Copyright © 2004, American Society for Microbiology. All Rights Reserved.
Sequence Elements Necessary for Transcriptional Activation of
BAD1
in the Yeast Phase of
Blastomyces dermatitidis
Peggy J. Rooney
1and Bruce S. Klein
1,2,3*
Department of Medical Microbiology and Immunology,1Department of Pediatrics,2and Department of Internal Medicine
and the Comprehensive Cancer Center,3University of Wisconsin Medical School, Madison, Wisconsin 53792
Received 27 February 2004/Accepted 2 April 2004
Blastomyces dermatitidisis a dimorphic fungal pathogen that converts from mycelia or conidia to a host-adapted yeast morphotype upon infection. Conversion to the yeast form is accompanied by the production of the virulence factor BAD1. Yeast-phase-specific expression of BAD1 is transcriptionally regulated, and its
promoter shares homology with that of the yeast-phase-specific geneYPS3ofHistoplasma capsulatum. Serial
truncations of the BAD1upstream region were fused to thelacZreporter to define functional areas in the
promoter. Examination of PBAD1-lacZfusions inB. dermatitidisindicated thatBAD1transcription is
upregu-lated in the yeast phase. The 63-nucleotide box A region conserved in theYPS3upstream region was shown to
be an essential component of the minimalBAD1promoter. A matched PYPS3-lacZconstruct indicated that this
same region was needed for minimalYPS3promoter activity inB. dermatitidistransformants. Reporter activity
inH. capsulatumtransformants similarly showed a requirement for box A in the minimalBAD1promoter.
Several putative transcription factor binding sites were identified within box A ofBAD1. Replacement of two
of these predicted sites within box A—a cAMP responsive element and a Myb binding site—sharply reduced transcriptional activity, indicating that these regions are critical in dictating the yeast-phase-specific
expres-sion of this crucial virulence determinant ofB. dermatitidis.
Most systemic fungal infections of immunocompetent hu-mans are caused by dimorphic fungi. This group of pathogens includes Blastomyces dermatitidis, Histoplasma capsulatum,
Paracoccidioides brasiliensis, Coccidioides immitis, and
Sporo-thrix schenkii. These fungi share the unique ability to undergo
reversible cellular differentiation, existing as an infectious form or morphotype in the environment but converting to a second, pathogenic form upon entry into the host. This morphological switch promotes host invasion and evasion (40). A change in fungal gene expression is one possible mechanism by which morphogenesis affects the outcome of infection. Factors re-quired for colonization of the host or modulation of the host immune response may be upregulated, while other factors only necessary for growth in the environment may be downregu-lated. Identification of products whose expression is linked only to the pathogenic phase may uncover virulence factors of these fungi. Also, since prevention of this essential develop-mental event may hold the key to control of infections by these fungi, identification of molecules necessary for the morpholog-ical switch could lead to new drug or vaccine targets that block the earliest events in colonization or infection.
Several genes have been identified whose expression is spe-cific to one morphotype, a subset of which have been shown to be required for virulence. For example, expression of SOWgp58, an extracellular matrix-binding protein ofC. immi-tis, is limited to the endosporulating spherule form, which is the parasitic phase of this fungus (22). Deletion of SOWgp
resulted in a reduction in virulence, and its product is pre-sumed to play a role as an adhesin (23). In H. capsulatum,
expression of the calcium-binding protein CBP1 was limited to the yeast phase, and a knockout strain lacking CBP1 was un-able to destroy macrophages in vitro or colonize in a mouse model of pulmonary histoplasmosis (45).BAD1ofB.
dermati-tidis, like CBP1, was also shown to be a yeast-phase-specific
gene and a virulence factor (41, 12).BYS1ofB. dermatitidisis a yeast-phase-specific gene of unknown function that contains 3⬘signals found in message with regulated stability (13), but no such signals were found in the 3⬘untranslated region ofBAD1
(unpublished observation). Promoter analysis of CBP1 re-vealed that this gene is transcriptionally upregulated in the yeast and identified a stretch of DNA required for yeast-phase expression of this gene (35).
B. dermatitidisexists as a mold in the acidic soil found along
the Mississippi and Ohio River valleys. Upon inhalation of conidia or mycelial fragments, this fungus converts to the yeast morphotype and colonizes the host alveoli (10). Infection can lead to an acute pulmonary disease or can disseminate to other sites, most notably bone and skin. Conversion to the yeast form induces the expression of BAD1, which becomes arrayed on the surface of the yeast and is released into the extracellular milieu (27, 41). A knockout strain lacking BAD1 was avirulent in a mouse model of infection, while a strain in which BAD1 expression was reconstituted was restored to virulence match-ing that of the parental strain (12). BAD1 has been shown to bind to complement receptor 3 and may serve as an adhesin, although recently it was shown that this property is dispensable for virulence (31, 11). Both cell-associated and secreted forms of the protein modulate the host inflammatory response in favor of the pathogen by downregulating production of the proinflammatory cytokine tumor necrosis factor alpha (17, 18). Expression of BAD1 is limited to the yeast phase and reg-ulated transcriptionally. Analysis of the promoter region
iden-* Corresponding author. Mailing address: University of Wisconsin, 600 Highland Ave., K4/434, Madison, WI 53792. Phone: (608) 263-9217. Fax: (608) 263-6210. E-mail: [email protected].
785
on September 8, 2020 by guest
http://ec.asm.org/
tified two regions of homology with that of another yeast-phase-specific gene,YPS3ofH. capsulatum(26). These regions were termed boxes B and A and shared 86 and 80% identity between theBAD1gene ofB. dermatitidisstrain 26199 and the
YPS3gene of H. capsulatumstrain G217B, respectively (see Fig. 1). An additional stretch of nucleotides upstream of boxes B and A in theYPS3upstream region, which is able to bind protein present in a yeast nuclear extract via electrophoretic mobility shift assay (24), was also identified in theBAD1 up-stream region. When theBAD1promoter was expressed het-erologously inH. capsulatum, its transcription was phase reg-ulated, indicating that the regulatory machinery is conserved between these two fungi (41).
Herein, we explored the sequences required for regulating expression of BAD1. Serial truncations of upstream sequences fused to a -galactosidase reporter were used to functionally analyze homologous regions of theBAD1andYPS3promoters in bothB. dermatitidisand H. capsulatum. We observed that the conserved box A region is an essential component of the minimalBAD1promoter. This same region is needed for min-imalYPS3promoter activity inB. dermatitidis, whereas addi-tional upstream sequence 5⬘ to box A is required for YPS3
transcription inH. capsulatum. We also pinpointed two puta-tive transcription factor binding sites within box A that are essential for upregulatingBAD1expression in the yeast phase.
MATERIALS AND METHODS
Strains and growth.B. dermatitidisstrain 26199 andSaccharomyces cerevisiae strain 204509 were obtained from the American Type Culture Collection
(Rock-ville, Md.).H. capsulatumG217Bura5-23(39) was provided by J. Woods (Uni-versity of Wisconsin).Agrobacterium tumefaciensstrain LBA1100 (4) was pro-vided by C. van den Hondel (Leiden University, Leiden, The Netherlands) and grown at 28°C on Luria broth supplemented with 0.1% glucose.S. cerevisiaewas grown on yeast extract-peptone-dextrose at 28°C.B. dermatitidisyeast was main-tained on Middlebrook 7H10 medium containing oleic acid-albumin complex prior to transformation. For reporter assays,B. dermatitidiswas grown as yeast at 37°C or as mycelia at 22°C inHistoplasmamacrophage medium (HMM) (58). Conversion from yeast to mold was accomplished by transferring log-phase yeast cultures to 22 to 25°C. For studies of mycelia, conversion was allowed to proceed for approximately 8 weeks of growth at 22 to 25°C on an orbital shaker.H. capsulatumwas grown on an orbital shaker in HMM or HMM supplemented with 100g of uracil/ml as yeast at 37°C in 5% CO2.
