Copyright © 2002, American Society for Microbiology. All Rights Reserved.
Genetic Classification and Differentiation of
Bartonella
Species Based
on Comparison of Partial
ftsZ
Gene Sequences
Zaher Zeaiter,
1Zhongxing Liang,
2and Didier Raoult
1*
Unite´ des Rickettsies, CNRS UMR 6020 IFR 48, Faculte´ de Me´decine, Marseille, France,
1and Division of Viral
and Rickettsial Diseases, National Center for Infections Diseases, Centers for Disease Control
and Prevention, Atlanta, Georgia 30333
2Received 25 April 2002/Returned for modification 9 June 2002/Accepted 15 July 2002
Currently, 19 species are recognized in the genus
Bartonella
, 7 of which are involved in an increasing variety
of human diseases. Development of molecular tools for detection, identification, and subtyping of strains and
isolates has promoted research on
Bartonella
spp. We amplified and sequenced the portion of the
ftsZ
gene
encoding the N-terminal region of the cell division protein for 13
Bartonella
species:
Bartonella alsatica
,
B.
birtlesii
,
B. doshiae
,
B. elizabethae
,
B. grahami
,
B. koehlerae
,
B. schoenbuchensis
,
B. taylorii
,
B. tribocorum
,
Bartonella vinsonii
subsp.
arupensis
,
Bartonella vinsonii
subsp.
berkhoffii
,
Bartonella vinsonii
subsp.
vinsonii
, and
B. bovis
Bermond et al.(“
B. weissii
”). Phylogenetically derived trees revealed four statistically supported groups,
indicating that sequencing of the
ftsZ
gene is a useful tool for identifying evolutionary relationships among
Bartonella
species. Furthermore, we amplified and sequenced the portion of the
ftsZ
gene encoding the
C-terminal region of the protein for 4
B. bacilliformis
isolates, 14
B. clarridgeiae
isolates, 14
B. quintana
isolates,
and 30
B. henselae
isolates that were obtained from different geographic regions, hosts, and clinical specimens.
B. clarridgeiae
and
B. quintana
sequences were highly conserved, while those of the four
B. bacilliformis
isolates
differed from the type strain at 5 positions. Among
B. henselae
strains isolated from cats and patients, only two
genotypes were detected: Houston and Marseille. Among 80 clinical samples we detected
Bartonella
spp. in 35
(43.75%) and found the assay to be comparable to that of a combined intergenic-spacer-region- and
pap31
-based PCR assay. Our results show the usefulness of the portion of the
ftsZ
gene encoding the C-terminal
region for diagnosis of
Bartonella
infections. More samples should be tested to study its usefulness for
epidemiological investigations.
The genus
Bartonella
contains aerobic, fastidious,
gram-neg-ative bacilli belonging to the alpha-2 subgroup of the class
Proteobacteria.
Recently the number of
Bartonella
species
iso-lated has increased markedly (5, 6, 15, 27), and the bacteria are
considered emerging pathogens involved in an increasing
num-ber of recognized diseases (1, 28, 38). Currently, 19
Bartonella
species are recognized, and all are associated with mammalian
hosts.
Bartonella taylorii
,
B. elizabethae
,
B. tribocorum
, and
B.
birtlesii
have been isolated from rats (6, 7, 11, 24);
B. grahamii
,
Bartonella vinsonii
subsp.
vinsonii
, and
B. doshiae
have been
recovered from voles (7, 11);
Bartonella vinsonii
subsp.
arupen-sis
has been isolated from mice (54);
B. alsatica
has been
isolated from rabbits (23);
B. koehlerae
,
B. clarridgeiae
,
B.
henselae
, and “
B. weissii
,” recently described as
B. bovis
Ber-mond et al., have been found in cats (5, 17, 29, 31, 35); and
B.
bovis
Bermond et al. has also been detected in cattle (5, 10).
Bartonella vinsonii
subsp.
berkhoffii
has been isolated from dogs
(10) and coyotes (13); “
B. washoensis
” has been demonstrated
in rodents (R. L. Regnery, personal communication);
B.
quin-tana
and
B. bacilliformis
have been isolated from humans (22,
40), and
B. schoenbuchensis
and
B. capreoli
have been isolated
from wild roe deer (5, 15). To date, 7 of the 19 species have
been implicated in human disease (28).
B. bacilliformis
is the
agent of bartonellosis (Carrion’s disease), which is endemic in
Andean valleys in South America.
B. quintana
and
B. henselae
,
etiologic agents of trench fever and cat scratch disease (CSD),
respectively, have also been associated with endocarditis and
bacillary angiomatosis in immunocompromised patients (1).
B.
elizabethae
and
B. vinsonii
subsp.
berkhoffii
cause endocarditis
(14, 46), and
B. vinsonii
subsp.
arupensis
was first isolated from
a febrile patient with heart valve disease in the United States
(54).
B. grahamii
has been implicated in cases of neuroretinitis
(30), and
B. clarridgeiae
is also suspected to be an agent of CSD
(32, 51). Because no distinguishing phenotypic characteristics
have been described for
Bartonella
species, their identification
and phylogenetic classification has been based mainly on
ge-netic studies. DNA hybridization and pulsed-field gel
electro-phoresis can be used for molecular characterization of
Bar-tonella
species (39, 47), but these techniques are not suitable
for routine use in a clinical laboratory. PCR-derived assays
allow detection and identification of the bacteria directly from
clinical samples even in conditions such as CSD, where
organ-isms are infrequently isolated in culture. Many DNA regions
and encoding gene sequences have been used in genetic
stud-ies: the 16S rRNA gene, the 16S–23S rRNA intergenic spacer
region (ITS) (26, 37), the citrate synthase gene (
gltA
) (8, 9, 25),
the riboflavin synthase alpha chain gene (
ribC
) (2), the heat
shock protein gene (
groEL
) (36, 55), the genes encoding the
PAP31 and 35-kDa proteins (33, 56), and the cell division
protein gene (
ftsZ
) (19, 29).
The FtsZ protein plays an important role in bacterial cell
division, and its gene sequence has been used to differentiate
three
Bartonella
species (29). Compared to other bacteria, the
* Corresponding author. Mailing address: Unite´ des Rickettsies,
CNRS UPRES-A 6020, Faculte´ de Me´decine, 27 boulevard Jean
Mou-lin, 13385 Marseille Cedex 05, France. Phone: (33) 4 91 32 43 75. Fax:
(33) 4 91 83 03 90. E-mail: [email protected].
3641
on May 15, 2020 by guest
http://jcm.asm.org/
FtsZ proteins of
Bartonella
species are nearly twice as large
and have an additional region at the C-terminal end (29, 42).
