region
(UTR) contain 105 bp and 288 bp, respectively. The clone included the canonical polyadenylation signal (AATAAA), typical of eukaryotic mRNA, and poly (A) tail. Conserved sequence and characteristic motifs of Five WD40 repeats were identified in the deduced amino acid sequences of MnEsc (Figure 1). In addition, the N-terminal region contains a potential nuclear localization signal (NLS) (residues 42-54, Figure 1).
Secondary structure of MnEsc was composed of alpha helix (1.1%), extended strand (51.0%) and Loop (47.9%). Matured protein composed of 6 casein kinaseⅡphosphorylation sites, 6 protein kinase C phosphorylation sites, an amidation site, 5 N-myristoylation sites, and 3 N-glycosylation sites (Figure 1). It has multiple potential functional sites for phosphorylation, Methylation and glycosylation.
Homology and Phylogenetic Analysis of MnEsc
Bioinformatic analyses with BLASTx searches of the public DNA sequences showed that the coding sequence (CDS) of M. nipponense displayed a high degree of identity with many Escs of arthropods. Sequence comparisons of the Esc deduced amino acids showed similarity of 67%, 65%, 64% and 63% to that of the Schistocerca Americana (AAC05332.1),
Danaus plexippus (EHJ72379.1), Junonia coenia (AAC05331.1) and Bombyx mori (NP_001188366.1), respectively (Figure 2). Furthermore, based on the results of the alignment of Esc sequences of the different organisms, the phylogenetic trees were constructed based on the neighbor-joining method using the complete Esc proteins deposited in NCBI by ClustalW1.81 and MEGA4.0. The NJ tree showed that MnEsc is closely related to Lepeophtheirus
26 M.nipponense Esc
38 P.humanus corporis Esc
40 S.americana Esc 15 D.plexippus Esc 15 B.mori Esc 23 L.salmonis Esc 22 D.melanogaster Esc . . M . . . . . . S . . . . . M P . . . . . S V . . . . . G T . . . . . N S . . . . . N S . . . . . S A . . . . . L K . . . . . K S . . . . . V K . . . . . K V . . . . . S M . . . . . L K . . . . M K N . . . . T N S . . . . P V H . . . . N N F . . M . G P H . . V M D N S . . S S D S E . . E S S D Q . . K D L I S . . R K S S T . . G V F N N . . R K L L T M M Y N D T E N N A G E E D F F E N S D S S S A E G S G D E S P D G D N N D E D E D E E S E L E A A A D S D D D D D D E D E E D D T E C T T T T D S S S S S S D C S S S S S S G V V V V V L D E G G E D D E 61 M.nipponense Esc 77 P.humanus corporis Esc
77 S.americana Esc 51 D.plexippus Esc 50 B.mori Esc 63 L.salmonis Esc 60 D.melanogaster Esc S S S S S S S G A T T T G A S S S S S P S N T T N N L Y A T T T T T T D D D D D R T N N N N N K N S T T T T R S . S S S S G T R R R R R A T S S S S S A S G E E E E L R T T T T T A S P P P P P S K T T T T T N S Q P N N N G P . T R . . S S . N . . . M . . F . . . S . . K . . . R S R G G T S K S R R R R R G T K K K V V R R G G G K K K S R K R K K K K A R R R R C R N S G R R S R K R R R R S G R K K G G S R G K K K K Q R R S I K K N S K . Q K . N T K G Q A K S K V K K V T F S I T I T T T K G V N K K G P P T N P P G K A S M V V R S K K K K K K R M L L P P L A 101 M.nipponense Esc 117 P.humanus corporis Esc