Plasmid constructions.A base vector to allow fusion of promoter fragments to theE. coli lacZcoding region was made by deletion of the BamHI site down-stream oflacZto create pWLT⌬3⬘Bam. Promoter fragments were amplified fromBAD1 andYPS3using the vectors pWLT (41) and pY1 (YPS3of H. capsulatumstrain G217B in the vector pGEM-Teasy, provided by J. Woods, University of Wisconsin) as templates, respectively. A fragment of theS. cerevi-siae ACT1locus, representing bases 1940 to 2002 (GenBank accession number L00026.1), was amplified from genomic DNA. TheURA5promoter ofH. cap-sulatumstrain G184AS (from⫺311 to⫹87) was amplified from pWU20 (57), using primers N-XURAFOR (5⬘-ATGCGGCCGCTCTAGAAAGCTTTATAT AAACTTGAGAG) and URABAMREV (5⬘-TAGGATCCATTGTAGCAATT CCCGCTGGGTC). Amplification ofBAD1promoter fragments was performed using forward primers in Table 1, with added 5⬘NotI and XbaI sites (5⬘-AAG CGGCCGCTCTAGA-) and reverse primer DELTAREV (5⬘-TAGGATCCAT TCTCCCGGTAGGTAGCTC). Amplification ofYPS3promoter fragments was performed using the forward primers in Table 1, including 5⬘NotI, XbaI, BsrGI, and StuI sites (5⬘-AAGCGGCCGCTCTAGATGTACAAGGCCT-) and reverse primer YPSREV (5⬘-TAGGATCCATTTTTGCGGTTGATTATTT). Modified BAD1andURA5promoter fragments were created by amplification, either via splicing by overlap extension (SOE) or primer-induced sequence alterations (Table 2). Creation of the ⫺741 A⌬::ACT1 and B ivs A⌬ACT1 promoter modifications required two-step SOE, in which two products were fused, and these fusion products were then spliced to a third fragment prior to cloning. Amplifications were performed using Elongase (Invitrogen Corp., Carlsbad, Calif.), and the primers used are listed in Table 2. Those creating the 5⬘end of the promoter included additional 5⬘NotI and XbaI sites.
PCR products were digested with NotI and BamHI and cloned into pWLT⌬3⬘Bam in replacement of the large (⬃2.7-kb)BAD1upstream region, and their sequences were confirmed. The 2.5-kbBAD1upstream region fused to lacZin T-DNA vector pBTS92 (41) was modified to receive promoter fragments. The ClaI site outside of thelacZcoding region, and the XbaI site not immedi-ately 5⬘of theBAD1promoter, were removed to create vector pBPR25. Pro-moters and a fused 5⬘ fragment oflacZwere removed by digestion of the
FIG. 1. Serial truncations of the upstream regions of BAD1and
YPS3. Shown are schematic representations of amplified fragments that were fused tolacZto assess the contributions of promoter regions toB. dermatitidis(Bd)BAD1andH. capsulatum(Hc)YPS3 transcrip-tion (A and B, respectively). Transcriptranscrip-tional start sites are indicated by arrows. Numbering of the 5⬘and 3⬘(at the translation-initiating ATG) ends of the promoter fragments are indicated relative to the transcrip-tional start sites (⫹1). At the right are promoter fragment designa-tions, based on either the distance upstream of the transcriptional start site or homology between theBAD1andYPS3promoters at boxes B and A, shown as shaded boxes.
TABLE 1. Primers for amplification ofBAD1andYPS3promoter truncation fragments
Gene Promotera Forward primer sequenceb
BAD1 ⫺1101 ACCCTGGTCGAATATTCCAGA
BAD1 ⫺741 GAGAGTACGGGTCGTATATTT
BAD1 B ivs A CAGATGTTTCGATAAGAGGCC
BAD1 ivs-box A TACTACCCCTATCCGATAATG
BAD1 box A ATTGTGTGGATAGACGTCATA
BAD1 No BA AGGTATCAGCTAGCCGGCTTG
BAD1 No TATA GCTCTCGTCTTGCTCCTTTTT
BAD1 No Start CGCCCAGTCATTCGTTCATTT
YPS3 ⫺741 AAGCTTTATTCAGCCGCAAGA
YPS3 B ivs A TAGATGTTTCGATAAGTCAAG
YPS3 ivs-box A ACAGCGCAGTGATGCAATAAT
YPS3 box A GTTGTTTAGACAGACGTCATA
YPS3 1/2 box A TTGACTCTTGTCCATGTTTCT
YPS3 No BA TAGCATCAGTTAGCCTGCCCA
aNomenclature based on areas of homology between the two promoters contained within the PCR product or the 5⬘end of PCR product numbered relative to the transcriptional start site.
bPrimers had 5⬘extensions (see Materials and Methods). TheYPS3⫺741 product was obtained by binding to a site containing a partial primer match.
on September 8, 2020 by guest
http://ec.asm.org/
pWLT⌬3⬘Bam-based vectors with XbaI and ClaI and cloned into theE. coli-A. tumefaciens-B. dermatitidisshuttle vector pBPR25.
Construction ofH. capsulatumtelomeric vectors necessitated reamplification ofBAD1promoter fragments using the forward primers listed in Table 1, with additional 5⬘NotI, XbaI, BsrGI, and StuI sites (see above) and reverse primer DELTAREV. PCR products were cloned into the NotI and BamHI sites of pWLT⌬3⬘Bam and confirmed by sequencing. To prevent read-through transcrip-tion previously noted in telomeric reporter constructs inH. capsulatum(data not shown), the transcriptional terminator region from theAspergillus nidulans trpC gene was amplified from pAN7-1 (37) with primers B-TRPCF (5⬘-AATGTAC AACTTAACGTTACTGAAATCATC) and S-TRPCR (5⬘-AAAGGCCTAAG AAGGATTACCTCTAAACA), cloned and sequenced (pPJ56), and subcloned into the BsrGI-StuI sites upstream of theBAD1andYPS3promoters fused to lacZ. Generation of a vector for selection inH. capsulatumbegan with deletion of a PmeI fragment from pWU56, an EcoRI deletion derivative of pWU55 (57) (provided by J. Woods, University of Wisconsin). ThePodospora anserina URA5 (PaURA5) marker from pWU55 was added into the EcoRI site, and a plasmid in which the marker would be transcribed towards the nearest PmeI site was selected for further modification (pPJ11). A kanamycin resistance (kanr) marker
amplified from pBIN19 (5) using primers ESKAN-U (5⬘-GATATCACTAGTC GATAAACCCAGCGAACCATT) and ESKAN-L (5⬘-GATATCACTAGTCA TCTAGGTACTAAAACAATT), with EcoRV and SpeI sites added at each end, was cloned into the EcoRV site to form pPJ11Kan. Terminator-promoter-lacZ -containing fragments were removed by digestion with XbaI and cloned into the SpeI site of pPJ11Kan in replacement of the kanrcassette. Clones were selected
in whichlacZwould be transcribed opposite thePaURA5selectable marker. Telomeric repeats able to be exposed by PmeI digestion were isolated from pWU55 and cloned into the NotI site of eachH. capsulatum-ready reporter vector.
Fungal transformation.Transformation ofB. dermatitidisusingA. tumefaciens strain LBA1100 derivatives was performed as previously described (41, 48). Transformants were selected on HMM containing 100 g of hygromycin B/ml. As the resulting plates were crowded with dense growth, transformant colonies were first patched and then streaked out for single-colony isolation on HMM containing 100g of hygromycin B/ml prior to inoculating liquid cultures for reporter analysis. This method of DNA delivery results in random insertion in the genome (48). Since the position of insertion can affect transgene expression, to allow comparison of transformant promoter activity among the vectors, eight transformants of each deletion vector were analyzed for-galactosidase (LacZ) activity and the results were averaged. To reduce the effects of passage number, constructs to be compared were transformed simultaneously into the same recipient culture.H. capsulatumtransformation with PmeI linearized plasmid DNA was accomplished using electroporation conditions described previously (57). Transformants were selected on HMM at 37°C with 5% CO2.
DNA analysis.Alignment and sequence comparison of theBAD1andYPS3 upstream regions was performed using ClustalW, v1.4, of the MacVector nucle-otide analysis software package (Accelrys, Burlington, Mass.). Determinations of sequence identity were performed using short sequence files of the regions being compared. The reported percentage identity between these promoters at box B and the region containing a probe used in gel shift assays (24) required use of sequence files containing additional sequence beyond the alignment of these regions highlighted in Fig. 1. This was necessary to eliminate gaps created at the ends of these regions when the approximately 1.1 kb of upstream sequence was aligned.
