0022-538X/81/080348-11$02.00/0
Specific
Sindbis Virus-Coded Function for Minus-Strand RNA
Synthesis
DOROTHEA L.SAWICKI,`* STANLEY G. SAWICKI,' SIRKKAKERANEN,2ANDLEEVI
KAARIAINEN2
Departmentof Microbiology, MedicalCollegeofOhio, Toledo, Ohio 43614,1 and Department of Virology, University of Helsinki, 00290Helsinki 29,Finland2
Received29January1981/Accepted20April1981
Thesynthesis of minus-strand RNAwasstudiedincell cultures infected with
the heat-resistant strain of Sindbis virus and with temperature-sensitive (ts)
mutants belonging to complementation groups A, B, F, and G, all of which
exhibitedanRNA-negative(RNA-) phenotype when infectionwasinitiated and maintainedat390C,thenonpermissivetemperature.When infected cultureswere
shiftedfrom28°C (thepermissive temperature)to39°Cat 3hpostinfection,the
synthesis of viral minus-strand RNA ceased in cultures infected withts mutants
ofcomplementationgroupsB andF, but continued in cultures infected with the parental virus and mutants ofcomplementation groups A and G. In cultures infected with tsll of complementation group B, the synthesis of viral minus-strand RNA ceased, whereas the synthesis of 42S and 26S plus-strand RNAs continued for atleast5 hafter theshift to39°C. However, when tsll-infected cultureswerereturnedto280C 1haftertheshiftto39°C, the synthesis of viral minus-strand RNA resumed, and therateof viral RNA synthesis increased. The
recovery of minus-strand synthesis required translation of new proteins. We
conclude thatatleastoneviralfunction isrequired for alphavirus minus-strand synthesis that isnotrequired for plus-strand synthesis. Incultures infected with
ts6 of complementation group F, the syntheses of both viral plus-strand and
minus-strand RNAswere drastically reduced after the shift to 390C. Since ts6
failed tosynthesize both plus-strand and minus-strand RNAs after the shiftto
390C,
at least one common viral component appears to be required for thesynthesis ofbothminus-strand andplus-strandRNAs.
As one of the initial steps in the infectious
cycle of the alphaviruses Sindbis virus (SIN)
and Semliki Forest virus, the parental42S ge-nomeRNAistranslated toyield aviral
RNA-dependentRNA polymerase whichutilizes the parental42Splus-strandRNA as atemplate for the synthesis of 42S minus-strand RNA. The 42Sminus-strandRNA in turn serves as a
tem-plate for the synthesis ofmore42S plus-strand
RNA and ofsubgenomic 26S mRNA,which is
translated into the viralstructuralproteins (15,
22, 29). During thefirst 3 hpostinfection (p.i.),
the rate ofsynthesisofplus-strand and
minus-strand RNAs increases; however, at
approxi-mately 3 to 3.5 h p.i., the synthesis of minus
strands ceases abruptly, whereas the synthesis
of both42S and 26S plus strandscontinues at a
constantratethroughout the infectious cycle (3,
25). During the early period of infection, the
synthesis of plusstrands occurs in about a
five-fold excess relative to minus-strand RNA
syn-thesis (25).
The RNA-dependent RNA polymerase
re-sponsibleforthesynthesisof 26S and 42S
plus-strand RNAsmustbestable,since theaddition
ofinhibitors ofproteinsynthesis after3h
post-infection does not significantly interfere with
plus-strand RNA synthesis (13, 25, 26, 30). We
haverecently shown that thesynthesis of minus-strand RNA in Semliki Forest virus-infected
cellsceaseswithin 15to20minafter the addition
of inhibitorsofprotein synthesis(25). Thus, the polymeraseresponsibleforminus-strand
synthe-sisdiffers from the viralpolymerasesynthesizing
plus-strandRNAbyhavingashorthalf-life.We have suggested (25) thatalphaviruses regulate
the rateoftranscription ofviral RNAby
regu-lating the number of minus-strand templates
and that thesynthesisofminus-strand RNA is
regulatedatthelevel of translationbya
mech-anism which utilizes one or more short-lived
polymerase proteins. One possible mechanism could involve an unstable or rapidly
turning-overpolymerase proteinthat isrequired onlyfor
minus-strandsynthesis.Wereport in this
com-municationtheidentification ofa temperature-348
on November 10, 2019 by guest
http://jvi.asm.org/
sensitive(ts) mutantof the heat-resistantstrain
of SIN (SIN HR) that selectively ceased
minVs-strand syntheis upon shiftingtothe
nonpermis-sivetemperature.
MATERIALS AND METHODS Cell cultures. Secondarycultures of chicken em-bryofibroblast (CEF) cells were prepared from pri-mary cultures of 10-day-old embryos obtained from the eggs ofleukosis-free flocks and were grown in plastic petri dishes inDulbecco modified Eagle mini-mal essential medium (DMEM) supplemented with 5% fetal bovine serum and 5% tryptose phosphate broth. BHK-21 cells, a continuous cell line derived from baby hamster kidneycells, were grown in plastic petri dishes in DMEM supplemented with 5% fetal bovineserum.