117 S.americana Esc 91 D.plexippus Esc 90 B.mori Esc 101 L.salmonis Esc 100 D.melanogaster Esc G Q Q P P N A Y Y Y Y Y Y Y K K K K K K K F F F F F C Y G V S N N M D C C T C C G T Y Y S S S H H L V V A A W V K K K K K R K E E E E E E E D D D D D D N H H H H H H H G G G G G Q G Q Q Q Q Q Q A P P P P P P N I I L L L I I F F F F F F F G G G G G G G V A A C C V V Q Q Q Q Q A A F F F F F M F N N N N N N N Q H H H H H T H H H H H H L L L L L L L L R K K R G D G E K E E E D K G G G G G . D Q E Q E E . E P P P P P P P L L L Q S R Q V I I I V V V F F F F F F F A A A A A A A T S A V V T T A V V V V V A G G G G G G G N S S S S N S N N N N N N N R R R R R R R 141 M.nipponense Esc 157 P.humanus corporis Esc
157 S.americana Esc 131 D.plexippus Esc 130 B.mori Esc 141 L.salmonis Esc 140 D.melanogaster Esc I V V V V V V S S T S S T T V I V I I V V Y Y Y Y Y Y Y Q R E E E E E C C C C C A C I D P P P L P E E E E E S R D N G S S N Q G G S G G G G S S G G G D G I I I F F V M K S K K K K Q L L L F F L L L L L L L L L R Q Q Q Q Q H C C C C C S C Y Y Y Y Y Y Y S A A A A A A D D D D D D D P P P P P P P D D D D D D D T T V V V A P E D D D D D D E E E E E E E N N N T T N V F Y Y F F F F Y Y Y Y Y Y Y T T T T T S T V C C C C V C A A A A A A A W W W W W W W S S S S S S S Y Y Y Y Y Y Y D D E E E D D T V E E E P L E E E E D S K T T S T T D T G G G G M G S R N K L L K S 181 M.nipponense Esc 197 P.humanus corporis Esc
197 S.americana Esc 171 D.plexippus Esc 170 B.mori Esc 181 L.salmonis Esc 180 D.melanogaster Esc P P P P P P P I Y L L L L L L L L L L L L A A A A A A A A V V V V A A A A A A A A A G G G G G G G S S S S S S Y R R R R R R R G G G G G G G V V I I I I V I I I V I I I R R R R R R R I I I I I V V F L F F F F I S C S H H S D L P P P P P V A E A A A A E T T T T T T Q M M L Q Q M N Q N S T T N E C C C C C C A V I I I I L V K R R K K K G H H H H H H N F Y Y Y Y Y Y I I I I V V I G G G G G G G H H H H H H H G G G G G G G N H H H H Q Q A A A A A C A I I I I I I I N N N N N N N E E E E E E E L L L V V L L K K K K K K K F F F F F F F H H H H H H H P P P P P P P 221 M.nipponense Esc 237 P.humanus corporis Esc
237 S.americana Esc 211 D.plexippus Esc 210 B.mori Esc 221 L.salmonis Esc 220 D.melanogaster Esc R K K R R K H D D D D D D K P P P P P P L N N N N N C Q L V L L L L L L L L L L L L L L L L L L L S S S S S S S V V V A A V G S S S S S S S K K K K K K K D D D D D D D H H H H H H H A A A A A N A L L L L L L I R R R R R R R M L L L L L L W W W W W W W N N N N N N N I I I I I I I R K K M M K Q T T T T T T S D D D D D D H I V V V V H V L C C C C C C V I I I I I I A A A A A A A T I I I I I I F F F F F F L G G G G G G G G G G G G G G V V V V V V V E E E E E E E A G G G G G G H H H H H H H R R R R R R R D D D D D D D E E E E E E E V V V V V V V L L L L L L L 261 M.nipponense Esc 277 P.humanus corporis Esc
277 S.americana Esc 251 D.plexippus Esc 250 B.mori Esc 261 L.salmonis Esc 260 D.melanogaster Esc S S S S S S S A A A A A A I D D D D D D D I F F F F F F D D D D D D N I L L L L R M E K L K K S R G G G G G G G T E E E E E D C K R R R Y R I I I I I I I A M M M M M V S S S S S S S C C C C C C S G G G G G G G M M M M M M M D D D D D D D H H H H H H H S S S S S S S L L L L L L L K K K K K K K I L L L L L L W W W W W W W K R R R R D C L L L L L F L T D D D D N N T K K K K T T D E D P P D P A K S S S H E M M M M M L F S H R N N K H S E E E E K H T V A A A V K I L V I I V I S K R K K K E Q N N Q Q L L S S S S S S S Y Y Y Y Y Y N T S L S N T T F F F F F H F
salmonis and Culex quinquefasciatus,separated from its homologues of flies (Figure 3).
