Improved Mg
2
1
-Based Reverse Transcriptase Assay for
Detection of Primate Retroviruses
JOHNNA F. SEARS,1ROY REPASKE,2†ANDARIFA S. KHAN1*
Laboratory of Retrovirus Research, Division of Viral Products, Center for Biologics Evaluation and Research,
U.S. Food and Drug Administration,1and Laboratory of Molecular Microbiology, National Institute of
Allergy and Infectious Diseases,2Bethesda, Maryland 20892
Received 30 December 1998/Accepted 22 February 1999
The reverse transcriptase (RT) assay is a simple, relatively inexpensive, widely used assay that can detect all retroviruses (known and novel retroviruses as well as infectious and defective retroviruses) on the basis of the
divalent cation requirement of their RT enzyme, i.e., Mg21or Mn21. Descriptions of various RT assays have
been published; however, they cannot be directly applied to the analysis of biological products or clinical samples without further standardization to determine the lower limit of virus detection (sensitivity), assay variability (reproducibility), or ability to detect different retroviruses (specificity). We describe the detection of
type E and type D primate retroviruses, which may be pathogenic for humans, by a new32P-based, Mg21
-containing RT assay. The results show that the sensitivity of detection is <3.2 50% tissue culture infective doses
(TCID50s) for human immunodeficiency virus type 1 (HIV-1) and <1 TCID50for simian immunodeficiency
virus isolated from a rhesus macaque (SIVmac). Analysis of recombinant HIV-1 RT enzyme indicated that 1025
U, which is equivalent to 4.25 3 104 virions, could be detected. Additionally, genetically distinct type D
retroviruses such as simian AIDS retrovirus and squirrel monkey retrovirus were also detected in the assay
with similar sensitivities. Thus, the improved RT assay can be used to detect genetically divergent Mg21
-dependent retroviruses of human and simian origin that can infect human cells and that therefore pose a potential health risk to humans.
All retroviruses can be divided into two categories on the basis of the presence of an Mg21- or Mn21-requiring RNA-dependent DNA polymerase that is termed reverse transcrip-tase (RT) (1, 22) and that is critical in the retroviral life cycle (23, 24). Each group includes retroviruses of diverse origins that are structurally distinct and genetically divergent but that share similar cation requirements for their RT activity. For example, different retrovirus types (21) such as avian type C retroviruses, primate type D retroviruses, and primate type E lentiviruses, which includes human immunodeficiency virus (HIV) type 1 (HIV-1), can be grouped together on the basis of the presence of an Mg21-requiring RT in these viruses. Thus, the detection of RT activity can generally indicate the presence of a retrovirus in the absence of specific information regarding its genome or protein content. Although RT assays generally detect about 105to 106virus particles and are not as sensitive as infec-tivity or PCR assays, they are broadly reactive and have been used for the detection and isolation of different types of novel retrovi-ruses including HIV-1 (2, 6). In addition, RT assays are routinely used in infectivity studies for the rapid and easy monitoring of retrovirus infection and replication. The detection of small amounts of retrovirus by the RT assay may be made possible by virus amplification in a susceptible cell line or by increasing the virus concentration in a sample so that it is above the detection limit of the assay, e.g., by centrifugation. The RT assay is also widely used for analysis of potential retroviral contaminants in biological products, which may be introduced during passage
through animals, during propagation in cell substrates, or from biological raw materials used in production (12).