The C-terminal region has a higher degree of sequence
diver-gence than the N-terminal region and has recently been used
for
B. henselae
subtyping (19).
In our study we determined a partial 900-base nucleotide
sequence of
ftsZ
encoding the N-terminal region for the main
Bartonella
species and assessed its usefulness in species
differ-entiation and for inferring interspecies phylogenetic
relation-ships. Furthermore, we investigated PCR of the portion of the
ftsZ
gene encoding the C-terminal region as a means of
de-tecting and identifying
Bartonella
spp. in 80 clinical samples.
We also studied the usefulness of sequencing the portion of the
ftsZ
gene encoding the C-terminal end in subtyping
B.
hen-selae
,
B. quintana
,
B. clarridgeiae
, and
B. bacilliformis
isolates
from patients and cats and for epidemiological investigations
of infections.
MATERIALS AND METHODS
Bartonellastrains, isolates, and DNA extraction.Strains and isolates used in this study are detailed in Tables 1 and 2.Bartonellaisolates were grown on 5% sheep blood agar (Biomerieux, Marcy l’E´toile, France) at 37°C under a 5% CO2-enriched atmosphere. Bacteria were harvested after 7 days of culture, and
DNA was extracted by the Chelex method (52). Genomic DNA was stored at 4°C until use as a template in PCR assays.
PCR amplification and DNA sequencing of the portion of theftsZgene en-coding the N-terminal region.Primers (Eurobio, Les Ulis, France) used for amplification and sequencing are shown in Table 3. PCRs were carried out in a PTC-200 automated thermocycler (MJ Research, Waltham, Mass.) using an Elongase DNA polymerase kit (Gibco-BRL, Cergy Pontoise, France) and prim-ers Bfp1 and Bfp2 (Table 3). Reaction mixtures (25l) contained the following (final concentrations): primers (0.5 pmoll⫺1each), deoxynucleoside
triphos-phates (dATP, dCTP, dGTP, and dTTP) (0.2 mMl⫺1each), 1l of buffer A,
4l of buffer B, 0.6l of Elongase enzyme mix, 2.5l of DNA (150 to 200 ng), and sterile water. PCR amplifications were performed as follows: a 4-min
dena-turation at 94°C was followed by 44 cycles of denadena-turation for 30 s at 94°C, annealing for 30 s at 55°C, and extension for 60 s at 68°C. Amplification was completed by holding the reaction mixture at 68°C for 10 min to ensure complete extension of the PCR products. These were separated by electrophoresis on 1% agarose gels, visualized by staining with ethidium bromide, and purified with the QIAquick PCR purification kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. PCR products were sequenced in both directions using primers Bfp1, Bfp2, Bfs3, and Bfs4 and theD-Rhodamine Terminator
Cycle Sequencing Ready Reaction kit (Perkin-Elmer, Coignieres, France) ac-cording to the manufacturer’s instructions. Sequencing products were resolved using an ABI 3100 automated sequencer (Perkin-Elmer).
Analysis of sequences and construction of phylogenetic trees.Sequence anal-ysis was performed with ABI Prism DNA Sequencing Analanal-ysis Software, ver-sion 3.0 (Perkin Elmer), and multisequence alignment was performed with CLUSTAL W software, version 1.81 (53). DNA sequence similarities were cal-culated by use of MEGA 2.1 software (S. Kumar, K. Tamura, I. B. Jakobsen, and M. Nei, Molecular Evolutionary Genetics Analysis software, Tempe, Ariz., 2001). Phylogenetic trees were obtained from DNA sequences by using the maximum-parsimony method (DNAPARS software in PHYLIP) (20), distance methods (DNADIST [distance matrix with Kimura 2 parameters or Jukes-Can-tor parameters] and NEIGHBOR [neighbor joining]), and the maximum-likeli-hood method (DNAMLK software in PHYLIP). Bootstrap replicates were per-formed to estimate the node reliability of the phylogenetic trees obtained by the three methods (12). Bootstrap values were obtained from 100 trees (18) gener-ated randomly with SEQBOOT and CONSENSE in the PHYLIP software package. Only values above 90 were considered significant. Phylogenetic trees were established by using TreeView, version 1.5 (43). Only neighbor-joining trees are presented in this report. The phylogenetic trees we obtained were compared with those available forBartonellaspecies in GenBank, which were inferred from analyses of the 16S rRNA,gltA,rpoB, ITS, andgroELgene sequences.
PCR amplification and DNA sequencing of the portion of theftsZgene en-coding the C-terminal region.Primers used for amplification and sequencing of Bartonellaisolates and clinical samples are described in Table 3. PCR was carried out as described above by using primers BaftsZF and BaftsZR with 56°C as the annealing temperature. Sequencing was performed as described above by using primers BaftsZF, BaftsZR, BhftsZ 1393.n, Bh ftsZ 1247.p, Bq ftsZseqF, Bq ftsZseqR, Bb ftsZseqF, and Bb ftsZseqR. The resulting sequences from the differentBartonellaspecies were compared in order to investigate the usefulness of the C-terminal region in genotyping.
Clinical samples and DNA extraction.Eighty lymph node biopsy, lymph node aspirate, or valve samples from 79 patients with suspected CSD or endocarditis were sent to the Unite´ des Rickettsies to be tested for the presence ofBartonella spp. during December 2001. Thirty-nine samples had been found positive for Bartonellaspp. by use of ITS- andpap31-based PCR assays (47, 56). Ten to 25 mg of tissue or 200l of aspirate was used for extraction of total genomic DNA with the QIAamp tissue kit (Qiagen) according to the manufacturer’s instructions. Samples were handled under sterile conditions to avoid the risk of cross-con-tamination. Extracted DNA was suspended in 125l of elution buffer and stored at 4°C. DNAs from 10 bacterial strains and isolates were used as a negative control:Rickettsia helvetica,Escherichia coli,Mycobacterium tuberculosis, Pseudo-monas aeruginosa,Tropheryma whipplei,Afipia felis,Coxiella burnetii,Bosea mas-siliensis,Staphylococcus aureus, andEnterococcus faecalis.
For the PCR assays, samples were thawed at room temperature and amplified with primers FTS1p and FTS2p by using 56°C as the annealing temperature. Sterile distilled water was used in negative controls. A seminestedgroEL-derived assay was carried out (56) on samples for which discrepant results had been obtained in theftsZassay and the combined ITS/pap31assay, which was per-formed as previously described (47, 56).