Temporal Expression Pattern and Tissues Distribution of MnEsc
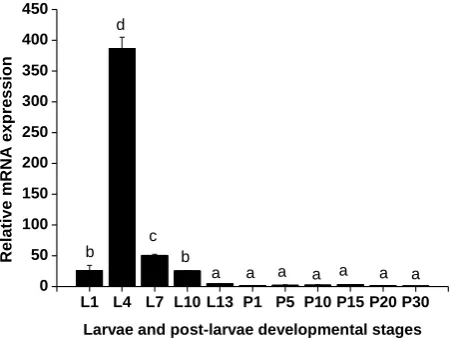
The relative expression levels of MnEsc normalized to β-actin during the developmental stages of embryo and postembryonic were determined by real-time quantitative PCR. During the embryo stages, the expression level of the MnEscwas stronger in the blastula stage (BS) than in the cleavage stage (CS) (P<0.05), and then it abruptly decreased to the lowest level at the gastrul stage (GS). Subsequently, the expression of the MnEsc stayed a low level from the nauplius stage (NS) to zoea stage (ZS) (Figure 4). During the postembryonic, the expression of MnEsc mRNA abruptly increased form 1 day post-hatching larvae (L1) to L4 and peaked at the L4, but it then gradually decreased from the L7 to L13 and reached the lowest level, as well as the one day before the metamorphosis. After metamorphosis, the larvae transition into post-larvae that resemble miniature adults. During the post-larva, it stayed a lowest level (Figure 5).
Real-time quantitative PCR analysis results showed that the relative expressions of MnEsc mRNA was widely distributed in all investigated tissues, including the testis, ovary, brain, abdominal ganglion, heart, intestine, eyestalk and muscle with the highest expression in brain and ovary (Figure 6).
dsRNA Interference Results
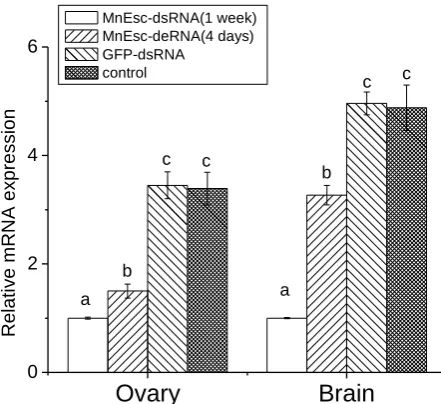
To establish a RNA interference technique in M. nipponense for usage in research of gene function, double strand RNA was systhesized using in vitro transcription. The MnEsc gene was examined by real-time quantitative PCR technique for determining the validity of RNA interference. The MnEsc gene expression was analyzed at different time points, i.e 4 days, 1 week after injection of dsRNA for examining the effect of RNA interference. The results were shown in Figure 7. The gene expression between GFP-dsRNA-injected and vehicle-injected control groups did not differ significantly from one another. After injection for 4 days and 1 week, the decrease of MnEsc expression could be observed in different tissues (ovary and brain) (Figure 7).