Several Mg21-based RT assays have been developed; how-ever, the viruses used in most of the studies have been avian myeloblastosis virus or HIV-1. Furthermore, the previous stud-ies describe details regarding assay development; however, there is little information about assay standardization, includ-ing sensitivity of virus detection, ability to detect different ret-roviruses, or assay variability. Such information is especially important when an RT assay is used to demonstrate the ab-sence of retroviral contaminants in biological products and in analyses of clinical samples from potentially infected individ-uals. Current HIV-1 RT assays have been used in infectivity studies, neutralization assays, and assessments of antiviral ef-fects. The original assays used [3H]deoxynucleoside triphos-phates to extend the oligonucleotide primer to produce the cDNA copy of the homopolymer template (2, 6, 10). Modifi-cations of the RT assays have been made to increase the sensitivity of virus detection which include the use of32P- and 125I-radiolabeled nucleotide substrates (7, 25). Additionally, RT assays with increased sensitivities have been developed with nonisotopically labeled nucleotides; however, this was achieved after a prolonged incubation, i.e., 15 to 24 h (3, 5, 20). In this paper we describe the standardization of a new 32 P-based RT cocktail, with a 2-h incubation period, for the gen-eral detection of retroviruses that contain Mg21-requiring RT, including type E lentiviruses (e.g., HIV-1 and simian immuno-deficiency virus [SIV]) as well as two distinct type D retrovi-ruses, i.e., simian AIDS retrovirus (SRV) (4, 16) and squirrel monkey retrovirus (SMRV) (8).
MATERIALS AND METHODS
RT assays.The new RT cocktail contains the following: 6 mM MgCl2, 3mg of poly(A) (P-L Biochemicals Inc. Milwaukee, Wis.) per ml, 0.021mg of p(dT)12–18 * Corresponding author. Mailing address: Laboratory of Retrovirus
Research, Division of Viral Products, Center for Biologics Evaluation and Research, U.S. Food and Drug Administration, 1401 Rockville Pike, HFM-454, Rockville, MD 20852-1448. Phone: (301) 827-0791. Fax: (301) 496-1810. E-mail: [email protected].
† Retired.
1704
on May 15, 2020 by guest
http://jcm.asm.org/
(Pharmacia Biotech, Piscataway, N.J.) per ml, 0.12% Nonidet P-40, 24 mM triethanolamine (TEA; U.S. Biochemical Corp., Cleveland, Ohio), and 28.8 mM EGTA (U.S. Biochemical Corp.). TEA-EGTA was formulated separately by mixing 3.93 ml of TEA and 13.146 g of EGTA in a final volume of 300 ml (pH 8.0). This cocktail was aliquoted and stored at220°C such that it would undergo only two freeze-thaws prior to its use. Additionally, the cocktail was thawed on ice or quickly at 37°C and was immediately put on ice. Under these handling conditions the cocktail was found to remain stable for at least a year, as deter-mined by the RT activity of a standard control virus. Just prior to use of the cocktail, the following were added per milliliter of chilled cocktail: 4ml of 1 M dithiothreitol, 3ml of [a-32P]dTTP (1.5mCi;.400 Ci/mmol; Amersham Corp., Arlington Heights, Ill.), and 1 ml of 1024M dTTP. The dithiothreitol and nonradioactive dTTP solutions were also aliquoted individually and stored at 220°C to avoid additional freezing and thawing. Ten microliters of test sample was incubated with 50 ml of the RT cocktail in a screw-cap, 1.5-ml conical polypropylene tube (Starstedt, Arlington, Tex.) for 2 h in a 37°C water bath. Five microliters of the reaction mixture was spotted, in duplicate, onto DE81 paper (Whatman, Maidstone, United Kingdom), air dried, and washed at room tem-perature on a rocker in 23SSC (13SSC is 0.15 M NaCl plus 0.015 M sodium citrate) four times each for 5 min and two times each for 1 min with 95% ethanol. The paper was air dried and exposed overnight at280°C to X-OMAT AR film (Kodak, Rochester, N.Y.). The next day the spots were cut out and counted in a scintillation counter.
In all the experiments described in this paper, the radioisotope was used prior to or on the manufacturer’s reference date. Dilutions of viruses and enzyme were prepared as described below and were immediately used for RT analyses. The negative control was complete medium (described below).