As a test of reporter transfer, selectedB. dermatitidistransformants were analyzed by Southern hybridization. Genomic DNA fromB. dermatitidiswas isolated using a liquid nitrogen grinding technique for cellular disruption,
fol-TABLE 2. Primers for amplification ofBAD1andYPS3modified promoter fragments
Promotera Primersb
⫺741 A⌬::ACT1...GAGAGTACGGGTCGTATATTT*/
GAAAGAAAGTACTCCGTCTGGATGTGCCGAATCAATAATGGAC; GTCCATTATTGATTCGGCACATCCAGACGGAGTACTTTCTTTC/ CAAGCCGGCTAGCTGATACCTATGGCTCCATCTTCCATGAAGGT; ACCTTCATGGAAGATGGAGCCATAGGTATCAGCTAGCCGGGCTTG/ TAGGATCCATTCTCCCGGTAGGTAGCTC
B ivs A⌬::ACT1...CAGATGTTTCGATAAGAGGCC*/
GAAAGAAAGTACTCCGTCTGGATGTGCCGAATCAATAATGGAC; GTCCATTATTGATTCGGCACATCCAGACGGAGTACTTTCTTTC/ CAAGCCGGCTAGCTGATACCTATGGCTCCATCTTCCATGAAGGT; ACCTTCATGGAAGATGGAGCCATAGGTATCAGCTAGCCGGGCTTG/ TAGGATCCATTCTCCCGGTAGGTAGCTC
Box A-PURA5...TTGTGTGGATAGACGTCATATATTGTTCCGTTTTGGCTTA*/ CATCACGGAGGGCATTTCCATCAACAGAAACATTGGCAAGAG; CTCTTGCCAATGTTTCTGTTGATGGAAATGCCCTCCGTGATG/ TAGGATCCATTGTAGCAATTCCCGCTGGGTC
ACT1-PURA5...TCCAGACGGAGTACTTTCTTTC*/
CATCACGGAGGGCATTTCCATTGGCTCCATCTTCCATGAAGGT; ACCTTCATGGAAGATGGAGCCAATGGAAATGCCCTCCGTGATG/ TAGGATCCATTGTAGCAATTCCCGCTGGGTC
A-CRE⌬::sp...TTGTGTGGAGACTACGTGCATATTGTTCCGTTTTGGCTTA*
A-Sox-5⌬::sp...TTGTGTGGATAGACGTCATAGACTACGTGCTTTTGGCTTAGGCTTCTTGCCA* A-Myb⌬::sp...TTGTGTGGATAGACGTCATATATTGACTACGTGCGGCTTAGGCTCTTGCCAATGT* A-NF-1⌬::sp ...TTGTGTGGATAGACGTCATATATTGTTCCGTTTTGGCTTA*/
ACAACAGAAACGCACGTAGTCGCCTAAGCCAAAACGGAACAA; TTGTTCCGTTTTGGCTTAGGCGACTACGTGCGTTTCTGTTGT/ TAGGATCCATTCTCCCGGTAGGTAGCTC
A-AREB6⌬::sp...TTGTGTGGATAGACGTCATATATTGTTCCGTTTTGGCTTA*/ ACAGCACGTAGTCTGGCAAGAGCCTAAGCCAAAAC; GTTTTGGCTTAGGCTCTTGCCAGACTACGTGCTGT/ TAGGATCCATTCTCCCGGTAGGTAGCTC
A-CRE/Myb⌬::sp...TTGTGTGGAGACTACGTGCATATTGACTACGTGCGGCTTAGGCTCTTGC*
aAll vectors contain modified P
BAD1fragments, unless otherwise indicated. Replacements of predicted transcription factor binding sites are indicated by the site (CRE) or the protein (Sox-5, Myb, NF-1, and AREB6) that may bind it.ACT1represents a 63-bp fragment of theS. cerevisiaeactin locus (see Materials and Methods). Sp, 10-bp spacer.
bPrimers indicated by an asterisk had 5⬘extensions (see Materials and Methods). Paired SOE primers are separated by a slash, and sets used together to generate the final promoter fragment are separated by a semicolon. The 10-base replacement sequences are underlined.
on September 8, 2020 by guest
http://ec.asm.org/
lowed by organic extraction (21). For each sample, 2.5g of genomic DNA was separately digested with EcoRI, isolating a fragment containing the PBAD1-or PYPS3-lacZfusion, and XbaI, which digests the T-DNA reporter vectors at a single site. Transforming plasmid DNAs digested with the same enzymes were included as controls. DNA analysis by Southern blot hybridization was per-formed using standard techniques (43). Blots were probed forlacZusing a BamHI fragment of pWLT (41). Probes for hybridizations were gel purified using the QIAquick gel extraction kit (QIAGEN, Inc., Valencia, Calif.) and radiolabeled using the Prime-a-Gene labeling system (Promega Corp., Madison, Wis.). Four transformants each for plasmids containing the No BA-lacZand No Start-lacZreporters (Table 1) were tested in this fashion to ensure that their relatively low level of-galactosidase (LacZ) activity was not due to rearrange-ment of the promoter-reporter fusion. Results indicated that the fate of trans-forming DNA was integration at a single site, with multimerization of the input DNA. Hybridizing fusion-containing EcoRI fragments were of the expected sizes (data not shown).
Five otherB. dermatitidistransformants whose-galactosidase (LacZ) activity levels were dramatically lower than those of other transformants of the same vector were similarly analyzed (data not shown). Results indicated the partial transfer of a single copy of the T-DNA in three transformants, resulting in the truncation of thelacZfusion. A fourth transformant with low LacZ activity gave a hybridizing PBAD1-lacZfragment of the expected size, but only a single (non-multimerized) copy of the transforming DNA was present in the genome. An additional transformant with low LacZ activity showed that transforming DNA was multimerized and the PYPS3-lacZhybridizing fragment was of the expected size. In this case, the lack of LacZ activity may have resulted from point muta-tions or rearrangements not detected by Southern hybridization. In transforma-tions of vectors containing partialBAD1promoter replacements, three transfor-mants showed low LacZ activity but were not analyzed by Southern hybridization. Since the fate of transforming DNA was not determined for all transformants, LacZ data from these eight aberrant transformants was included in Fig. 2 and 4 and in statistical analyses.
H. capsulatumtransformants were screened by Southern hybridization to se-lect clones in which the transforming DNA was not integrated into the genome. H. capsulatumgenomic DNA was isolated according to the published protocol of Woods and Goldman (55). Undigested genomic DNA samples were probed with an EcoRI fragment of pWU56 containing thePaURA5gene. Episomal vectors were noted as appearing in multiple sizes, including the size of the linearized transforming DNA, a larger form that corresponds to the size of a linear, dimerized vector, and sometimes appearing as both forms in the same transfor-mant. Transformants selected for analysis contained dimerized vectors, the pre-dominant fate of the transforming DNA. To determine the relative copy number of episomes inH. capsulatumtransformants, genomic DNA was serially diluted in round-bottomed wells of a microtiter plate. DNA was denatured by NaOH (final concentration, 0.1 N), with incubation at 37°C for 5 min, followed by neutralization by addition of SSC (1⫻SSC is 0.15 M NaCl plus 0.015 M sodium citrate) to a final concentration of 6⫻. DNA was transferred to nytran using a 96-well vacuum blotting manifold (Schleicher & Schuell, Keene, N.H.). Samples to be compared (e.g., all PBAD1-lacZtransformants) were included on the same membrane. Membranes were first hybridized with thePaURA5probe to deter-mine the relative amounts of transforming DNA per well. Blots were then stripped and reprobed with a 700-bp XhoI-SmaI fragment of theH. capsulatum URA5 (HcURA5) gene (isolated from pWU20) to determine the relative amounts of genomic DNA/well. Hybridization signals were quantified using a Storm blot imaging system and ImageQuant for Macintosh software (Amersham Biosciences, Piscataway, N.J.). ThePaURA5/HcURA5hybridization signal was determined for each well. For each clone, this derived signal was averaged across wells of the dilution series within linearity. The average for each clone was divided by the lowest average value of a clone in its comparison group (P BAD1-lacZor PYPS3-lacZtransformants) to give the relative copy number factor, which varied from 1 to 3.5 (data not shown).