301 M.nipponense Esc
317 P.humanus corporis Esc
317 S.americana Esc 291 D.plexippus Esc 290 B.mori Esc 301 L.salmonis Esc 300 D.melanogaster Esc N N N N N N S P A S P P T Q S A A H H Q E R R R R R K K S S S A A L S V N L L L K T R R R R R K L P P P P P N P F F F F F F F P E D N N P P T S S S S T T L H L L L E V Q E K K K L T Q E E E E C K N H H H H H H F F F F F F F P P P P P P P D D D D D L D F F F F F F F S S S S S S S T T T T T T T R R R R R R R D D D D D D D I I I I I I I H H H H H H H R R R R R R R N N N N N N N Y Y Y Y Y Y Y V V V V V V V D D D D D D D C C C C C C C V V V V V C V R R R R R R Q W W W W W W W L I L M M F F G G G G G G G R D D D D N N F F F L L F F I V V I I I V L L L L L L L 339 M.nipponense Esc 355 P.humanus corporis Esc
355 S.americana Esc 329 D.plexippus Esc 328 B.mori Esc 339 L.salmonis Esc 340 D.melanogaster Esc S S S S S S S K K K K K K K S S S S S S S C C C C C C C E E E E E E E N N N N N N N T C C A A T A I I I I I I I V V V I I V V C C C C C C C W W W W W W W K K K K K K K P P P P P P P G G G G G G G L R R R R P Q L L L L L L L N E E E E D H Q D D D D S Q . . . . . . S . . . . . . F T K K T T I E E E E D E S Q L L L L L I V K R K R R K K H N T P P P P N N N G G I S D E D D D N D T T T N N N S N N N S S K S V V V V V V C T T T T T S T I I I I M I I I I I V V I I H H H H H H A K R R R R K E F F F F F F F D E E D D D E Y Y Y Y Y F Y K K R K K K D E E E E E D E 355 M.nipponense Esc 395 P.humanus corporis Esc
395 S.americana Esc 368 D.plexippus Esc 367 B.mori Esc 379 L.salmonis Esc 380 D.melanogaster Esc C C C C C N C E E E E E D E I I I I I I I W W W W W W W F F F F F F F M V V I I V V R R R R R R R F F F F F F F A S A A A S G L M M V V M F D D D D D D N F F F Y Y A P W W W S S D W Q Q Q Q Q Q Q K K K R R N K C I I V V L V . L L I I L I . A A A A A A . L L L L L L . G G G G G G . N N N N N N . Q Q Q Q Q Q . A V C C V Q . G G G G G G . R K K K K K . T T T T T V . F F M M Y Y . V V V V I V . W W W W W W . D D E E D E . L L L L L L . D D G G D D . V V G N V P . P S V V E S . D D A A D D . P P G G P P . N S G G S E . L Q . . S G . A S S S T A . K R R R K H 355 M.nipponense Esc 435 P.humanus corporis Esc
435 S.americana Esc 408 D.plexippus Esc 407 B.mori Esc 419 L.salmonis Esc 420 D.melanogaster Esc . C C V V F M . I T S S T T . T A L Q V T . L L L L L L . S T V V S H . H H H H H N . P P P P P S . K R R R K R . C C C C C S . T V V V N V . S A A A V A . A A A A A T . V I V V I V . R R R R R R . Q Q Q Q Q Q . T T V V T I . S S T T S A . L L L L F F . S S S S S S . R R R R K R . D D N N G D . G G G G G A . S S K K D S . L V I I I V . L L L L C L . V L L L I V . C C T T C Y . V V C C G V . C C C C C C . D D D D D D . D D D D D D . G G G G G A . T T T T T T . I I I I I V . W W W W W W . R R R R R R . W W W W W W . D D D D D N . R R R R R R . V I V V Q R 355 M.nipponense Esc 437 P.humanus corporis Esc
437 S.americana Esc 412 D.plexippus Esc 411 B.mori Esc 422 L.salmonis Esc 425 D.melanogaster Esc . L P H S N Q . T V N A S T . . . G S V T . . . S N . S . . . . . . I
Schistocerca americana
Tribolium castaneum Pediculus humanus corporis
Danaus plexippus Bombyx mori
Junonia coenia
Culex quinquefasciatus
Macrobrachium nipponense Lepeophtheirus salmonis
Glossina morsitans morsitans Musca domestica
Drosophila melanogaster Drosophila virilis
100 99
100 85 100 83
80 92
89 57
0.05
Figure 3. A phylogenetic tree was constructed based on the comparisons of amino acid sequences. Species names are listed on the right of the tree. Their accession number in GenBank as followed: Pediculus humanus corporis (XP_002427573.1), Schistocerca Americana (AAC05332.1), Danaus plexippus (EHJ72379.1), Glossina morsitans morsitans (ADD18468.1), Bombyx mori (NP_001188366.1, Drosophila melanogaster (NP_477431.1), Drosophila virilis (XP_002052298.1), Junonia coenia (AAC05331.1), Culex quinquefasciatusand (XP_001842089.1), Tribolium castaneum (XP_973780.1), Musca domestica (AAC05333.1), Lepeophtheirus salmonis (ACO12729.1).