Viruses.HIV-1 was prepared in human peripheral blood mononuclear cells (PBMCs) by using strain LAI, which had been grown in CEM clone 12D7 cells (obtained as strain LAV.04from M. A. Martin, National Institute of Allergy and Infectious Diseases). For virus titration, human PBMCs (106 cells/ml) were stimulated with phytohemagglutinin (PHA; Murex Diagnostics, Dartford, United Kingdom), which was used at a final concentration of 250 ng/ml in complete RPMI 1640 medium containing 10% fetal bovine serum (Hyclone, Logan, Utah), 10 mM HEPES (Biofluids, Rockville Md.), 250 U of penicillin per ml, 250mg of streptomycin per ml, and 2 mML-glutamine (Life Technologies, Gibco BRL, Grand Island, N.Y.). After 3 days, the PHA-containing medium was replaced with medium containing 10% interleukin-2 (Hemagen Diagnostics, Columbia, Md.), and the cells were resuspended at a final concentration of 106 cells/0.1 ml. Virus infection was set up in a 24-well plate: 0.1 ml of 1021to 1026 serial dilutions of virus were initially incubated for 1 h at 37°C with 0.1 ml of 106 PBMCs. After 1 h, 1.8 ml of medium was added and the cells were cultured in a 5% CO2atmosphere at 37°C. After 3 to 4 days, half of the medium was removed and was replaced with fresh medium. On day 7 postinfection, the supernatant was filtered through a 0.45-mm-pore-size filter unit (Spin-X Centri-fuge Tube Filters; Costar, Cambridge, Mass.) and collected for the RT assay. The virus infections were set up in quadruplicate. The virus titer was calculated as described by Reed and Muench (18) and was expressed as the 50% tissue culture infectious dose (TCID50) per milliliter. The titer of the HIV-1 stock was 104.5 TCID50s/ml in human PBMCs.
SIVmac was obtained by ligation of cloned 59 and 39 DNA fragments of SIVmac-mm239(11) and transfection in 1743CEM cells. The virus obtained was propagated in rhesus monkey PBMCs to produce a large-scale virus stock (6a). The titer of the monkey-grown SIVmacstock was determined as described above for HIV-1, except that the monkey PBMCs were stimulated with 500 ng of PHA per ml. The titer of SIVmac239at passage 1 in autologous monkey PBMCs was determined as 104.0TCID
50s/ml.
For RT assay analysis of HIV-1 and SIV, each virus was serially diluted in complete RPMI 1640 medium. At least two independently prepared dilutions were assayed in at least two separate RT assays.
SRV type 1 (SRV-1) and SMRV were purchased from Advanced Biotechnol-ogies Inc. (Columbia, Md.) and were at 8.26 3107and 1.92 31011virus particles/ml, respectively, on the basis of electron microscopy. Serial dilutions of the viruses were made in complete RPMI 1640 medium. For each virus, one dilution series was used and the RT assay was done in duplicate.
RT enzyme.Recombinant HIV-1 RT was purchased from Worthington Bio-chemical Corporation (Freehold, N.J.; supplied as 48.7 U/ml). Serial dilutions of the enzyme were made in complete medium and were used immediately in RT assays.
RESULTS
The linear range of the RT reaction was determined by assaying the RT activity at various times ranging from 30 to 180 min (Fig. 1). Independent RT reactions with 10 ml of undiluted HIV-1 and SIVmacwere set up for each time point as described in Materials and Methods. Each reaction was terminated at the times indicated in Fig. 1 by spotting in duplicate 5 ml of the reaction mixture onto DE81 filter paper. All the reaction mixtures were spotted on a single
filter paper until the reaction mixture from last time point was spotted. The paper was then washed as described in Materials and Methods. The results indicated that the RT activities for HIV and SIV were directly proportional to the reaction times up to 180 min. The rate of dTTP incorpora-tion was found to be directly proporincorpora-tional to the virus con-centration as well as to the amount of [32P]dTTP used in the assay (data not shown). The RT activity increased with the amount of [32P]dTTP with 1 to 10ml (0.5 to 5mCi) of the radiolabeled deoxynucleoside triphosphate.