Statistical analysis.Fisher’s exact test was used to compare the results of the combined ITS/pap31assay and theftsZassay. Observed differences were con-sidered significant when thePvalue was⬍0.05 for two-tailed tests.
RESULTS
Amplification of the portion of the
ftsZ
gene encoding the
N-terminal region for all the
Bartonella
species used in the
experiments yielded a single product of nearly 900 bp. Pairwise
comparison of these and the reported (Table 1)
ftsZ
sequences
revealed a sequence similarity ranging from 81.2 to 98.3%
(Table 4). When compared to the sequence similarities of
[image:2.587.42.285.92.307.2]other genes of
Bartonella
species available in GenBank,
ftsZ
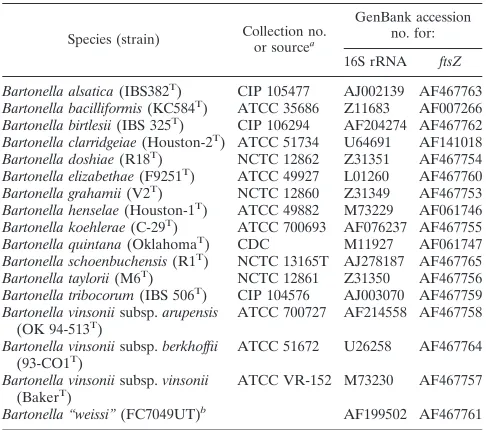
TABLE 1. Bacterial strains and sequences used for N-terminal
sequencing and phylogeny
Species (strain) Collection no.or sourcea
GenBank accession no. for: 16S rRNA ftsZ Bartonella alsatica(IBS382T) CIP 105477 AJ002139 AF467763
Bartonella bacilliformis(KC584T) ATCC 35686 Z11683 AF007266
Bartonella birtlesii(IBS 325T) CIP 106294 AF204274 AF467762
Bartonella clarridgeiae(Houston-2T) ATCC 51734 U64691 AF141018
Bartonella doshiae(R18T) NCTC 12862 Z31351 AF467754
Bartonella elizabethae(F9251T) ATCC 49927 L01260 AF467760
Bartonella grahamii(V2T) NCTC 12860 Z31349 AF467753
Bartonella henselae(Houston-1T) ATCC 49882 M73229 AF061746
Bartonella koehlerae(C-29T) ATCC 700693 AF076237 AF467755
Bartonella quintana(OklahomaT) CDC M11927 AF061747
Bartonella schoenbuchensis(R1T) NCTC 13165T AJ278187 AF467765
Bartonella taylorii(M6T) NCTC 12861 Z31350 AF467756
Bartonella tribocorum(IBS 506T) CIP 104576 AJ003070 AF467759
Bartonella vinsoniisubsp.arupensis
(OK 94-513T) ATCC 700727 AF214558 AF467758
Bartonella vinsoniisubsp.berkhoffii
(93-CO1T) ATCC 51672 U26258 AF467764
Bartonella vinsoniisubsp.vinsonii
(BakerT) ATCC VR-152 M73230 AF467757
Bartonella “weissi”(FC7049UT)b AF199502 AF467761
aAbbreviations: CIP, Collection de l’Institut Pasteur; Paris, France; ATCC, American Type Culture Collection, Manassas, Va.; NCTC, National Collection of Type Cultures, Central Public Health Laboratory, London, United Kingdom; CDC, Centers for Disease Control and Prevention, Atlanta, Ga.
bRecently described asB. bovisBermond et al.
on May 15, 2020 by guest
http://jcm.asm.org/
sequence similarity was found to be similar to those of
gltA
(83.4 to 96.1%),
rpoB
(85.9 to 96%), and
groEL
(83.1 to 98%),
higher than that of the ITS (69.1 to 99.7%), and lower than
that of the 16S ribosomal DNA (rDNA) (97.7 to 99.8%)
(Fig. 1).
Bartonella
phylogeny derived from
ftsZ
sequences.
For each
of the 17
Bartonella
species, a sequence of 788 bp could be used
for alignment and comparison. Phylogenetic trees derived by
using parsimony and distance methods showed consistent
[image:3.587.55.538.84.648.2]to-pologies and statistical support (Fig. 1). The
Bartonella
species
TABLE 2.
Bartonella
strains and isolates used for C-terminal amplification and sequencing
Species (strain) Clinical sourcea Geographic origin B. henselaegenotype
B. quintana(FullerT) Trench fever patient Yugoslavia
B. quintana(OklahomaT) Bacteremia, HIV-positive patient United States
B. quintana(URBQMTF 20) Blood culture France
B. quintana(URBQMTF 47) Bacteremia, homeless patient France
B. quintana(URBQMIE 48) Endocarditis patient France
B. quintana(URBQMTF 88) Blood culture France
B. quintana(URBQMTF 95) Bacteremia, homeless patient France
B. quintana(URBQMTF 96) Blood culture France
B. quintana(URBQMNHP 90) Body louse France
B. quintana(URBQMNHP 94) Body louse France
B. quintana(URBQMNHP 102) Body louse France
B. quintana(URBQMNHP 103) Body louse France
B. quintana(URBQMTF 108) Body louse France
B. henselaeMarseille (U8) Lymph node, CSD patient France Marseille
B. henselaeMarseille (U9) Endocarditis patient France Marseille
B. henselaeHouston-1 HIV-positive patient United States Houston
B. henselae(SA-2) Suspected CSD patient United States Houston
B. henselae(90-615) Lymph node, CSD patient United States Houston
B. henselae(CAL-1) Septicemia United States Marseille
B. henselaeEN1 Endocarditis patient France Marseille
B. henselaeEN2 Endocarditis patient France Marseille
B. henselaeEN3 Endocarditis patient France Marseille
B. henselae1129 Endocarditis patient France Houston
B. henselae5327 Endocarditis patient France Marseille
B. henselae5097 Endocarditis patient France Houston
B. henselaeC20 Cat France Houston
B. henselaeC45 Cat France Houston
B. henselaeC51 Cat France Marseille
B. henselaeC52 Cat France Marseille
B. henselaeC53 Cat France Marseille
B. henselaeC77 Cat France Houston
B. henselaeC78 Cat France Houston
B. henselaeC85 Cat France Houston
B. henselaeC87 Cat France Houston
B. henselaeC96 Cat France Houston
B. henselaeNZ1 Cat of patient with CSD Australia Marseille
B. henselaeNZ2 Cat of patient with CSD Australia Marseille
B. henselaeNZ3 Cat of patient with CSD Australia Marseille
B. henselaeNZ4 Cat of patient with CSD Australia Marseille
B. henselaeNZ5 Cat of patient with CSD Australia Marseille
B. henselaeNZ6 Cat of patient with CSD Australia Marseille
B. henselaeNZ7 Cat of patient with CSD Australia Houston
B. henselaeNZ8 Cat of patient with CSD Australia Marseille
B. bacilliformisT Bartonellosis patient Peru
B. bacilliformisCuzco 8 Bartonellosis patient Peru
B. bacilliformisCuzco 14 Bartonellosis patient Peru
B. bacilliformisCuzco 269 Bartonellosis patient Peru
B. bacilliformisCuzco 812 Bartonellosis patient Peru
B. clarridgeiae Cat United States
B. clarridgeiaeC23 Cat France
B. clarridgeiaeC44 Cat France
B. clarridgeiaeC48 Cat France
B. clarridgeiaeC49 Cat France
B. clarridgeiaeC53 Cat France
B. clarridgeiaeC68 Cat France
B. clarridgeiaeC69 Cat France
B. clarridgeiaeC71 Cat France
B. clarridgeiaeC73 Cat France
B. clarridgeiaeC74 Cat France
B. clarridgeiaeC75 Cat France
B. clarridgeiaeC76 Cat France
B. clarridgeiaeC157 Cat France
aHIV, human immunodeficiency virus.