C S B S G S N S P S Z S 0
5 0 1 0 0 1 5 0 2 0 0 2 5 0
R
e
la
ti
v
e
m
R
N
A
e
x
p
r
e
s
s
io
n
E m b r y o d e v e lo p m e n t a l s t a g e s a b a a c
d
Figure 4. The expression profile of MnEsc at the different development stages of embryos were revealed by real-time quantitative PCR. The amount of MnEsc mRNA was normalized to the β-actin transcript level. Data are shown as means ± SE of three repeated samples during the embryos, larvae and post-larvae. CS-cleavage stage; BS-blastula stage; GS-gastrula stage; NS-nauplius stage; PS-protozoea stage; ZS-zoea stage.
L1 L4 L7 L10 L13 P1 P5 P10 P15 P20 P30 0
50 100 150 200 250 300 350 400 450
Relat
ive
m
R
NA
exp
ressi
o
n
Larvae and post-larvae developmental stages
a a a a a a a
b b
c d
Discussion
In the present study, we have successfully cloned the full-length cDNA of Escgene based on the EST sequence from a M. nipponense testis cDNA library. The cDNA sequence encoded a protein of 355 amino acids having a molecular weight of 40.201 kDa. From the result of alignment, this deduced amino acid sequence shares high homology with those previously reported arthropoda Esc proteins, which confirmed the high conservation of this protein. The insect extra sex combs protein contains multiple WD repeats (Gutjahr et al., 1995; Sathe et al., 1995; Simon et al., 1995), which was essential for its function as a repressor of homeotic genes (Sathe et al., 1995). Conserved sequence of five WD40 repeated motifs were identified in the deduced amino acid sequences of Esc (van der Voorn and Ploegh,
1992; Neer et al., 1994). The widespread distribution of the WD40 repeats throughout MnEsc protein implies that its function is mainly by contacting other proteins (Gutjahr et al., 1995; Komachi et al., 1994; Whiteway et al., 1994).
Previous studies indicated that Esc product was required only in the very early stages of embryonic development (Gutjahr et al., 1995; Simon et al., 1995; Sathe et al., 1995). In the present study, expression of Esc mRNA increased at the early embryo, and then drop to a low level during larval growth folloeing increasing again during pupal stages, which is consistent with a transient role in establishing stable long term PcG-mediated repression (Sathe et al., 1995; Katsuhito et al., 2008). Our study was the first to report the involvement of Esc in the development of embryo and postembryonic and in other tissues of M. nipponense. Real-time quantitative PCR analysis
T O AG B H In Et M
0 10 20 30 40
Re
la
tiv
e
m
RN
A
e
x
p
re
s
s
io
n
Different tissues in mature prawns
a a
a
b b
c
d d
Figure 6. The expression profile of MnEsc in different tissues was revealed by real-time quantitative PCR. The amount of Esc mRNA was normalized to the β-actin transcript level. Data are shown as means ± SE of three replicates in various tissues. T-testis; O-ovary; B-brain; AG-abdominal ganglion; H-heart; In-intestine; Et-Eyestalk; M-muscle.