Virus. SIN HR and theRNA-negative (RNA-) ts mutantsused in thisstudywerethe generousgift of E. R. Pfefferkorn(Dartmouth MedicalSchool, Hano-ver,N.H.) and have been characterized previously (5, 6, 22,29).Growth andpurification of each virus stock anddetermination of its infectivity by plaque assayat both the permissive (280C) and the nonpermissive
(390C)temperatureswere aspreviouslydescribed(16). Infection and RNAlabeling.In all of the experi-mentspresented, with the exception of those in Fig.1 andTable 1,weemployedCEFmonolayers in 50-mm petri dishes infected with either SIN HRor oneof the mutants at a multiplicity of infection (MOI) of50. After themonolayerwaswashed with Hanks balanced salt solutionat 28°C, the viruswasadsorbedto the CEF cells for1hat280Cinaninoculum of0.5 to 1.0 ml ofcompleteDMEM (DMEM containing0.2% bo-vine serum albumin, 22 mM HEPES
[N-2-hydroxy-ethylpiperazine-N'-2-ethanesulfonic acid, pH 7.4],and
1 ug of actinomycin D per ml). At the end of the adsorption period, thevirus-containing medium was removed,complete DMEM at280C wasadded,and the cultures were incubated at 280C for the times indicated in thetext.Toaccomplishthe shiftto390C,
themonolayerswerewashed withHanks balanced salt
solution at 390C before the addition of complete
DMEMat390C.When culturesat390Cwerereturned
to280C,theywerewashedoncewithHanks balanced salt solution at 280C, followed by the addition of complete DMEMat280C.SIN RNAwaslabeled with
[5,6-3H]uridineat afinalconcentration of250,uCi/ml
per 50-mm petri dish, unless otherwise indicated in thetext.
Sinceourstudies involved the incubation of infected cellsat twodifferent temperatures,wedetermined the kinetics ofSIN RNAsynthesisatthesetwo tempera-tures. At various times after infection, SIN HR-in-fectedcells were labeled for60min with[3H]uridine, and the cultures were solubilized with 2% sodium
dodecylsulfate in ET buffer(0.01 MEDTA,0.01 M
Tris-hydrochloride [pH 7.4]). DNA was sheared by
passageseveraltimesthrougha27-gauge needle. Sam-ples were taken for protein determination by the method ofLowryetal.(17) and for total acid-insoluble radioactivity. Wefound, ashave others (15, 16, 22), that theentirereplicationcycletakesapproximately
twiceaslong when incubationisat280Cthanwhen it
is at 390C. When the maximum number of transcrip-tion complexes had formed and RNA synthesis was at aconstant rate(by 4 to 5 h at390Cand by 8 to 9 h at 280C), [3H]uridine incorporation into RNA during a 1-hpulse-label in cultures maintained at390Cwas twice that obtained from cultures maintained at280C(data not shown). Therefore, to adjust for the effect of temperature on the observed rate of transcription, a pulse of 60 min was given to cultures to be labeled at
280C,and a 30-min pulse was used for cultures labeled
at390C.
Isolation of SIN RF RNA. At the times indicated, the infected cells were washed once with ice-cold phosphate-buffered saline,followed by the addition of 2ml of lysis buffer (0.15 M NaCl, 0.01 M Tris-hydro-chloride [pH 7.4], 1.5 mM MgCl2) containing 0.65% Triton X-100. The cells were scraped from the surface of the petri dish and transferred to 15-ml conical centrifuge tubes; the solution was vigorously blended inaVortex mixer before being centrifuged at 250 x g for10minat40Ctopellet the nuclei. The cytoplasmic supernatantwasremoved and adjusted to 1% sodium dodecyl sulfate. The solution was extracted twice with phenol, followed by two extractions with chloroform-isoamyl alcohol (96:4). RNA was precipitated from the aqueousphase by the addition of LiCl to 0.2 M and ethanol to 70%. After overnight storage at-200C, RNAwascollectedby centrifugation in an SW41 rotor at98,000xgfor1hat0°C. The dried precipitate was dissolved in0.2ml ofdigestion buffer (0.15 M NaCl, 0.05 M Tris-hydrochloride [pH 6.8], 1 mM EDTA) anddigested with0.1 ugof pancreatic RNase perml atroomtemperaturefor15min.A solution of 0.05 M Tris-hydrochloride (pH 6.8) and 1 mM EDTA was addedtolower theNaCl concentration to 0.1 M, and ethanolwasthen addedto afinal concentration of 35% (vol/vol). The samples were immediately applied to columns of CF-11cellulose by the procedure of Frank-lin(11)for isolation ofreplicative form (RF) RNA.