Quantitation of-galactosidase activity.Histoplasmayeast andB. dermatitidis yeast and mycelial cultures were grown in HMM and assayed for-galactosidase activity as previously described (41).-Galactosidase activity was normalized to the protein content of extracts, which were quantified using 10l of supernatant in the BCA microtiter plate assay (Pierce Chemical Co., Rockford, Ill.). Activity fromH. capsulatumtransformants was then also normalized to the relative copy number of vector by dividing the protein-normalized value by the relative copy number factor. Pairwise comparisons of-galactosidase activity values were performed using the two-sided Wilcoxon test.
RESULTS
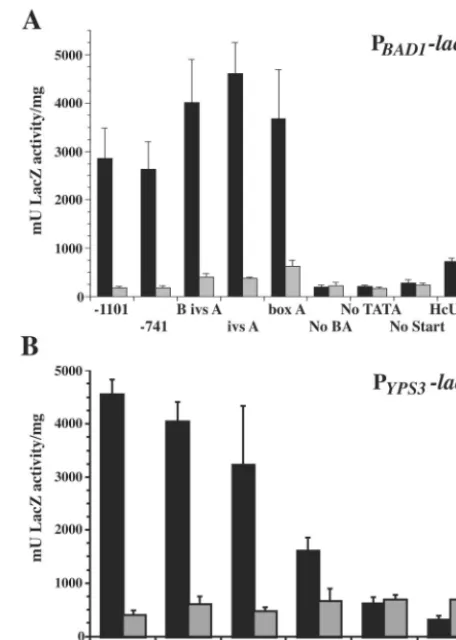
Activity of PBAD1-lacZ fusions in B. dermatitidis. To map
functional regions of theBAD1promoter, a series of vectors containing nested deletions of theBAD1promoter fused to the
E. coli lacZcoding region were introduced intoB. dermatitidis
by Agrobacterium-mediated transformation. Constructs were
designed to test the contributions of boxes B and A, as well as upstream sequences, toBAD1transcription (Fig. 1A). Trans-formants were first analyzed for yeast-phase LacZ activity (Fig. 2A). Those harboring constructs containing BAD1 promoter sequence down to and including the 63-nucleotide box A se-quence possessed robust yeast-phase reporter activity. Within this set, there were no significant differences in LacZ expres-sion. In contrast, transformants with fusions lacking box A yielded LacZ activity consistent with those of control PBAD1
-lacZvectors that lacked the TATA signal (No TATA-lacZ) or
FIG. 2. -Galactosidase activity in B. dermatitidis PBAD1-lacZ, PYPS3-lacZ, and control PURA5-lacZ transformants. B. dermatitidis 26199 transformed withlacZfusion vectors was analyzed in both the yeast and mycelial phase for-galactosidase activity using ano -nitro-phenyl-D-galactopyranoside assay. Activity levels for yeast are shown as black bars, and the levels for mold are shown as shaded bars. Test promoters drivinglacZexpression are indicated on thexaxis according to the designations indicated in Table 1 and/or Fig. 2. Eight indepen-dent transformants of each vector were analyzed in three experiments, and the results were averaged. Values are reported in milliunits of -galactosidase activity per milligram of protein and represent the mean⫾standard error. (A)-Galactosidase activity of transformants of BAD1 promoter truncations fused to lacZ. (B) -Galactosidase activity of transformants ofYPS3promoter truncations fused tolacZ.
on September 8, 2020 by guest
http://ec.asm.org/
transcriptional start site (No Start-lacZ). This represents the background transcriptional activity in transformants of pro-moterless lacZ vectors. The sharp drop in LacZ activity be-tween vectors containing box A and those lacking this region is statistically significant (P⬍0.01).
Reporter activity was also quantified for each transformant grown as a mold. The level of mold-phase LacZ activity de-tected was similar for transformants of all constructs, including the No TATA-lacZ and No Start-lacZ controls (Fig. 2A). These data point to the presence of a positively acting region contributing to yeast-phase activity. Transformants of a control vector in which theH. capsulatum URA5promoter was fused to
lacZshowed similar levels of activity in yeast and mold phases.
Activities of PYPS3-lacZ fusions in B. dermatitidis. The
up-stream region of the YPS3 gene of H. capsulatumwas also analyzed inB. dermatitidisusing a set of constructs containing nestedYPS3promoter deletions fused tolacZ. To determine if the areas of homology between theBAD1andYPS3promoters were functionally conserved, these constructs were designed to match the set ofBAD1promoter deletions (Fig. 1B). Deletions of theYPS3promoter showed a slightly different trend than that seen for theBAD1promoter. While transformants of the
PBAD1-lacZfusions showed consistent LacZ activity from all
constructs containing box A, those of the PYPS3-lacZ fusions displayed a stepwise decline in activity as theYPS3promoter drivinglacZtranscription was successively deleted from the 5⬘
end (Fig. 2B). For example, reporter activity was significantly greater from the construct containing both boxes B and A (B ivs A-lacZ) than from that containing box A alone (box A-lacZ; P ⬍0.001). Though less robust than the reporter activity of longerYPS3upstream regions fused tolacZ, the activity of box A-lacZtransformants was still much greater than that of No BA-lacZtransformants (P⬍0.001). As was the case with the
BAD1 promoter, the loss of box A from theYPS3promoter reduced reporter transcription to background levels. An addi-tional similarity between transformants of theBAD1andYPS3
reporter constructs was the lack of mold-phase LacZ activity. Thus, as withBAD1, a positively acting region is involved in the yeast-phase transcription of YPS3inB. dermatitidis transfor-mants.
An extra construct was included in the analysis of theYPS3
promoter in which the box A region was truncated. This vector lacked the 5⬘half of box A, leaving only the 3⬘half and down-stream sequence fused to lacZ (1/2 box A-lacZ) (Fig. 1B). Strains harboring this fusion gave yeast-phase LacZ activity between that of box A-lacZand that of No BA-lacZ transfor-mants (Fig. 2B). The decrease in activity of transfortransfor-mants of box A-lacZvectors versus that of 1/2 box A-lacZvectors was highly significant (P ⬍ 0.001). Although the level of yeast-phase LacZ activity of 1/2 box A-lacZwas greater than that of the No BA-lacZ fusion (P ⬍ 0.05), it had returned to the background levels seen in mold-phase transformants. This sug-gests that box A is an essential component of the minimal promoter forYPS3.
-Galactosidase activities of BAD1 and YPS3 promoter
truncations inH. capsulatum.A subset of theBAD1andYPS3
-nested promoter deletions fused tolacZwas tested inH.
cap-sulatumto determine if the upstream regions of these genes
function in a manner similar to that seen inB. dermatitidis. In
H. capsulatum, positional effects necessitating the screening of
multiple transformants for each reporter construct can be avoided by the use of vectors that can remain episomal (56). To this end, reporter constructs were cloned into telomeric vec-tors. Yeast-phase H. capsulatum transformants in which the transforming vectors remained episomal were analyzed for LacZ activity. Although quality genomic DNA could not be isolated from mycelium-phase cultures, cells which had under-gone transition to mold were converted back to yeast and reexamined to ensure that the transforming DNA remained episomal. In nearly all samples tested, the transforming DNA had integrated into the genome after conversion, potentially affecting the level of reporter transcription (data not shown). As such, LacZ activity was quantified exclusively from yeast-phase cultures.
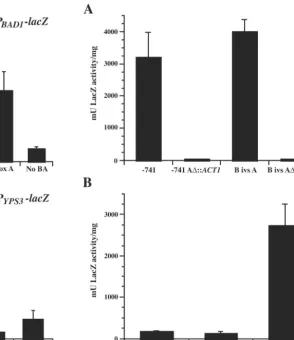
Overall, transcriptional activity from PBAD1-lacZ fusions demonstrated a similar requirement for box A in yeast-phase
H. capsulatum, but some fusions gave unexpected results.
Transformants containing the longestBAD1upstream regions (⫺1101-lacZand⫺741-lacZ) expressed background levels of LacZ activity, equivalent to that in untransformedH. capsula-tum (Fig. 3A; data not shown). Those with BAD1upstream regions from box B (B ivs A-lacZ) to box A (box A-lacZ) possessed LacZ activity, though at a much lower level than that of matchedB. dermatitidistransformants. As with the matched integrated constructs inB. dermatitidis, however, there was no significant difference in LacZ activity between these constructs. Similarly, the No BA-lacZtransformant did not express LacZ activity above background levels, indicating that the minimal promoter region forBAD1inH. capsulatumis the same as that
inB. dermatitidisand requires box A.