Ovary
Brain
0 2 4 6
Rela
ti
ve
m
RN
A
e
xp
re
ssio
n
MnEsc-dsRNA(1 week) MnEsc-deRNA(4 days) GFP-dsRNA
control
a a
b
b
c c
c c
indicated that MnEsc is mainly expressed in the blastula stage of embryonic and the fourth larvae, which was similar with previous studies in the early developmental embryos of Drosophila (Gutjahr et al., 1995) and medaka (Norihisa et al., 2004). These data indicated that the MnEsc might be related to the embryogenesis and organogenesis of the M. nipponense. The Esc gene regulated the body segment and controlled the terminal pattern of leg sex comb differentiation of male Drosophila (Chiyoko et al., 1965; Papaceit et al., 1991; Struh et al., 1982). During the postembryonic stage of M. nipponense, we found that the expression of MnEsc displayed a transient increasing in 4 days post-hatching larvae, and then its expression in the post-larvae did not change significantly after metamorphosis. In a follow up study (unpublished), the eyestalk and branchiostegal spine of M. nipponense are developed from 4 days post-hatching larvae to 11 days post-hatching larvae according to morphological observation. Therefore, we speculated that the expression pattern of MnEsc during postembryonic development may concern to pre-metamorphic larval changes of morphology and the organogenesis of larvae.
The genetic evidence indicates that the maternal Esc contribution deposited in the oocyte is critical for early embryonic development (Struhl, 1981). At very late stages of Drosophila embryo, Esc is again expressed zygotically in specific regions of the brain of Drosophila (Gutjahr et al., 1995). The results of present study showed that MnEsc mRNA expressed in all detected tissues. The highest level is observed in brain and ovary of the oriental river prawn. These data indicate that the MnEsc mRNA may be also maternally deposited in the oocyte, its biological function may be controlled by central nervous system. RNAi experiment showed that the decrease of MnEsc mRNA expression could be observed in different tissues after injection of MnEsc-dsRNA for four days and 1 week, indicating that RNA interference could be induced systemically and the interference effect could be transported among different tissues. But no exterior appearance change of experimental shrimps was observed. This may due to two reasons: firstly, the time is not long enough from interference to the observation, the M. nipponense have not been molted to change the phenotype. Secondly, experimental shrimps are mature, their phenotype is not easy to change after interference.
In conclusion, this study is the first report of an Esc gene in the oriental river prawn, M. nipponense. MnEsc may play important roles in the embryogenesis, oogenesis and morphological differentiation of the early larva, and may be used as a molecular marker for future studies of M. nipponense embryonic development. Moreover, a stable RNA interference technology could provide an effective molecular biological approach for the identification of gene function and antiviral research in M. nipponense.
Acknowledgements
The project was supported by the National Natural Science Foundation of China (Grant No.31272654), the National Science & Technology Supporting Program of the 12th Five-year Plan of China (Grant No. 2012BAD26B04), the Jiangsu Provincial Natural Science Foundation for Young Scholars of China (Grant No. BK2012091) and the Science & Technology Supporting Program of Jiangsu Province (Grant No. BE2012334).
References
Altschul, S. F., Gish, W., Miller, W., Myers, E. W. and Lipman, D. J. 1990. Basic local alignment search tool. Journal of Molecular Biology, 215: 403-410. doi: 10.1016/S0022-2836(05)80360-2.
Bracken, A. P. and Helin, K. 2009. Polycomb group proteins: navigators of lineage pathways led astray in cancer. Nature Reviews Cancer, 9(11):773-784. doi: 10.1038/nrc2736.
Breen, T. R. and Duncan, I. M. 1986. Maternal expression of genes that regulate the bithorax complex of Drosophila melanogaster. Developmental Biology, 118: 442-456. doi: 10.1016/0012-1606(86)90015-1. Budd, G. E. and Telford, M. J. 2009. The origin and
evolution of arthropods. Nature, 457: 812-817. doi: 10.1038/nature07890.
Bureau of Fishery, Ministry of Agriculture, P.R.C. 2009. Fisheries economic statistics. In: China Fishery Yearbook. China Agricultural Press, Beijing, 236pp. Chen, Y., Zhu, Q., Chen, H., Zhu, X. L., Cui, Z. and Qiu, G.