on May 15, 2020 by guest
http://jcm.asm.org/
were divided into two clades with significant bootstrap values
(99%); the first contained
B. birtlesii
,
B. schoenbuchensis
, and
B. bovis
Bermond et al. in one arm and
B. clarridgeiae
and
B. bacilliformis
in the second arm. The second clade contained
three clusters. All three
B. vinsonii
subspecies (
B. vinsonii
subsp.
vinsonii
,
B. vinsonii
subsp.
arupensis
, and
B. vinsonii
subsp.
berkhoffii
) grouped together in the first cluster (100%);
the second contained
B. henselae
,
B. koehlerae
, and
B. quintana
(99%); the third contained
B. elizabethae
,
B. tribocorum
, and
B. grahamii
(99%). The branching of
B. taylorii
,
B. alsatica
, and
B. doshiae
in the different groups was not reliable (70, 46, and
32%, respectively).
Comparison of the sequences of the
ftsZ
gene encoding the
C-terminal region for subtyping
Bartonella
species isolates.
A
fragment of nearly 885 bp of the portion of the
ftsZ
gene
encoding the C-terminal region was amplified from four
iso-lates of
B. bacilliformis
, 14 of
B. clarridgeiae
, 14 of
B. quintana
,
and 30 of
B. henselae
. The sequences of all the fragments were
compared with one another and with those available in
Gen-Bank (Table 2). The sequences of each of the 14
B. quintana
isolates were identical to one another and to that previously
described for
B. quintana
(Oklahoma) (29). The sequences of
all
B. clarridgeiae
isolates used were identical to that reported
for
B. clarridgeiae
(GenBank accession no. AF141018). While
the sequences of all four of our
B. bacilliformis
isolates were
identical (accession no. AF467752), they differed from that of
B. bacilliformis
Tat 5 positions: 1071, 1279, 1490, 1587, and
1676 (numbered relative to the
ftsZ
gene of
B. bacilliformis
,
accession no. AF007266). Variation at the first and fourth of
these positions yielded silent mutations. Two previously
de-scribed genotypes (4, 16, 19, 49, 50, 55, 56) were detected
among the
B. henselae
isolates we tested (Table 2). The
Hous-ton sequence (accession no. AF161249) was found in 43.3% of
our isolates; the remainder were of the Marseille genotype
(accession no. AF161251).
Use of
ftsZ
C-terminal-derived primers for detection and
identification of
Bartonella
spp. directly from clinical samples
and comparison of their efficiency with that of the combined
ITS/
pap31
PCR assay.
All negative controls gave no PCR
products. When the 80 clinical samples which had previously
been tested with the combined ITS/
pap31
PCR assay (41, 48)
[image:4.587.49.540.84.239.2]were assayed, C-terminal
ftsZ
amplicons were detected in 35
TABLE 3. Primers used for PCR and/or sequencing
Primera Bartonellaspecies Primer sequence Source or
reference
Bfp1 (ap,s)
All
5⬘-ATTAATCTGCAYCGGCCAGA-3⬘
This study
Bfp2 (ap,s)
All
5⬘-ACVGADACACGAATAACACC-3⬘
This study
Bfs3 (s)
All
5⬘-TTACAAAAATCYGTTGATAC-3⬘
This study
Bfs4 (s)
All
5⬘-GTATCAACRGATTTTTGTAA-3⬘
This study
BaftsZF (ap,s)
All
5⬘-GCTAATCGTATTCGCGAAGAA-3⬘
This study
BaftsZR (ap,s)
All
5⬘-GCTGGTATTTCCAAYTGATCT-3⬘
This study
BhftsZ 1393.n (s)
B. henselae, B. clarridgeiae
5⬘-GCGAACTACGGCTTACTTGC-3⬘
19
Bh ftsZ 1247.p (s)
B. henselae, B. clarridgeiae
5⬘-CGGTTGGAGAGCAGTTTCGTC-3⬘
19
Bq ftsZseqF (s)
B. quintana
5⬘-GCACATATTCTTGATGAGAT-3⬘
This study
Bq ftsZseqR (s)
B. quintana
5⬘-CCCCTATCATCTCATCAAG-3⬘
This study
Bb ftsZseqF (s)
B. bacilliformis
5⬘-GCGCATGTTCTTAGTGAAAT-3⬘
This study
Bb ftsZseqR (s)
B. bacilliformis
5⬘-CCTGTATACGTGATGCATTT-3⬘
This study
FTS1p (ap,s)
All
5⬘-GCCTTCTCATCCTCAACTT-3⬘
This study
FTS2p (ap,s)
All
5⬘-CAGCCTCTTCACGATGTG-3⬘
This study
aap, amplification primer; s, sequencing primer.