H. capsulatumPYPS3-lacZactivity differed in some aspects
from PBAD1-lacZactivity in this fungus. The longest PYPS3-lacZ construct (⫺741-lacZ) produced a high level of LacZ activity (2,200 relative mU/mg). Also, while transformants of vectors containing B ivs A-lacZand ivs A-lacZexpressed LacZ activity, LacZ activity was reduced to background levels in the box A-lacZtransformant (Fig. 3B).
Box A is necessary but not sufficient for transcriptional
activation in B. dermatitidis. Serial truncations of theBAD1
promoter fused tolacZdemonstrated that inclusion of box A was necessary for minimalBAD1promoter activity. To deter-mine whether sequences upstream of box A could compensate for a loss of this region, the 63-bp box A sequence was replaced within constructs containing 741 or 361 bp ofBAD1promoter upstream of the transcriptional start site, fused tolacZ. Box A (⫺175 to⫺237) was replaced with a 63-bp fragment amplified from the coding region of theS. cerevisiaeactin gene. Resulting constructs containing ⫺741 A⌬::ACT1-lacZ and B ivs A⌬::
ACT1-lacZ, and⫺741-lacZand B ivs A-lacZmatched controls,
were introduced intoB. dermatitidis, and transformants were analyzed for yeast-phase LacZ activity. Replacement of box A in either promoter background resulted in a loss of LacZ activity, indicating that this region is necessary for transcription from theBAD1promoter (Fig. 4A).
Since box A is required for activating transcription from the
BAD1promoter in the yeast phase, this sequence was analyzed for its ability to confer transcriptional activation on a foreign promoter inB. dermatitidis. For this purpose, we employed the
H. capsulatum URA5promoter, which produces a low level of
activity inB. dermatitidis(Fig. 2A). Box A, or a 63-bp fragment
on September 8, 2020 by guest
http://ec.asm.org/
of theS. cerevisiaeactin gene, was fused upstream of aURA5
promoter fragment containing 175 bp upstream of the tran-scriptional start site, drivinglacZtranscription. The placement of box A, or the actin fragment, relative to the transcriptional start site was conserved between these constructs and the
BAD1box A-lacZvector. No statistically significant increase in reporter activity was observed from yeast-phaseB. dermatitidis
transformants of the box A-PURA5-lacZ vector over that for those harboring theACT1-PURA5-lacZvector (Fig. 4B). Both groups possessed relatively low levels of promoter activity com-pared to that of the BAD1box A-lacZtransformants. These data indicate that the box A region of theBAD1promoter is not sufficient for transcriptional activation of the H.
capsula-tum URA5promoter.
Analysis ofBAD1upstream region for transcription factor
binding sites.Since the minimalBAD1promoter region inB.
dermatitidis requires box A, we sought to determine if this
region contained any potential binding sites for known tran-scription factors. To this end, the box A sequence was sub-jected to analysis by MatInspector (v3.0) (http://www.genomatix .de) (38), comparing it to both the fungal and vertebrate tran-scription factor binding site libraries. The most probable matches for transcription factors binding this region include a cAMP-responsive element (CRE) and binding sites for the mammalian transcription factors Sox-5, v-Myb, nuclear factor 1 (NF-1), and
FIG. 3. -Galactosidase activity inH. capsulatumPBAD1-lacZand PYPS3-lacZtransformants. Selected yeast-phaseH. capsulatumstrain G217B transformants in which thelacZfusion vectors were episomal were analyzed for-galactosidase activity using ano-nitrophenyl-D
-galactopyranoside assay. Independent transformants analyzed are in-dicated along thexaxis according to the promoter fragment driving
lacZexpression (see Fig. 2). Transformants were analyzed in three experiments, and the results were averaged. Values, scored in milli-units of-galactosidase activity per milligram of protein, were normal-ized to the relative vector copy number per genome (see Materials and Methods) and are reported as relative milliunits of galactosidase ac-tivity per milligram of protein, representing the mean ⫾ standard error. (A)-Galactosidase activities of transformants of BAD1 pro-moter truncations fused to lacZ. (B) -Galactosidase activities of transformants ofYPS3promoter truncations fused tolacZ.
FIG. 4. -Galactosidase activity inB. dermatitidistransformants of reporter fusions containing modifiedBAD1 orURA5promoters. B. dermatitidis26199 transformed withlacZfusion vectors was analyzed in the yeast phase for-galactosidase activity using ano-nitrophenyl -D-galactopyranoside assay. Test promoters drivinglacZexpression are indicated on thexaxis according to the designations indicated in the Results section. Eight independent transformants of each vector were analyzed in three experiments, and the results were averaged. Values are reported as milliunits of-galactosidase activity per milli-gram of protein and represent the mean ⫾standard error. (A)  -Galactosidase activities of transformants of constructs in which the
BAD1promoter is fused tolacZ, comparing activities from box A-con-taining vectors versus those in which box A was replaced with sequence from the actin gene (ACT1) ofS. cerevisiae. (B)-Galactosidase ac-tivities of transformants harboring theURA5promoter fused tolacZ, with the upstream addition of either the box A region of theBAD1
promoter or a similarly sized fragment of theACT1coding region. The activity of theBAD1box A-lacZconstruct is included for comparison.
on September 8, 2020 by guest
http://ec.asm.org/
AREB6 (Fig. 5A). Although fungal homologs to a subset of these factors have been identified, the presence of consensus binding sites may simply suggest that transcription factors with similar DNA binding domains could interact at these locations. In S.
cerevisiae, three ATF/CREB proteins that associate with CRE
have been identified, including both repressors and activators of transcription (51, 19). Sox-5 is a member of a protein family involved in governing cell fate decisions during embryogenesis. These proteins contain a high-mobility group (HMG)-type DNA binding domain that associates with the minor groove and bends DNA, allowing establishment of a local chromatin structure fa-vorable to transcription (36). HMG-box proteins have been iden-tified in fungi, including some factors regulating mating in
Schizo-saccharomyces pombe (20). Overlapping the Sox-5 binding site
was a match with the consensus v-Myb binding site (6). The Myb family of proteins includes transcriptional activators found in many eukaryotes, including plants, mammals, fungi, and Drosoph-ila. Myb family members share a repeated helix-turn-helix DNA binding domain (50). Although the ubiquitous nuclear factor 1 has not been identified in any fungi, its binding site has been noted in another fungal promoter (34, 47). AREB6 is a transcrip-tion factor possessing two zinc-finger DNA binding domains and is important for tissue-specific expression and development in vertebrates (25). It can act as either a positive or a negative regulator of gene expression, depending on the concentration and promoter context. A match to the consensus binding site for the N-terminal zinc finger is present in box A, but AREB6 is pro-posed to only bind this sequence when in conjunction with a second consensus site (CACCTGT) and in such context act as a negative regulator. This second consensus site was not identified in or adjacent to the box A region of theBAD1promoter. The box A region of YPS3contains several identical binding elements, including the CRE and a partial AREB6 site.
Numerous putative transcription factor binding sites were identified elsewhere within the upstream region of theBAD1
promoter (Fig. 6). From the 5⬘end of box B (⫺361) to 741 bp upstream of the transcriptional start site are additional con-sensus matches for v-Myb and CREB binding sites. Also of note, a stress response element (STRE) (16) and a MADS box factor binding site similar to those which bind Rlm1 and Mcm1
of S. cerevisiae and the human myocyte enhancer factor 2
(Mef2a) were identified in this same region (2, 32, 53). Box B contains a GATA site (44), and the intervening sequence be-tween boxes B and A contains a second GATA site and a heat shock element (HSE) homologous to those of S. cerevisiae
(14). Downstream of box A, additional Sox, GATA, and MADS box factor binding sites were identified, as well as binding sites for the A. nidulans conidiation-regulating tran-scription factors AbaA and StuA (3, 15).
Two regions influence transcriptional activation by box A.