F. 2012. The morphological and histological observation of embryonic development in the oriental river prawn Macrobrachium nipponense. Journal of shanghai ocean university (China), 21(1): 33-40. Chiyoko, T. and Curt, S. 1965. The developmental
autonomy of extra sex combs in Drosophila melanogaster. Developmental Biology, 11(1): 50-81. doi:10.1016/0012-1606(65)90037-0
Duncan, I. M. 1982. Polycomblike: a gene that appears to be required for the normal expression of the bithorax and Antennapedia complexes of Drosophila melanogaster. Genetics, 102: 49-70. http://www.genetics.org/content/102/1/49.
Dura, J. M., Brock, H. W. and Santamaria, P. 1985. Polyhomeotic: a gene in Drosophila meIanogaster required for correct expression of segment identity. Molecular and General Genetics, 198: 220-231. doi:10.1007/BF00382998.
Faust, C., Lawson, K. A., Schork, N. J., Thiel, B. and Magnuson, T. 1998. The Polycomb-group gene eed is required for normal morphogenetic movements during gastrulation in the mouse embryo. Development, 125:4495–4506.
http://dev.biologists.org/content/125/22/4495. Girton, J. R. and Jeon, S. H. 1994. Novel embryonic and
adult homeotic phenotypes are produced by pieiohomeotic mutations in Drosophila. Developmental Biology, 161:393-407. doi:org/10.1006/dbio.1994.1040.
WD-40 repeat family. EMBO Journal, 14: 4296– 4306. PMCID: PMC394514
Ingham, P. 1984. A gene that regulates the bithorax complex differentially in larval and adult cells of Drosophila. Cell, 37: 815-823. doi: 10.1016/0092-8674(84)90416-1.
Jones, R. S. and Gelbart, W. M. 1990. Genetic analysis of the Enhancer of zeste locus and its role in gene regulation in Drosophila melanogaster. Genetics, 126:185-199. PMCID: PMC1204123.
Joyce, N. G., Li, R. H., Kelly, M. and Simon, J. 1997. Evolutionary Conservation and Predicted Structure of the Drosophila extra sex combs Repressor Protein. Molecular and cellular biology, 17(11): 6663–6672. doi:0270-7306/97/$04.0010
JÜrgens, G. 1985. A group of genes controlling the spatial expression of the bithorax complex in Drosophila. Nature, 316:153-155. doi:10.1038/316153a0.
Katsuhito, O., Donna, M., Birgit, C., Axel, I. and Vincenzo, P. 2008. ESC, ESCL and their roles in Polycomb Group mechanisms, mechanisms of development, 125: 527–541. doi:10.1016/j.mod.2008.01.002. Kohler, C. and Hennig, L. 2010. Regulation of cell identity
by plant Polycomb and trithorax group proteins. Current Opinion in Genetics & Devlopment, 20(5):541-547. doi: org/10.1016/j.gde.2010.04.015. Komachi, K., Redd, M. J. and Johnson, A. D. 1994. The
WD repeats of Tup1 interact with the homeo domain protein alpha 2. Genes and Development, 8(23): 2857-2867.doi:10.1101/gad.8.23.2857.
Lewis, E.B. 1978. A gene complex controlling segmentation in Drosophila. Nature, 276: 565-570. PMID: 103000.
Li, Z. Q., Tsuneyuki, T., Kosuke, S., Zhu, L., Xu, J., Hiroaki, M., Jae, M. L. and Takahiro, K. 2012. Identification and characterization of Polycomb group genes in the silkworm, Bombyx mori. Molecular Biology Reports, 39:5575-5588. doi: 10.1007/s11033-011-1362-5
Livak, K. J. and Schmittgen, T. D. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2 (−Delta Delta C (T))
Method. Methods, 25:402–408.
doi:org/10.1006/meth.2001.1262.
Morey, L. and Helin, K. 2010. Polycomb group protein-mediated repression of transcription. Trends in Biochemical Sciences, 35(6):323-332. doi: 10.1016/j.tibs.2010.02.009.