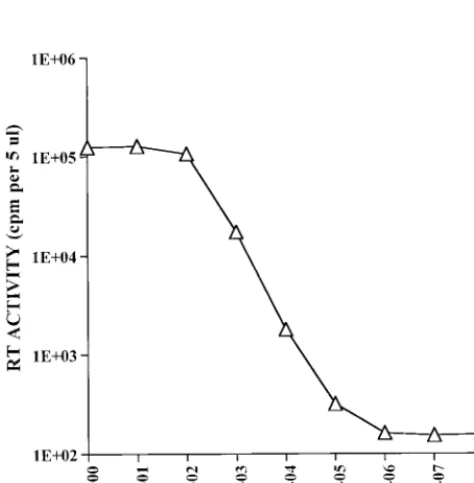
The sensitivity of detection of type E lentiviruses in the RT assay was determined with HIV-1 and SIV. The results are presented in Table 1. Ten microliters of undiluted virus (first sample) and serial dilutions were assayed for RT activity. The sensitivity of detection of HIV-1 was,3.2 TCID50s. A similar sensitivity of detection was seen with a different, independently prepared HIV-1 stock (13). The sensitivity of detection of SIVmac was ,1 TCID50. Furthermore, in repeated, parallel experiments, the sensitivity of HIV-1 and SIV detection was 10-fold greater by the new RT assay than by a previously published assay (25). In a comparison of the new RT assay with another HIV-1 RT assay published by Hoffman et al. (10), sevenfold greater RT activity was detected when two HIV-1 concentrations were tested with the new cocktail (18a). To further assess assay sensitivity, RT analysis was done with serial dilutions of recombinant HIV-1 RT. Similar results were ob-tained with two independently prepared dilutions. The results of RT analysis of one dilution series, which was assayed in duplicate, are presented in Fig. 2. The sensitivity of detection was 1025U of HIV-1 RT enzyme.
The ability of the new RT assay to detect other retrovi-ruses containing an Mg21-requiring RT enzyme, such as type D retroviruses, was assessed with SRV and SMRV. In this case the sensitivity of detection was based upon the use of virus stocks with known numbers of particles, as deter-mined by electron microscopy, and therefore may be less
FIG. 1. Linear range of the RT assay. The linearity of the RT assay was determined by HIV-1LAIand SIVmac-mm239. Ten microliters of undiluted virus was assayed at the indicated time intervals. The RT activity is indicated. The mean6standard deviation from one experiment is indicated.Œ, HIV-1;■, SIV; E, medium.
on May 15, 2020 by guest
http://jcm.asm.org/
accurate than when the HIV-1 and SIV stocks were used, in which case the number of infectious particles was deter-mined on the basis of an infectivity assay. The results pre-sented in Table 2 indicate that about 2 3 105 virions of SMRV were detected and that about 83104virions of SRV were detected.
It should be noted that because32P was used in the assay, the results could be monitored by both scintillation counting and autoradiography. A parallel analysis was done in all cases; it was found that a weakly positive signal was easier to visualize from the autoradiogram. On the basis of autoradiography, a result was positive if the counts were 50% above the back-ground RT activity.
DISCUSSION
The RT assay is widely used for the general detection of known and novel retroviruses. This is primarily because it is easy, quick, and relatively inexpensive to perform. Recently, highly sensitive PCR-based RT assays which can detect 3 to 100 virions have been developed (9, 17, 19); however, their use at this time is limited because the assay is technically demand-ing and expensive. Thus, parallel efforts have continued to increase the sensitivity of retrovirus detection by the conven-tional RT assays.
In this paper, we describe the standardization of an im-proved Mg21-based RT assay which can detect different types of primate retroviruses. The sensitivity of the assay was deter-mined to be ,1 TCID50 of SIVmac and ,3.2 TCID50s of HIV-1. Analysis of the recombinant HIV-1 RT enzyme indi-TABLE 1. Detection of HIV-1 and SIV in RT assay
HIV-1 SIV
No. of infectious
particlesa RT activityb Autoradresultc No. of infectiousparticlesa RT activityb Autoradresultc
316.2 4,25361,705 1 100 3,54561,184 1
63.2 1,3476410 1 20 9696123 1
31.6 7356188 1 10 5056116 1
6.3 258618 1 2 183640 1
3.2 183621 1 1 147645 1
0.63 115627 2 0.2 108631 2
0.32 109639 2 0.1 108648 2
0.063 98632 2 0.02 108637 2
0.032 107633 2 0.01 100636 2
Medium 102644 2 Medium 102644 2
aNumber of infectious virions in 10ml of undiluted (first) sample and serially diluted samples on the basis of an HIV-1 stock titer of 104.5TCID
50s per ml in human PBMCs and an SIV stock titer of 104.0TCID
50s per ml in rhesus monkey PBMCs.