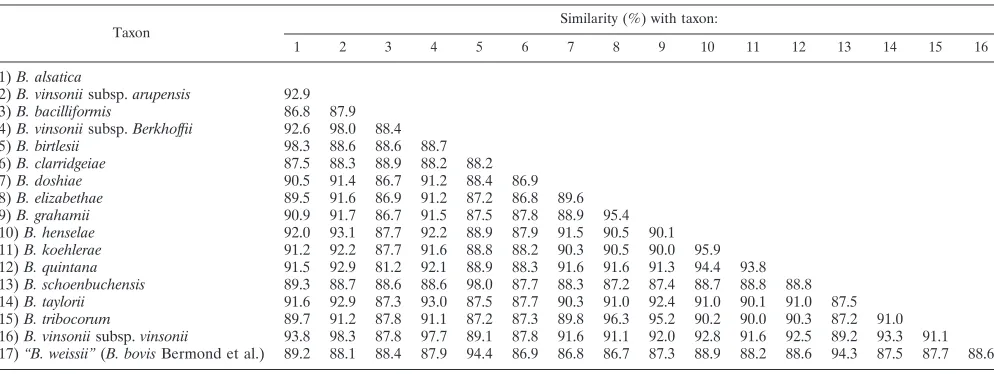
TABLE 4. Level of
ftsZ
DNA sequence similarity for
Bartonella
species
Taxon Similarity (%) with taxon:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
1)B. alsatica
2)B. vinsoniisubsp.arupensis 92.9 3)B. bacilliformis 86.8 87.9 4)B. vinsoniisubsp.Berkhoffii 92.6 98.0 88.4
5)B. birtlesii 98.3 88.6 88.6 88.7
6)B. clarridgeiae 87.5 88.3 88.9 88.2 88.2
7)B. doshiae 90.5 91.4 86.7 91.2 88.4 86.9
8)B. elizabethae 89.5 91.6 86.9 91.2 87.2 86.8 89.6 9)B. grahamii 90.9 91.7 86.7 91.5 87.5 87.8 88.9 95.4 10)B. henselae 92.0 93.1 87.7 92.2 88.9 87.9 91.5 90.5 90.1 11)B. koehlerae 91.2 92.2 87.7 91.6 88.8 88.2 90.3 90.5 90.0 95.9 12)B. quintana 91.5 92.9 81.2 92.1 88.9 88.3 91.6 91.6 91.3 94.4 93.8 13)B. schoenbuchensis 89.3 88.7 88.6 88.6 98.0 87.7 88.3 87.2 87.4 88.7 88.8 88.8 14)B. taylorii 91.6 92.9 87.3 93.0 87.5 87.7 90.3 91.0 92.4 91.0 90.1 91.0 87.5 15)B. tribocorum 89.7 91.2 87.8 91.1 87.2 87.3 89.8 96.3 95.2 90.2 90.0 90.3 87.2 91.0 16)B. vinsoniisubsp.vinsonii 93.8 98.3 87.8 97.7 89.1 87.8 91.6 91.1 92.0 92.8 91.6 92.5 89.2 93.3 91.1 17)“B. weissii”(B. bovisBermond et al.) 89.2 88.1 88.4 87.9 94.4 86.9 86.8 86.7 87.3 88.9 88.2 88.6 94.3 87.5 87.7 88.6
on May 15, 2020 by guest
http://jcm.asm.org/
[image:4.587.44.541.543.730.2]samples (43.75%) from 34 patients. The overall correlation
between the C-terminal
ftsZ
assay results and those of the
ITS-and
pap31
-derived assay was 89.7%, but this was not significant
(
P
⫽
0.052). Four samples were negative by the
ftsZ
assay but
positive with the ITS-
pap31
assay; three of these samples were
also positive in the seminested
groEL
-derived assay (56).
DISCUSSION
In the past decade, a number of new
Bartonella
species have
been described (6, 15, 27) and comparisons of 16S rDNA
sequences have led to many taxonomic changes in the genus
Bartonella
(7, 11). Although comparison of 16S rDNA gene
sequences has been useful in phylogenetic studies at the genus
level (41), its use has been questioned in studies at the species
level (21; M. Hasegawa and T. Hashimoto, Letter, Nature
361:
23, 1993). Other genes have been used empirically in
at-tempts to classify the
Bartonella
species: the
gltA
gene (9), the
rpoB
gene (45), the 16S–23S rRNA ITS (26), and the
groEL
(55) gene. The FtsZ protein plays an important role in
bacte-rial cell division, and recently its sequence was established for
four
Bartonella
species (29). In our study we sequenced the
900-base sequence encoding the N-terminal region (partial) of
the
ftsZ
gene for all recognized
Bartonella
species. The
se-quences were generally well conserved (81.2 to 98.3% [Table
4]) between species, but the sequence divergence present
al-lowed us to develop a phylogenetic tree (Fig. 1) which was well
supported for most of the strains studied. We compared this
tree with those inferred from sequences of the 16S rDNA,
gltA
,
rpoB
, ITS, and
groEL
genes of
Bartonella
species available in
GenBank (Fig. 1). With the
ftsZ
sequences, the
Bartonella
species were divided into two well supported clades which were
also obtained with the
groEL
and
rpoB
sequences. Within these
clades, various supported clusters could be found with the
different DNA sequences. The statistically supported cluster
formed by the subspecies of
B. vinsonii
in the
ftsZ
tree was also
found in the
rpoB
-, ITS-, and
groEL
-derived trees. A cluster
containing
B. henselae
and
B. koehlerae
was found in the
ITS-derived tree, while a cluster containing
B. henselae
and
B.
quintana
was obtained in the
rpoB
- and
groEL
-derived trees. A
cluster including
B. tribocorum
,
B. elizabethae
, and
B. grahamii
was present in the phylogenetic trees established by using the
gltA
,
rpoB
, ITS, and
groEL
sequences.
B. taylorii
was included
in this cluster in the ITS- and
groEL
-derived trees. A cluster
formed by
B. bovis
Bermond et al. and
B. birtlesii
was found in
the
groEL
-derived tree, and a cluster of
B. bovis
Bermond et al.
and
B. schoenbuchensis
was found in the 16S rDNA-inferred
tree. The similarities we found between the phylogenetic trees
derived with the
ftsZ
gene sequences and those derived with
other genes shows that
fstZ
gene sequencing should be
[image:5.587.57.532.75.342.2]consid-ered a useful tool to be included in phylogeny studies.