Neer, E. J., Schmidt, C. J., Nambudripad, R. and Smith, T. F. 1994. The ancient regulatory-protein family of WD-repeat proteins. Nature, 371: 297-300. doi: 10.1038/371297a0.
Norihisa, S., Atsushi, S., Kouji, Y. and Toru, H. 2004. Participation of Polycomb group gene extra sex combs in hedgehog signaling pathway. Biochemical and Biophysical Research Communications, 323: 523-533. doi:10.1016/j.bbrc.2004.08.125
Oktaba, K., Gutierrez, L., Gagneur, J., Girardot, C., Sengupta, A. K., Furlong, E. E. and Muller, J. 2008. Dynamic regulation by Polycomb group protein complexes controls pattern formation and the cell cycle in Drosophila. Developmental Cell, 15(6):
877-889.doi:10.1016/j.devcel.2008.10.005.
Papaceit, M., Antonio, J. S. and Prevosti, A. 1991. Genetic analysis of extra sex combs in the hybrids between Drosophila subobscura and D. madeirensis. Genetics, 84: 107-114.doi: 10.1007/BF00116550.
Qiao, H., Fu, H. T., Jin, S. B., Wu, Y., Jiang, S. F., Gong, Y. S. and Xiong, Y. W. 2012. Constructing and random sequencing analysis of normalized cDNA library of testis tissue from oriental river prawn (Macrobrachium nipponense).Comparative Biochemistry and Physiology D-genomics & proteomics, 7(3): 268-276. doi:10.1016/j.cbd. 2012.04.003.
Sathe, S. S. and Harte, P. J. 1995. The Drosophila extra sex combs protein contains WD motifs essential for its function as a repressor of homeotic genes. Mechanisms of Development, 52:77–87.
doi:org/10.1016/0925-4773(95)00392-E.
Sato, T., Hayes, P. H. and Denell, R. E. 1983. Homeosis in Drosophila: maternal effect of the Enhancer of Polycomb locus and its interaction with Polycomb and related loci. Developmental Genetics, 4:185-198. doi: 10.1002/dvg.1020040305
Simon, J., Bornemann, D., Lunde, K. and Schwartz, C. 1995. The extra sex combs product contains WD40 repeats and its time of action implies a role distinct from other Polycomb group products. Mechanisms of Development, 53: 197-208. doi:org/10.1016/0925-4773(95)00434-3.
Struhl, G. 1981. A gene product required for correct initiation of segmental determination in Drosophila. Nature, 293: 36-41. doi:10.1038/293036a0.
Struhl, G. and Akam, M. 1985. Altered distributions of Ultrabithorax transcripts in extra sex combs mutant embryos of Drosophila. EMBO Journal, 4: 3259-3264. PMCID: PMC554652.
Struhl, G. and Brower, D. 1982. Early role of the ESC1 gene product in the determination of segments in Drosophila. Cell, 31: 285-292. doi:10.1016/0092-8674(82)90428-7.
Tamura, K., Dudley, J., Nei, M. and Kumar, S. 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Molecular Biology and Evolution 24: 1596-1599. doi: 10.1093/molbev/ msm092.
van der Voorn, L. and Ploegh, H. L. 1992. The WD-40 repeat. Febs Letters, 307: 131-134. doi: 10.1016/0014-5793(92)80751-2.
Wang, Q., Fang, D.A., Sun, J.L., Wang, Y., Wang, J. and Liu, L.H. 2012. Characterization of the vasa gene in the Chinese mitten crab Eriocheir sinensis: A germ line molecular marker. Journal of Insect Physiology, 58:960–965. doi: 10.1016/j.jinsphys.2012.04.012 Whiteway, M., Clark, K.L., Leberer, E., Dignard, D. and
Thomas, D.Y. 1994. Genetic identification of residues involved in association of α and β G-protein subunits. Molecular and Cellular Biology, 14:3223-3229. doi: 10.1128/MCB.14.5.3223.