bRT activity is counts per minute per 5ml of spotted reaction mixture. In the case of HIV-1 the mean6standard deviation was calculated for two independent RT
assays in which each sample was spotted twice, and in the case of SIV the mean6standard deviation was calculated for four spots (two per sample) from two independent RT assays. Each assay was performed with an independently prepared virus dilution series.
cAutorad, autoradiography. An autoradiogram was exposed overnight. The results are indicated as positive and negative and do not reflect differences in the
[image:3.612.54.548.84.218.2]intensities of the positive signals.
[image:3.612.55.292.451.692.2]FIG. 2. Detection of recombinant HIV-1 RT enzyme. Serial dilutions of HIV-1 RT enzyme were analyzed by the RT assay. The RT activity is indicated. The mean6standard deviation was calculated for four spots obtained from two RT assays which were done in duplicate with one dilution series.
TABLE 2. Detection of type D retroviruses in RT assays Virus and no. of
virus particlesa RT activityb Autoradresultc
SMRV
1.923107 5,1636228 1
1.923106 574624 1
1.923105 114610 1
1.923104 7965 2
1.923103 7765 2
SRV
8.253105 5476146 1
8.253104 8569 1
8.253103 65615 2
aThe number of virus particles in 10ml is indicated on the basis of the particle
count obtained by electron microscopy by Advanced Biotechnologies Inc. The first sample in the case of SMRV represents a 1:100 dilution of the original sample, and the rest of the samples are 10-fold serial dilutions. In the case of SRV the first sample is undiluted virus and the rest of the samples are 10-fold serial dilutions.
bThe RT activity is counts per minute per 5ml and is reported as the mean6
standard deviation for two RT assays which were done in duplicate, with each sample being spotted twice.
cAutorad, autoradiography. Results are based upon visual examination of an
overnight exposure of the autoradiogram. The results are indicated as positive and negative and do not reflect the differences in the intensities of the positive signals.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:3.612.312.548.495.622.2]cated the detection of 1025 U, which, on the basis of the molecular weight (117,000), was equivalent to 3.43106 mol-ecules of RT. Since it is reported that 80 molmol-ecules of RT are present per HIV-1 particle (15), the sensitivity of detection was calculated to be,4.253104virions. These results thus indi-cate a possible 1:10,000 ratio of infectious virus to total parti-cles, which is consistent with previously reported results (14). In addition to primate lentiviruses, type D retroviruses of ge-netically diverse origins were detected at about 83104virions of SRV, which was isolated from an Old World monkey, and about 23 105 virions of SMRV, which was isolated from a New World monkey.
The sensitivity of detection was achieved with a relatively short incubation time (2 h), in contrast to some other RT assays that require prolonged incubation times (i.e., 15 to 24 h) for increased detection (5, 20). The sensitivity of the new RT assay for HIV and SIV detection can be further improved by increasing the incubation time to up to 3 h and/or by increasing the amount of [32P]dTTP. Conversely, with samples containing adequate amounts of virus, a minimum amount of radiolabeled dTTP and/or a reduced reaction time can be used, since the counts obtained are directly proportional to the incubation time and to the [32P]dTTP concentration.
The results obtained with the new RT assay were highly reproducible; similar results were obtained with regard to the sensitivity of detection of HIV and SIV when independently prepared virus stocks or dilutions were used. However, some variability in the counts per minute incorporated can occur between different assays, especially due to handling, e.g., when the sample or the radioisotope is pipetted or when the final reaction mixture is spotted onto the filter paper. We have reduced interassay variability by using designated and accu-rately calibrated pipetting devices for the different handling procedures. One caveat of the new RT assay is that if the reaction is done in a CO2incubator, screw-cap tubes must be used to avoid lowering of the pH in the reaction, which sup-presses the RT activity (18b).