The impact of putative transcription factor binding sites within box A on transcription from theBAD1promoter was assessed by modification of theBAD1box A-lacZconstruct, in which a 10-bp sequence encompassing each putative binding site was replaced. B. dermatitidistransformants of five modified con-structs and the control box A-lacZvector were analyzed for LacZ activity in the yeast phase (Fig. 5B). Removal of the putative Sox-5, NF-1, or AREB6 binding sites (replacing⫺217 to⫺208,⫺194 to⫺185, and⫺186 to⫺177, respectively) did not significantly affect reporter activity compared to the box
FIG. 5. Possible transcription factor binding sites within the box A region and their effect on transcription from the BAD1 promoter. (A) Transcription factor binding sites in which the four core bases of the consensus were found at 100% identity and the entire consensus was 90% or greater by MatInspector matrix similarity. Sites are named for the proteins recruited, shown in shaded boxes over the box A sequence. (B and C)-Galactosidase activities of yeast-phaseB. der-matitidistransformants ofBAD1wild-type and modified box A-lacZ
vectors. On thexaxes, modified box A promoters, in which a 10-bp region is replaced, are indicated by a delta symbol, followed by the predicted transcription factor binding site (CRE), or the protein (Sox-5, Myb, NF-1, and AREB6) that may bind the site found within the altered region. Eight independent transformants of each vector were analyzed in three experiments, and the results were averaged. Values are reported as milliunits of-galactosidase activity per milli-gram of protein and represent the mean⫾standard error. Panels B and C represent analyses of transformant sets generated in separate experiments.
on September 8, 2020 by guest
http://ec.asm.org/
A-lacZ control. However, transformants in which ⫺228 to
⫺219 or⫺213 to⫺204, encompassing the predicted CRE and the Myb binding site, were replaced displayed a significant reduction in LacZ activity relative to that with box A-lacZ(P⬍
0.005). Although showing reduced transcriptional activity, nei-ther region alone was essential for the transcriptional activa-tion conferred by box A on theBAD1promoter inB.
derma-titidis. However, replacement of both the CRE and the Myb
binding site within box A resulted in a dramatic reduction in transformant LacZ activity compared to that of the wild-type box A-lacZor either single site replacement reporter (P ⬍
0.001) (Fig. 5C). Thus, the upstream sequences from⫺228 to
⫺219 and⫺213 to⫺204, containing predicted CRE and Myb binding sites, together are critical in transcriptional activation of theBAD1promoter.
DISCUSSION
Herein, we demonstrate the presence of a region conferring transcriptional activation of bothBAD1andYPS3in the yeast phase. Transcription of theH. capsulatumyeast-phase-specific factor CBP1is also activated in the yeast phase, although it does not show sequence similarity to the upstream regions of
YPS3orBAD1. Although both CBP1and YPS3are upregu-lated in the yeast phase,H. capsulatummay possess at least two distinct mechanisms for regulating these phase-specific genes. We found no evidence for repression ofBAD1transcription in the mold phase. TheBAD1promoter contains p30 M bind-ing sites (also present in theYPS3promoter) at positions⫺20,
⫹19, and⫹47 relative to the transcriptional start site (41). In studies ofH. capsulatum, Abidi et al. detected a protein which bound these sites that is present only in nuclear extracts of mycelia, which led them to hypothesize that YPS3 expression is negatively regulated in the mycelial phase (1). None of the
BAD1 promoter truncations fused tolacZyielded activity in mold-phase cells, as one might expect from deletion of a
re-pressor. However, since box A is upstream of the p30 M bind-ing sites and is required for activation of transcription, our approach of serially truncating the BAD1 promoter 5⬘ to 3⬘
may not have been suitable for uncovering deregulated, mold-phase reporter expression. Determining whether the TCC mo-tif regulatesBAD1transcription may require deletions of p30 M sites within the context of a longer promoter including box A. Thus, while box A is required for transcriptional activation, p30 M sites could still contribute to regulation ofBAD1 pro-duction. Activation of transcription in yeast and repression in mold would likely result in tightly regulated expression of this essential virulence factor.
Perhaps the most significant finding from these studies is that the box A region conserved between yeast-phase-specific genes BAD1and YPS3is a required element of the minimal promoter region for BAD1 expression inB. dermatitidisand its heterologous expression inH. capsulatum. This 63-nucleotide box A region is also essential to the minimal promoter region
forYPS3inB. dermatitidis. Interestingly, theYPS3box A-lacZ
fusion was inactive inH. capsulatumtransformants. Since nei-ther gene is shared in the reciprocal fungus, these findings of conserved regulation suggest that the transcriptional machin-ery is shared in these closely related fungi, though subtle dif-ferences in composition or sequence element recognition may exist. This could account for the unexpected silencing and hyperactivation seen only in select H. capsulatum reporter transformants.
DNA sequence analysis identified putative transcription fac-tor binding sites within box A, which were tested functionally, and other sites outside of this region. Replacement of the sequence encompassing the CRE within theBAD1box A-lacZ
vector resulted in a reduction in transcriptional activity inB.
dermatitidis transformants. CRE-binding protein (CREB) is
ubiquitously expressed. In eukaryotes, protein kinases phos-phorylate CREB in response to an increase in cAMP levels, enabling transactivation functions (7). CREB proteins in S.
pombeare targets of signal transduction pathways for handling
osmotic stress and sexual development (8). Phosphorylated CREB protein recruits CREB-binding protein (CBP), which is a gen-eral coactivator molecule able to associate with sevgen-eral tran-scription factors. One mode by which CBP is thought to me-diate CREB-dependent transcriptional activation is by acting as a bridging molecule, linking the DNA-bound CREB to the general transcription factor TFIIB (46). CBP is also an acetyl-transferase. This activity can assist in chromatin remodeling to enhance transcriptional activity or can affect the function of other transcription factors by their direct acetylation (7).
Removal of the predicted Myb binding site from the box A-lacZ vector similarly resulted in a loss in transcriptional activation. The Myb protein family members share a DNA binding motif made up of two to three repeats, each forming a helix-turn-helix structure (33). Vertebrate Myb proteins can also associate with the transactivation domain of CBP, using the same region that binds CREB (46). Transcriptional acti-vation by Myb proteins may be mediated in much the same way as CREB-induced transactivation. Although no homolog of CBP has been identified inS. cerevisiae, tests of v-Myb mole-cules mutated in the activation domain which binds CBP showed an effect on transcriptional activity in a yeast model. This finding suggests the possibility of a functional yeast CBP
FIG. 6. Locations of putative transcription factor binding sites of theBAD1promoter outside of box A. Sites are labeled for the type of protein or proteins recruited (Myb, MADS box, GATA, Sox, AbaA, and StuA) or the sequence element itself (STRE and HSE) and are shown in their relative positions on theBAD1promoter map. Binding sites listed share 100% identity with the four core bases of the tran-scription factor consensus binding site, and the entire consensus was 90% or greater by MatInspector matrix similarity. Note that the MADS box transcription factor binding sites were identified as match-ing that of the mammalian factor Mef2a (32). TheBAD1 transcrip-tional start site (⫹1) is shown as an arrow, and boxes B and A are shown as shaded areas. Actual locations of the binding sites are as follows: STRE,⫺488 to⫺484; Myb,⫺463 to⫺458; CRE,⫺391 to ⫺384; MADS box,⫺411 to⫺402 and⫹69 to⫹78; GATA,⫺352 to ⫺347,⫺310 to⫺305, and⫺54 to⫺49; HSE,⫺277 to⫺264; Sox,⫺146 to⫺140; AbaA,⫹99 to⫹104; StuA,⫹114 to⫹121.
on September 8, 2020 by guest
http://ec.asm.org/
homolog that is involved in Myb-mediated transactivation (52). Recently, a Myb-like protein was identified inGiardia lamblia.
Giardia, like dimorphic fungi, requires cellular differentiation
for its success as a pathogen. In this protozoan, gMyb2 was shown to be important for the expression of the encystation-specific gene gbpi-b, encoding glucosamine-6-phosphate isomerase B, the first enzyme in the biosynthetic pathway for synthesis of cyst wall polysaccharides (49). The binding site for gMyb2 was found in the promoters of four encystation-induced genes. The fungus A. nidulans also utilizes a Myb domain containing transcription factor for inducing genes involved in a cellular shape change. In this case, the transcription factor FlbD is required for early conidiophore production (54). Since Myb family members are involved in regulation of differenti-ation events in diverse systems, from vertebrates (hematopoi-esis) (33) and plants (hair cell formation inArabidopsis) (28) to fungi and protozoans, it is not difficult to envision a role for a Myb-like protein ofB. dermatitidis in the modulation of the yeast-phase-specific transcription ofBAD1.