We believe that it is important to consider the sequences of
several genes in phylogeny studies. Although each
gene-de-rived tree will differ from the others and will have different
FIG. 1. Comparison of neighbor-joining trees based on 16S rDNA,
gltA
,
rpoB
, ITS,
groEL
, and
ftsZ
partial or complete sequences. Bootstrap
values at tree nodes are based on 100 replicates; values of
⬎90 are boldfaced. Trees were unrooted, and only topology was shown for these trees.
a
,
gltA
, citrate synthase;
rpoB
, beta subunit of RNA polymerase; ITS, 16S–23S rRNA ITS;
groEL
, heat shock protein;
ftsZ
, cell division protein.
b, range of the level of DNA sequence similarity for each gene used. Designations for species and subspecies consist of the letter
B
(for
Bartonella
)
and the following abbreviations:
ber
,
vinsonii
subsp.
berkhoffii
;
vin
,
vinsonii
subsp.
vinsonii
;
aru
,
vinsonii
subsp.
arupensis
;
tri
,
tribocorum
;
eli
,
elizabethae
;
gra
,
grahamii
;
tay
,
taylorii
;
als
,
alsatica
;
dos
,
doshiae
;
hen
,
henselae
;
qui
,
quintana
;
koe
,
koehlerae
;
cla
,
clarridgeiae
;
bir
,
birtlesii
;
sho
,
schoenbuchensis
;
bac
,
bacilliformis
;
bov
,
bovis
Bermond et al.
on May 15, 2020 by guest
http://jcm.asm.org/
levels of statistical support, it has been found that groupings
obtained with two different sequences at bootstrapping values
over 90% are stable and reliable (48). In previous phylogenetic
studies
B. bacilliformis
was chosen to be the outgroup, but
because new
Bartonella
species have been described recently
(6, 15) we chose to draw an unrooted tree.
Because
Bartonella
species are implicated in an increasing
variety of human diseases, the development of species-specific
tools for their detection and identification in clinical samples is
becoming more crucial, especially in light of the difficulties in
culturing these bacteria (34). In 1996, Drancourt et al. (16)
reported two serotypes of
B. henselae
(Houston and Marseille),
and later Bergmans et al. (4) confirmed by 16S rDNA gene
sequence analysis that there were two genotypes of
B. henselae
,
genotypes I and II, corresponding to the Houston and
Mar-seille serotypes, respectively. More recently, further studies
have confirmed the presence of these two subspecies (3, 4, 19,
33, 49, 50, 56). Many genes have been used to characterize
Bartonella
isolates (2, 8, 9, 27, 36, 37, 55), and the cell division
protein (FtsZ) has also been used for detection (29) and
sub-typing of
Bartonella
species (19). In our study we amplified and
sequenced the
ftsZ
sequence corresponding to the C-terminal
region for 4
B. bacilliformis
isolates, 14
B. clarridgeiae
isolates,
14
B. quintana
isolates, and 30
B. henselae
isolates from
differ-ent geographic regions, hosts, and clinical samples. The
se-quences of the
B. clarridgeiae
and
B. quintana
isolates were
identical to those of the type strains. Similarly, sequencing of
the
groEL
gene could not be used to differentiate
B. quintana
isolates (55). When the ITS sequence was used for subtyping,
however,
B. quintana
isolates were found to belong to three
genotypes and different sequences were found for all the
B.
clarridgeiae
isolates studied (26). This difference may be
ex-plained by the high degree of variability of ITS sequences. The
ftsZ
sequence data may show the homogeneity of the
B.
clar-ridgeiae
and
B. quintana
isolates. The sequences of the four
B.
bacilliformis
isolates we studied were identical to one another
but different from that of
B. bacilliformis
Tat 5 positions, only
3 of which yielded significant amino acid substitutions. Among
the 30
B. henselae
isolates we studied, only Houston and
Mar-seille genotypes were found and there was no evidence of
genotype III, detected by Ehrenborg et al. (19).
We also tested whether the C-terminal
ftsZ
assay could
de-tect the DNAs of
Bartonella
species in clinical samples and
compared its sensitivity with that of a combined ITS-
pap31
assay. We believe that false-positive PCR results due to
con-tamination problems may be prevented by using a number of
primer pairs which target different genes. Addition of the
ftsZ
gene to the panel of genes available for diagnosis of infections
by PCR may be useful, and it may be a good tool for the
“suicide” PCR application (44).
Conclusion.
We confirmed that using one pair of primers
enables the comparison of partial
ftsZ
sequences for all
Bar-tonella
species and that this is a useful tool for detection and
identification, which should facilitate routine work on clinical
samples. The sequences obtained were also useful in
phyloge-netic analyses at the species level, and the results obtained
correlated closely with those obtained in previous studies using
other markers. Furthermore, we showed that
Bartonella
spe-cies occur in two clades and that
B. bacilliformis
belongs to a
robust and well-defined clade. Using multiple DNA sequences
seems to be the most suitable way to reliably infer phylogeny.
We also showed the usefulness of
ftsZ
C-terminal region
se-quencing in the direct detection and identification of
Bar-tonella
species in clinical samples and for subtyping
B. henselae
and
B. bacilliformis
isolates. Its usefulness for epidemiological
studies should be further investigated by using a diverse range
of clinical samples.
ACKNOWLEDGMENTS
We thank Yves Piemont for providing the
B. schoenbuchensis
strain,
Jennifer Robson for providing
B. henselae
isolates from Australian
cats, and Pat Kelly for reviewing the manuscript.
REFERENCES
1.Anderson, B. E., and M. A. Neuman. 1997.Bartonellaspp. as emerging human pathogens. Clin. Microbiol. Rev.10:203–219.
2.Bereswill, S., S. Hinkelmann, M. Kist, and A. Sander. 1999. Molecular analysis of riboflavin synthesis genes inBartonella henselaeand use of the ribC gene for differentiation ofBartonellaspecies by PCR. J. Clin. Microbiol. 37:3159–3166.
3.Bergmans, A. M. C., C. M. A. de Jong, G. Van Amerongen, C. S. Schot, and L. M. Schouls.1997. Prevalence ofBartonellaspecies in domestic cats in The Netherlands. J. Clin. Microbiol.35:2256–2261.
4.Bergmans, A. M. C., J. F. P. Schellekens, J. D. A. Van Embden, and L. M. Schouls.1996. Predominance of twoBartonella henselaevariants among cat scratch disease patients in The Netherlands. J. Clin. Microbiol.34:254–260. 5.Bermond, D., H. J. Boulouis, R. Heller, G. Van Laere, H. Monteil, B. B. Chomel, A. Sander, C. Dehio, and Y. Piemont.2002.Bartonella bovis Ber-mond et al. sp.nov. andBartonella capreolisp. nov., isolated from European ruminants. Int. J. Syst. Evol. Microbiol.52:383–390.
6.Bermond, D., R. Heller, F. Barrat, G. Delacour, C. Dehio, A. Alliot, H. Monteil, B. Chomel, H. Boulouis, and Y. Pie´mont.2000.Bartonella birtlesii sp. nov., isolated from small mammals (Apodemusspp.). Int. J. Syst. Evol. Microbiol.50:1973–1979.