The RT assay has been used for the detection of known and novel retroviruses from infected cells both in assessing the replication of viruses and for evaluating antiretroviral treatments (e.g., in the case of treatments for HIV-1 infec-tion). We have used the RT assay described in this paper as a general detection strategy in a multicombinational analysis with specific detection strategies such as DNA and RNA PCR assays to demonstrate the absence of detectable HIV or SIV in several monovalent lots of oral, poliovirus vaccine (13). The new RT assay was especially useful because of its increased sensitivity for the detection of retroviral RT com-pared to those of other, similar assays and because of its low background signal.
ACKNOWLEDGMENTS
The following reagents were obtained through the AIDS Research and Reference Reagent Program, Division of AIDS, National Institute of Allergy and Infectious Diseases: p239SpE39and p239SpSp59cloned DNAs (from Ronald Desrosiers). We thank Teresa A. Galvin for preparing the titered SIVmac-mm239stock, S. Tabriz Ali for preparing
the titered HIV-1 stock, and Theodore Bryan for technical assistance. We also acknowledge Malcolm A. Martin for support in formulation of the RT cocktail and Keith Peden, Hana Golding, and Muhammad Shahabuddin for review of the manuscript.
REFERENCES
1.Baltimore, D.1970. RNA-dependent DNA polymerase in virions of RNA tumour viruses. Nature226:1209–1211.
2.Barre-Sinoussi, F., J. C. Chermann, F. Rey, M. T. Nugeyre, S. Chamaret, J.
Gruest, C. Dauguet, C. Axler-Blin, F. Vezinet-Brun, C. Rouzioux, W. Rosen-baum, and L. Montagnier.1983. Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS). Science220:868–871.
3.Cook, R. F., S. J. Cook, and C. J. Issel.1991. A nonradioactive micro-assay for released reverse transcriptase activity of a lentivirus. BioTechniques 13:380–386.
4.Daniel, M. D., N. W. King, N. L. Letvin, R. D. Hunt, P. K. Sehgal, and R. C. Desrosiers.1984. A new type D retrovirus isolated from macaques with an immunodeficiency syndrome. Science223:602–605.
5.Eberle, J., and R. Seibl.1992. A new method for measuring reverse tran-scriptase activity by ELISA. J. Virol. Methods40:347–356.
6.Gallo, R. C., S. Z. Salahuddin, M. Popovic, G. M. Shearer, M. Kaplan, B. F. Haynes, T. J. Palker, R. Redfield, J. Oleske, B. Safai, G. White, P. Foster, and P. D. Markham.1984. Frequent detection and isolation of cytopathic retroviruses (HTLV-III) from patients with AIDS and at risk for AIDS. Science224:500–503.
6a.Galvin, T. A., and A. S. Khan.Unpublished data.
7.Gronowitz, J. S., M. Neumuller, J. Lennerstrand, R. Bhikhabhai, T. Unge, H. Weltman, and C. F. R. Kallander.1991. Carrier bound templates for single tube reverse transcriptase assays and for combined purification and activity analyses, with special reference to HIV. Biotechnol. Appl. Biochem. 13:127–142.
8.Heberling, R. L., S. T. Barker, S. S. Kalter, G. C. Smith, and R. J. Helmke. 1977. Oncornavirus: isolation from a squirrel monkey (Saimiri sciureus) lung culture. Science195:289–292.
9.Heneine, W., S. Yamamoto, W. M. Switzer, T. J. Spira, and T. M. Folks. 1995. Detection of reverse transcriptase by a highly sensitive assay in sera from persons infected with human immunodeficiency virus type 1. J. Infect. Dis.171:1210–1216.
10. Hoffman, A. D., B. Banapour, and J. A. Levy.1985. Characterization of the AIDS-associated retrovirus reverse transcriptase and optimal conditions for its detection in virions. Virology147:326–335.
11. Kestler, H., T. Kodama, D. Ringler, M. Marthas, N. Pedersen, A. Lackner, D. Regier, P. Sehgal, M. Daniel, N. King, and R. Desrosiers.1990. Induction of AIDS in rhesus monkeys by molecularly cloned simian immunodeficiency virus. Science248:1109–1112.