Loss of both the CRE and Myb binding site within box A resulted in a substantial reduction in reporter activity com-pared to replacement of either single site, indicating that these two regions together are critical for transcriptional activation of theBAD1locus. It is of note that theYPS31/2 box A-lacZ
construct, which gave equivalent levels of LacZ activity in yeast- and mold-phase transformants (Fig. 2B), is lacking the regions corresponding to the CREB, Sox-5, and v-Myb binding sites within box A ofBAD1. Although both CREs and Myb binding sites were found upstream of box A, these sites were unable to compensate for the loss of box A in the ⫺741
A⌬::ACT1-lacZand B ivs A⌬::ACT1-lacZvectors.
The binding sites of other transcription factors implicated in governing gene expression during changes in cellular morphol-ogy were identified outside of box A (Fig. 6). MADS-box transcription factor binding sites found in theBAD1promoter shared homology to the consensus binding site for theS.
cer-evisiaeprotein Mcm1p, whoseC. albicanshomolog was shown
to be important for morphogenesis (42). Binding sites for the
A. nidulansconidiation regulators AbaA and StuA were also
identified in the BAD1 upstream. The processes of conidia generation and yeast formation from fungi of a filamentous morphotype share similarities. Both require the coupling of nuclear division with cytokinesis to go from a multicellular mycelium to single cells. Conidiation requires the initial step of generating unicellular forms (metulae and phialides) that give rise to conidia. The formation of these structures is governed by several transcription factors, including AbaA and StuA (30, 29). In the dimorphic fungusPenicillium marneffeii, we see the linkage between conidiation and yeast formation, since AbaA was shown to be required for both processes (9). MADS-box transcription factor, AbaA, and StuA binding sites were found downstream of box A. Since this region was included in the
BAD1box A-lacZconstruct, these sites could contribute to the level of transcriptional activity seen in its transformants. The inability of box A alone to stimulate transcriptional activation of the H. capsulatum URA5promoter suggests that elements downstream of box A may contribute to transcriptional activa-tion from the BAD1 promoter.
The identification of HSE in theBAD1upstream was sur-prising, since heat shock responses are generally thought of as
reactions to a stress on the cells, rather than a condition for which an organism has a specially adapted morphotype. It is known, however, that the heat shock factor inducing transcrip-tion of heat shock proteins inS. cerevisiae is encoded by an essential gene, indicating that heat shock proteins are neces-sary for survival under normal growth conditions. STRE is found in nearly 200 potential STRE-regulated genes in yeast. Differing patterns of expression of stress-induced genes con-taining STRE are thought to be due to promoter context. The combination of the HSE with the STRE is found in at least two yeast genes,SSA3, which encodes an Hsp70 involved in pre-vention of protein aggregation, andHSP104, which has been shown to rescue heat-inactivated proteins (16). The HSE and STRE are both found in theBAD1upstream region. Although these elements were found upstream of the minimal promoter, they may still play a role in regulation of BAD1 expression. Utilization of these elements may be responsible for the rapid onset ofBAD1gene expression after morphogenesis is stimu-lated by switching the cells from 25 to 37°C, while other factors may be involved in maintenance of BAD1 transcription in stable yeast-phase cells.
ACKNOWLEDGMENTS
This work was supported by NIH grants AI035681 and AI50882 from the USPHS and by a Burroughs Wellcome Fund Scholar Award in Molecular Pathogenic Mycology (B.S.K.).
We thank Jens Eickhoff of the Department of Biostatistics and Medical Informatics, University of Wisconsin, for assistance with sta-tistical analyses.
REFERENCES
1. Abidi, F. E., H. Roh, and E. J. Keath.1998. Identification and characteriza-tion of a phase-specific, nuclear DNA binding protein from the dimorphic pathogenic fungusHistoplasma capsulatum. Infect. Immun.66:3867–3873. 2. Acton, T. B., H. Zhong, and A. K. Vershon.1997. DNA-binding specificity of
Mcm1: operator mutations that alter DNA-bending and transcriptional ac-tivities by a MADS box protein. Mol. Cell. Biol.17:1881–1889.
3. Andrianopoulos, A., and W. E. Timerlake.1994. TheAspergillus nidulans abaAgene encodes a transcriptional activator that acts as a genetic switch to control development. Mol. Cell. Biol.14:2503–2515.
4. Beijersbergen, A., A. den Dulk-Ras, R. A. Schilperoort, and P. J. J. Hooy-kaas.1992. Conjugative transfer by the virulence system ofAgrobacterium tumefaciens. Science256:1324–1327.
5. Bevan, M.1984. Binary Agrobacteriumvectors for plant transformation. Nucleic Acids Res.12:8711–8721.
6. Biedenkapp, H., U. Borgmeyer, A. E. Sippel, and K. H. Klempnauer.1988. Viral myb oncogene encodes a sequence-specific DNA-binding activity. Na-ture335:835–837.
7. Blobel, G. A.2000. CREB-binding protein and p300: molecular integrators of hematopoietic transcription. Blood95:745–755.
8. Borgnes-Walmsley, M. I., and A. R. Walmsley.2000. cAMP signaling in pathogenic fungi: control of dimorphic switching and pathogenicity. Trends Microbiol.8:133–141.
9. Borneman, A. R., M. J. Hynes, and A. Andrianopoulos.2000. TheabaA homologue ofPenecillium marneffeiparticipates in two developmental pro-grammes: conidiation and dimorphic growth. Mol. Microbiol.38:1034–1047. 10. Bradsher, R. W.1996. Histoplasmosis and blastomycosis. Clin. Infect. Dis.
22:S102–S111.
11. Brandhorst, T., M. Wu¨thrich, B. Finkel-Jimenez, and B. Klein.2003. A C-terminal EGF-like domain governs BAD1 localization to the yeast surface and fungal adherence to phagocytes, but is dispensable in immune modula-tion and pathogenicity ofBlastomyces dermatitidis. Mol. Microbiol.48:53–65. 12. Brandhorst, T. T., M. Wu¨thrich, T. Warner, and B. S. Klein.1999. Targeted gene disruption reveals an adhesin indispensable for pathogenicity of Blas-tomyces dermatitidis. J. Exp. Med.189:1207–1216.
13. Burg, E. F., III, and L. H. Smith, Jr.1994. Cloning and characterization of bys1, a temperature-dependent cDNA specific to the yeast phase of the pathogenic dimorphic fungusBlastomyces dermatitidis. Infect. Immun.62:
2521–2528.
14. Chen, J., and D. S. Pederson.1993. A distal heat shock element promotes the rapid response to heat shock of theHSP26gene in the yeastSaccharomyces cerevisiae. J. Biol. Chem.268:7442–7448.
on September 8, 2020 by guest
http://ec.asm.org/
15. Dutton, J. R., S. Johns, and B. L. Miller.1997. StuAp is a sequence-specific transcription factor that regulates developmental complexity inAspergillus nidulans. EMBO J.16:5710–5721.
16. Estruch, F. 2000. Stress-controlled transcription factors, stress-induced genes and stress tolerance in budding yeast. FEMS Microbiol. Rev.24:469– 486.
17. Finkel-Jimenez, B., M. Wu¨thrich, T. Brandhorst, and B. S. Klein.2001. The WI-1 adhesin blocks phagocyte TNF-␣production, imparting pathogenicity onBlastomyces dermatitidis. J. Immunol.166:2665–2673.
18. Finkel-Jimenez, B., M. Wu¨thrich, and B. S. Klein.2002. BAD1, an essential virulence factor ofBlastomyces dermatitidis, suppresses host TNF-␣ produc-tion through TGF--dependent and -independent mechanisms. J. Immunol.
168:5746–5755.