7.Birtles, R. J., T. G. Harrison, N. A. Saunders, and D. H. Molyneux.1995. Proposals to unify the generaGrahamellaandBartonella, with descriptions ofBartonella talpaecomb. nov.,Bartonella peromyscicomb. nov., and three new species,Bartonella grahamiisp. nov.,Bartonella tayloriisp. nov., and Bartonella doshiaesp. nov. Int. J. Syst. Bacteriol.45:1–8.
8.Birtles, R. J., S. Hazel, K. Brown, D. Raoult, M. Begon, and M. Bennett. 2000. Subtyping of uncultured bartonellae using sequence comparison of 16S/23S rRNA intergenic spacer regions amplified directly from infected blood. Mol. Cell. Probes14:79–87.
9.Birtles, R. J., and D. Raoult.1996. Comparison of partial citrate synthase gene(gltA)sequences for phylogenetic analysis ofBartonellaspecies. Int. J. Syst. Bacteriol.46:891–897.
10.Breitschwerdt, E. B., and D. L. Kordick.2000.Bartonellainfection in ani-mals: carriership, reservoir potential, pathogenicity, and zoonotic potential for human infection. Clin. Microbiol. Rev.13:428–438.
11.Brenner, D. J., S. O’Connor, H. H. Winkler, and A. G. Steigerwalt.1993. Proposals to unify the generaBartonellaandRochalimaea, with descriptions ofBartonella quintanacomb. nov.,Bartonella vinsoniicomb. nov.,Bartonella henselaecomb. nov., andBartonella elizabethaecomb. nov., and to remove the familyBartonellaceaefrom the orderRickettsiales. Int. J. Syst. Bacteriol. 43:777–786.
12.Brown, J. K. M.1994. Bootstrap hypothesis tests for evolutionary trees and other dendrograms. Proc. Natl. Acad. Sci. USA91:12293–12297. 13.Chang, C. C., R. W. Kasten, B. B. Chomel, D. C. Simpson, C. M. Hew, D. L.
Kordick, R. Heller, Y. Piemont, and E. B. Breitschwerdt.2000. Coyotes (Canis latrans)as the reservoir for a human-pathogenicBartonellasp.: mo-lecular epidemiology ofBartonella vinsoniisubsp.berkhoffiiinfection in coy-otes from central coastal California. J. Clin. Microbiol.38:4193–4200. 14.Daly, J. S., M. G. Worthington, D. J. Brenner, W. C. Moss, D. G. Hollis, R. S.
Weyant, A. G. Steigerwalt, R. E. Weaver, M. I. Daneshvar, and S. P. O’Connor.1993.Rochalimaea elizabethaesp. nov. isolated from a patient with endocarditis. J. Clin. Microbiol.31:872–881.
15.Dehio, C., C. Lanz, R. Pohl, P. Behrens, D. Bermond, Y. Pie´mont, K. Pelz, and A. Sander.2001.Bartonella schoenbuchiisp. nov., isolated from the blood of wild roe deer. Int. J. Syst. Evol. Microbiol.51:1557–1565. 16.Drancourt, M., R. Birtles, G. Chaumentin, F. Vandenesch, and D. Raoult.
1996. New serotype ofBartonella henselaein endocarditis and cat-scratch disease. Lancet347:441–443.
17.Droz, S., B. Chi, E. Horn, A. G. Steigerwalt, A. M. Whitney, and D. J. Brenner.1999.Bartonella koehleraesp. nov., isolated from cats. J. Clin. Microbiol.37:1117–1122.
18.Efron, B., E. Halloran, and S. Holmes.1996. Bootstrap confidence levels for phylogenetic trees. Proc Natl. Acad. Sci USA93:13429–13434. (Original printing,93:7085–7090.)
on May 15, 2020 by guest
http://jcm.asm.org/
19.Ehrenborg, C., L. Wesslen, A. Jakobson, G. Friman, and M. Holmberg.2000. Sequence variation in theftsZgene ofBartonella henselaeisolates and clin-ical samples. J. Clin. Microbiol.38:682–687.
20.Felsenstein, J.1989. PHYLIP—phylogeny inference package (version 3.2). Cladistics5:164–166.
21.Fox, G. E., J. D. Wisotzkey, and P. J. Jurtshuk.1992. How close is close: 16S rRNA sequence identity may not be sufficient to guarantee species identity. Int. J. Syst. Bacteriol.42:166–170.
22.Gray, G. C., A. A. Johnson, S. A. Thornton, W. A. Smith, J. Knobloch, P. W. Kelley, E. L. Obregon, H. M. Arones, and F. S. Wignall.1990. An epidemic of Oroya fever in the Peruvian Andes. Am. J. Trop. Med. Hyg.42:215–221. 23.Heller, R., M. Kubina, P. Mariet, P. Riegel, G. Delacour, C. Dehio, F. Lamarque, R. Kasten, H. J. Boulouis, H. Monteil, B. Chomel, and Y. Pie-mont.1999.Bartonella alsaticasp. nov., a newBartonellaspecies isolated from the blood of wild rabbits. Int. J. Syst. Bacteriol.49:283–288. 24.Heller, R., P. Riegel, Y. Hansmann, G. Delacour, D. Bermond, C. Dehio, F.
Lamarque, H. Monteil, B. Chomel, and Y. Pie´mont.1998.Bartonella tribo-corumsp. nov., a newBartonellaspecies isolated from the blood of wild rats. Int. J. Syst. Bacteriol.48:1333–1339.
25.Houpikian, P., P. E. Fournier, and D. Raoult.2001. Phylogenetic position of Bartonella vinsoniisubsp.arupensisbased on 16S rDNA andgltAgene se-quences. Int. J. Syst. Evol. Microbiol.51:179–182.
26.Houpikian, P., and D. Raoult.2001. 16S/23S rRNA intergenic spacer regions for phylogenetic analysis, identification, and subtyping ofBartonellaspecies. J. Clin. Microbiol.39:2768–2778.
27.Houpikian, P., and D. Raoult.2001. Molecular phylogeny of the genus Bartonella: what is the current knowledge? FEMS Microbiol. Lett.200:1–7. 28.Jacomo, V., P. J. Kelly, and D. Raoult.2002. Natural history ofBartonella infections (an exception to Koch’s postulate). Clin. Diagn. Lab. Immunol. 9:8–18.
29.Kelly, T. M., I. Padmalayam, and B. R. Baumstark.1998. Use of the cell division protein FtsZ as a means of differentiating amongBartonellaspecies. Clin. Diagn. Lab. Immunol.5:766–772.
30.Kerkhoff, F. T., A. M. Bergmans, Z. van Der, and A. Rothova.1999. Dem-onstration ofBartonella grahamiiDNA in ocular fluids of a patient with neuroretinitis. J. Clin. Microbiol.37:4034–4038.