12. Khan, A. S.1996. Retrovirus screening of vaccine cell substrates, p. 155–160. InF. Brown and A. S. Lubiniecki (ed.), Developments in biological stan-dardization: viral safety and evaluation of viral clearance from biopharma-ceutical products, vol. 88. Karger, Basel, Switzerland.
13. Khan, A. S., M. Shahabuddin, T. Bryan, B. H. Joshi, S. Lee, and I. K. Hewlett.1996. Analysis of live, oral poliovirus vaccine monopools for human immunodeficiency virus type 1 and simian immunodeficiency virus. J. Infect. Dis.174:1185–1190.
14. Kimpton, J., and M. Emerman.1992. Detection of replication-competent and pseudotyped human immunodeficiency virus with a sensitive cell line on the basis of activation of an integratedb-galactosidase gene. J. Virol.66: 2232–2239.
15. Layne, S. P., M. J. Merges, M. Dembo, J. L. Spouge, S. R. Conley, J. P. Moore, J. L. Raina, H. Renz, H. R. Gelderblom, and P. L. Nara.1992. Factors underlying spontaneous inactivation and susceptibility to neutraliza-tion of human immunodeficiency virus. Virology189:695–714.
16. Marx, P. A., D. H. Maul, K. G. Osborn, N. W. Lerche, P. Moody, L. J. Lowenstine, R. V. Henrickson, L. O. Arthur, M. Gravell, W. T. London, J. L. Sever, J. A. Levy, R. B. Munn, and M. B. Gardner.1984. Simian AIDS: isolation of a type D virus and disease transmission. Science223:1083–1086. 17. Pyra, H., J. Boni, and J. Schupach.1994. Ultrasensitive retrovirus detection by a reverse transcriptase assay based on product enhancement. Proc. Natl. Acad. Sci. USA91:1544–1548.
18. Reed, L. J., and H. A. Muench.1938. Simple method of estimating fifty per cent endpoints. Am. J. Hyg.27:493–497.
18a.Repaske, R.Data not shown. 18b.Repaske, R.Unpublished data.
19. Silver, J., T. Maudru, K. Fujita, and R. Repaske.1993. An RT-PCR assay for the enzyme activity of reverse transcriptase capable of detecting single viri-ons. Nucleic Acids Res.21:3593–3594.
20. Suzuki, K., T. Saito, M. Kondo, M. Osanai, S. Watanabe, T. Kano, K. Kano, and M. Imai.1995. Poly A-linked non-isotopic microtiter plate reverse tran-scriptase assay for sensitive detection of clinical human immunodeficiency virus isolates. J. Virol. Methods55:347–356.
21. Teich, N.1984. Taxonomy of retroviruses, p. 25–207.InR. Weiss, N. Teich, H. Varmus, and J. Coffin (ed.), RNA tumor viruses, molecular biology of tumor viruses, vol. 1, 2nd ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
22. Temin, H. M., and S. Mizutani.1970. RNA-dependent DNA polymerase in virions of Rous sarcoma virus. Nature (London)226:1211–1213.
on May 15, 2020 by guest
http://jcm.asm.org/
23.Varmus, H., and R. Swanstrom.1984. Replication of retroviruses, p. 369– 512.InR. Weiss, N. Teich, H. Varmus, and J. Coffin (ed.), RNA tumor viruses, molecular biology of tumor viruses, vol. 1, 2nd ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
24. Varmus, H., and R. Swanstrom.1985. Replication of retroviruses, p. 75–134. InR. Weiss, N. Teich, H. Varmus, and J. Coffin (ed.), RNA tumor viruses,
molecular biology of tumor viruses, vol. 2, 2nd ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
25. Willey, R. L., D. H. Smith, L. A. Lasky, T. S. Theodore, P. L. Earl, B. Moss, D. J. Capon, and M. A. Martin.1988. In vitro mutagenesis identifies a region within the envelope gene of the human immunodeficiency virus that is critical for infectivity. J. Virol.62:139–147.