19. Garcia-Gimeno, M. A., and K. Struhl.2000. Aca1 and Aca2, ATF/CREB activators inSaccharomyces cerevisiae, are important for carbon source uti-lization but not the response to stress. Mol. Cell. Biol.20:4340–4349. 20. Grosschedl, R., K. Giese, and J. Pagel.1994. HMG domain proteins:
archi-tectural elements in the assembly of nucleoprotein structures. Trends Genet.
10:94–100.
21. Hogan, L. H., and B. S. Klein. 1997. Transforming DNA integrates at multiple sites in the dimorphic fungal pathogenBlastomyces dermatitidis. Gene186:219–226.
22. Hung, C.-Y., N. M. Ampel, L. Christian, K. R. Seshan, and G. T. Cole.2000. A major cell surface antigen ofCoccidioides immitiswhich elicits both hu-moral and cellular immune responses. Infect. Immun.68:584–593. 23. Hung, C.-Y., J.-J. Yu, K. R. Seshan, U. Reichard, and G. T. Cole.2002. A
parasitic phase-specific adhesin ofCoccidioides immitiscontributes to the virulence of this respiratory fungal pathogen. Infect. Immun.70:3443–3456. 24. Ignatov, A., and E. J. Keath.2002. Gel shift assay of nuclear extracts from Histoplasma capsulatumdemonstrates the presence of several DNA binding proteins. Infect. Immun.70:2238–2241.
25. Ikeda, K., and K. Kawakami.1995. DNA binding through distinct domains of zinc-finger-homeodomain protein AREB6 has different effects on gene transcription. Eur. J. Biochem.233:73–82.
26. Keath, E. J., and F. E. Abidi.1994. Molecular cloning and sequence analysis ofyps-3, a yeast-phase-specific gene in the dimorphic fungal pathogen His-toplasma capsulatum. Microbiology140:759–767.
27. Klein, B. S., and S. L. Newman.1996. Role of cell surface molecules on Blastomyces dermatitidisin host:pathogen interactions. Trends Microbiol.
4:246–251.
28. Martin, C., and J. Paz-Ares.1997. MYB transcription factors in plants. Trends Genet.13:67–73.
29. Miller, K. Y., J. Wu, and B. L. Miller.1992. StuA is required for cell pattern formation inAspergillus. Genes Dev.6:1770–1782.
30. Mirabito, P. M., T. H. Adams, and W. E. Timberlake.1989. Interactions of three sequentially expressed genes control temporal and spatial specificity in Aspergillusdevelopment. Cell57:859–868.
31. Newman, S. L., S. Chaturvedi, and B. S. Klein.1995. The WI-1 antigen on Blastomyces dermatitidis yeasts mediates binding to human macrophage CD18 and CD14 receptors. J. Immunol.154:753–761.
32. Nurrish, S., and R. Treisman.1995. DNA binding specificity determinants in MADS-box transcription factors. Mol. Cell. Biol.15:4076–4085.
33. Oh, I.-H., and E. P. Reddy.1999. Themyb gene family in cell growth, differentiation and apoptosis. Oncogene18:3017–3033.
34. Paonessa, G., F. Gounari, R. Frank, and R. Cortese.1988. Purification of a NF1-like DNA-binding protein from rat liver and cloning of the correspond-ing cDNA. EMBO J.7:3115–3123.
35. Patel, J. B., J. W. Batanghari, and W. E. Goldman.1998. Probing the yeast phase-specific expression of theCBP1gene inHistoplasma capsulatum. J. Bacteriol.180:1786–1792.
36. Penvy, L. H., and R. Lovell-Badge.1997.Soxgenes find their feet. Curr. Opin. Genet. Dev.7:338–344.
37. Punt, P. J., R. P. Oliver, M. A. Dingemanse, P. H. Pouwels, and C. A. M. J. J. van den Hondel.1987. Transformation ofAspergillusbased on the hygro-mycin B resistance marker fromE. coli.Gene56:117–124.
38. Quandt, K., K. Frech, H. Karas, E. Wingender, and T. Werner.1995. MatInd and MatInspector: new fast and versatile tools for detection of consensus matches in nucleotide sequence data. Nucleic Acids Res.23:4878–4884. 39. Retallack, D. M., E. L. Heinecke, R. Gibbons, G. S. Deepe, Jr., and J. P.
Woods.1999. The URA5 gene is necessary for Histoplasma capsulatum growth during infection of mouse and human cells. Infect. Immun.67:624– 629.
40. Rooney, P. J., and B. S. Klein.2002. Linking fungal morphogenesis with virulence. Cell. Microbiol.3:127–138.
41. Rooney, P. J., T. D. Sullivan, and B. S. Klein.2000. Selective expression of the virulence factor BAD1 upon morphogenesis to the pathogenic yeast form ofBlastomyces dermatitidis: evidence for transcriptional regulation by a con-served mechanism. Mol. Microbiol.39:875–889.
42. Rottmann, M., S. Dieter, H. Brunner, and S. Rupp.2003. A screen in Saccharomyces cerevisiaeidentifiedCaMCM1, an essential gene inCandida albicanscrucial for morphogenesis. Mol. Microbiol.47:943–959.
43. Sambrook, J., E. F. Fritsch, and T. Maniatis.1989. Molecular cloning: a laboratory manual, 2nd ed.Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
44. Scazzocchio, C.2000. The fungal GATA factors. Curr. Opin. Microbiol.
3:126–131.
45. Sebghati, T. S., J. T. Engle, and W. E. Goldman.2000. Intracellular para-sitism byHistoplasma capsulatum: fungal virulence and calcium dependence. Science290:1368–1372.
46. Shimikama, N., J. Lyon, and N. E. La Thangue.1997. The p300/CBP family: integrating signals with transcription factors and chromatin. Trends Cell Biol.7:230–236.
47. Soll, D. R.1997. Gene regulation during high-frequency switching in Can-dida albicans. Microbiology143:279–288.
48. Sullivan, T. D., P. J. Rooney, and B. S. Klein.2002. Agrobacterium tume-faciens integrates transfer DNA into single chromosomal sites of dimorphic fungi and yields homokaryotic progeny from multinucleate yeast. Eukaryot. Cell1:895–905.
49. Sun, C.-H., D. Palm, A. G. McArthur, S. G. Sva¨rd, and F. D. Gillin.2002. A novel Myb-related protein involved in transcriptional activation of encysta-tions genes inGiardia lamblia. Mol. Microbiol.46:971–984.
50. Tanikawa, J., T. Yasukawa, M. Enari, K. Ogata, Y. Nishimura, S. Ishii, and A. Sarai.1993. Recognition of specific DNA sequences by thec-myb pro-tooncogene product: role of three repeat units in the DNA-binding domain. Proc. Natl. Acad. Sci. USA90:9320–9324.
51. Vincent, A. C., and K. Struhl.1992. ACR1, a yeast ATF/CREB repressor. Mol. Cell. Biol.12:5394–5405.
52. Wang, D.-M., and J. S. Lipsick.2002. Mutational analysis of the transcrip-tional activation domains of v-Myb. Oncogene21:1611–1615.
53. Watanabe, Y., K. Irie, and K. Matsumoto.1995. YeastRLM1encodes a serum response factor-like protein that may function downstream of the Mpk1 (Slt2) mitogen-activated protein kinase pathway. Mol. Cell. Biol.15:
5740–5749.
54. Wieser, J., and T. H. Adams.1995.flbDencodes a Myb-like DNA-binding protein that coordinates initiation ofAspergillus nidulansconidiophore de-velopment. Genes Dev.9:491–502.
55. Woods, J. P., and W. E. Goldman.1992. In vivo generation of linear plasmids with addition of telomeric sequences byHistoplasma capsulatum. Mol. Mi-crobiol.6:3603–3610.
56. Woods, J. P., and W. E. Goldman.1993. Autonomous replication of foreign DNA inHistoplasma capsulatum: role of native telomeric sequences. J. Bacteriol.175:636–641.
57. Woods, J. P., E. L. Heinecke, and W. E. Goldman.1998. Electrotransforma-tion and expression of bacterial genes encoding hygromycin phosphotrans-ferase and-galactosidase in the pathogenic fungusHistoplasma capsulatum. Infect. Immun.66:1697–1707.
58. Worsham, P. L., and W. E. Goldman.1988. Quantitative plating of His-toplasma capsulatum without addition of conditioned medium or sid-erophores. J. Med. Vet. Mycol.26:137–143.