31.Koehler, J. E., C. A. Glaser, and J. W. Tappero.1994.Rochalimaea henselae infection: a new zoonosis with the domestic cat as a reservoir. JAMA271: 531–535.
32.Kordick, D. L., E. J. Hilyard, T. L. Hadfield, K. H. Wilson, A. G. Steigerwalt, D. J. Brenner, and E. B. Breitschwerdt.1997.Bartonella clarridgeiae, a newly recognized zoonotic pathogen causing inoculation papules, fever, and lymph-adenopathy (cat scratch disease). J. Clin. Microbiol.35:1813–1818. 33.La Scola, B., Z. Liang, Z. Zeaiter, P. Houpikian, P. Grimont, and D. Raoult.
2002. Genotypic characteristics of two serotypes of Bartonella henselae. J. Clin. Microbiol.40:2002–2008.
34.La Scola, B., and D. Raoult.1999. Culture ofBartonella quintanaand Bar-tonella henselaefrom human samples: a 5-year experience (1993 to 1998). J. Clin. Microbiol.37:1899–1905.
35.Lawson, P. A., and M. D. Collins.1996. Description ofBartonella clarridgeiae sp. nov. isolated from the cat of a patient withBartonella henselaesepticemia. Med. Microbiol. Lett.5:64–73.
36.Marston, E. L., J. W. Sumner, and R. L. Regnery. 1999. Evaluation of intraspecies genetic variation within the 60-kDa heat-shock protein gene (groEL) ofBartonellaspecies. Int. J. Syst. Bacteriol.49:1015–1023. 37.Matar, G. M., B. Swaminathan, S. B. Hunter, L. N. Slater, and D. F. Welch.
1993. Polymerase chain reaction-based restriction fragment length polymor-phism analysis of a fragment of the ribosomal operon fromRochalimaea species for subtyping. J. Clin. Microbiol.31:1730–1734.
38.Maurin, M., R. J. Birtles, and D. Raoult.1996. A review of bartonellae and
their infections, p. 587–610.InJ. Kazar and R. Toman. (ed.), Rickettsiae and rickettsial diseases. Veda, Bratislava, Slovakia.
39.Maurin, M., V. Roux, A. Stein, F. Ferrier, R. Viraben, and D. Raoult.1994. Isolation and characterization by immunofluorescence, sodium dodecyl sul-fate-polyacrylamide gel electrophoresis, Western blot, restriction fragment length polymorphism-PCR, 16S rRNA gene sequencing, and pulsed-field gel electrophoresis ofRochalimaea quintanafrom a French patient with bacil-lary angiomatosis. J. Clin. Microbiol.32:1166–1171.
40.McNee, J. W., A. Renshaw, and E. H. Brunt.1916. “Trench fever”: a relaps-ing fever occurrrelaps-ing with the British forces in France. Br. Med. J.12:225–234. 41.Olsen, G. J., and C. R. Woese.1993. Ribosomal RNA: a key to phylogeny.
FASEB J.7:113–123.
42.Padmalayam, I., B. Anderson, M. Kron, T. Kelly, and B. Baumstark.1997. The 75-kilodalton antigen ofBartonella bacilliformisis a structural homolog of the cell division proteinftsZ. J. Bacteriol.179:4545–4552.
43.Page, R. D.1996. TreeView: an application to display phylogenetic trees on personal computers. Comput. Appl. Biosci.12:357–358.
44.Raoult, D., G. Aboudharam, E. Crubezy, G. Larrouy, B. Ludes, and M. Drancourt.2000. Molecular identification by “suicide PCR” ofYersinia pestis as the agent of medieval Black Death. Proc. Natl. Acad. Sci. USA97:12800– 12803.
45.Renesto, P., J. Gouvernet, M. Drancourt, V. Roux, and D. Raoult.2001. Use ofrpoBgene analysis for detection and identification ofBartonellaspecies. J. Clin. Microbiol.39:430–437.
46.Roux, V., S. J. Eykyn, S. Wyllie, and D. Raoult.2000.Bartonella vinsonii subsp.berkhoffiias an agent of afebrile blood culture-negative endocarditis in a human. J. Clin. Microbiol.38:1698–1700.
47.Roux, V., and D. Raoult.1995. Inter-and intraspecies identification of Bar-tonella (Rochalimea)species. J. Clin. Microbiol.33:1573–1579.
48.Roux, V., and D. Raoult.2000. Phylogenetic analysis of members of the genus Rickettsia using the gene encoding the outer-membrane protein rOmpB (ompB). Int. J. Syst. Evol. Microbiol.50:1449–1455.
49.Sander, A., C. Bu¨hler, K. Pelz, E. Von Cramm, and W. Bredt.1997. Detec-tion and identificaDetec-tion of twoBartonella henselaevariants in domestic cats in Germany. J. Clin. Microbiol.35:584–587.
50.Sander, A., M. Ruess, K. Deichmann, N. Bo¨hm, and W. Bredt.1998. Two different genotypes ofBartonella henselaein children with cat-scratch disease and their pet cats. Scand. J. Infect. Dis.30:387–391.
51.Sander, A., A. Zagrosek, W. Bredt, E. Schiltz, Y. Piemont, C. Lanz, and C. Dehio.2000. Characterization ofBartonella clarridgeiaeflagellin (FlaA) and detection of antiflagellin antibodies in patients with lymphadenopathy. J. Clin. Microbiol.38:2943–2948.
52.Stein, A., and D. Raoult.1992. A simple method for amplification of DNA from paraffin-embedded tissues. Nucleic Acids Res.20:5237–5238. 53.Thompson, J. D., D. G. Higgins, and T. J. Gibson.1994. CLUSTAL W:
improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res.22:4673–4680.
54.Welch, D. F., K. C. Carroll, E. K. Hofmeister, D. H. Persing, D. A. Robison, A. G. Steigerwalt, and D. J. Brenner.1999. Isolation of a new subspecies, Bartonella vinsonii subsp.arupensis, from a cattle rancher: identity with isolates found in conjunction withBorrelia burgdorferiandBabesia microti among naturally infected mice. J. Clin. Microbiol.37:2598–2601. 55.Zeaiter, Z., P. E. Fournier, H. Ogata, and D. Raoult.2002. Phylogenetic
classification ofBartonellaspecies by comparinggroELsequences. Int. J. Syst. Evol. Microbiol.52:165–171.
56.Zeaiter, Z., P. E. Fournier, and D. Raoult.2002. Genomic variation of Bartonella henselaedetected in lymph nodes from patients with cat scratch disease. J. Clin. Microbiol.40:1023–1030.