CF-11 cellulose columnswerepoured in 3-ml plastic syringes (2.5 ml of packed CF-11cellulose) andwere washed first with modifed STE buffer (0.1 M NaCl, 0.05 MTris-hydrochloride [pH 6.8], 10 mMEDTA) containing 1%beta-mercaptoethanol, followedby ex-tensivewashing with STE buffer (0.1 M NaCl,0.05M Tris-hydrochloride[pH6.8],1mMEDTA) containing 35%ethanol. Thesampleswereappliedtothecolumns and washed with10 to 15mlof STE buffercontaining
35% ethanol, followed by an equal volume of STE
buffercontaining 15%ethanol;RFRNAwaseluted in STEbuffer. We routinely recovered 2 to5% of the
applied radioactivityasRF RNA. RF RNAwas
col-lectedby ethanolprecipitationinthe presence of0.2 MLiCl and25 Lgof carrierratliver RNA.
Thisprocedure quantitatively recovered RF RNA from cellsinfected with SIN HR or tsmutantsof SIN HR. Thesameamountof RF RNAwasobtainedby either thesucrose gradient method used previously (24, 25) or the CF-11 cellulose method of Franklin (11). Furthermore, bothofthesepreparationsofRF RNAcontainedthesameproportionoflabeled minus-strand RNA andsedimentedat15Sto20Sinsucrose
gradients. RFRNAisolatedbytwocyclesofsucrose
gradientcentrifugation (24, 25)boundtoCF-11
cellu-lose, and 100% ofappliedRF RNAwasrecoveredin
on November 10, 2019 by guest
http://jvi.asm.org/
350
the STE buffer fraction. Less than 0.1% of labeled
single-stranded viral RNAwasfound to elute in the
STE buffer fraction after treatment with this concen-trationofpancreatic RNase.CellularDNA eluted in STEbuffercontaining15%ethanol and thus could be efficiently removed from the RF RNA fraction.
Quantitationof minus-strand RNAsynthesis.
Minus-strandRNAwasmeasuredby determiningthe
amountof[3H]uridineincorporatedintoRF RNA that hybridized to unlabeled 42S plus-strand RNA. The conditions of hybridization were identical to those reported byuspreviously (24, 25).Labeled RNA that
wasprotected frompancreatic RNasedigestionwas
takentobelabeled minus-strandRNA. Itshould be noted that this minus-strand RNArepresentedthat whichremainedaftertreatmentof thereplicative in-termediates with low levels of pancreatic RNase to obtain RF RNA.Such RNasedigestionwouldremove
any single-stranded, nascent minus-strand or
plus-strand RNA in thereplicativeintermediates. Materials. [5,6-3H]uridine (53 Ci/mmol)was pur-chased from Amersham Radiochemicals, London, England. Cycloheximide was obtained from Calbi-ochem, LaJolla,Calif.,andCF-11 cellulose was ob-tained from Whatman, Inc., Clifton, N.J. All other materialswerefrompreviouslydescribedsources (23-25).
RESULTS
Blocking of minus-strandRNAsynthesis
in SIN HR-infected cells by inhibition of
proteinsynthesis.Figure1showsthe effectof
blocking translation on the synthesis of viral
RNA(Fig.1A) and minus-strand RNA(Fig. 1B)
incells infected with SIN HR. In untreatedcells,
the overallrateofviralRNAsynthesisincreased
exponentially beginning at 1 to 1.5 h p.i. and
continued until3.5 to4hp.i., afterwhichtime it becameconstantorslowlydecreased(Fig.1A).
The rateofsynthesisof minus-strand RNA
in-creasedatthesametime that the overallrateof viral RNAsynthesis increased and ceasedatthe time that theoverallrateofviral RNAsynthesis becameconstant(Fig. 1B). When cycloheximide
wasaddedduring theperiodin which therates
of synthesis of viral plus-strand and
minus-strand RNAswereincreasing,theoverallrateof synthesis of viral plus-strand RNA stopped in-creasing and became constant or slowly declined
(Fig. 1A), and the synthesis of minus-strand
RNA ceased within 30 min aftertheaddition of
cycloheximide (Fig. 1B). Cycloheximidehad no
effectontheoverallrateofviral RNAsynthesis
whenit wasadded after the nornal cessation of
minus-strand synthesis had occurred and after
the overall rate of viral RNA synthesis had
become constant (data not shown). These
re-sults,which werethesameregardlessofwhether
CEForBHK-21cellswereusedfortheinfection,
demonstrated thatminus-strandtranscriptionin
SIN-infected cells was shut off selectively by
to
0
E
oU
z
2
/
1E-II 2 _
en I I I I I I
-J B
WU120
a
10
8
64
6 CL
2
HOURS p.1.
FIG. 1. Rate of SIN HR RNA synthesis in the presence and absence of cycloheximide. BHK-21 monolayers in50-mmpetridisheswereinfected with SIN HR(MOI, 50) andweremaintainedat37°Cin the presenceofactinomycin D. At2hp.i., medium containingcycloheximide (100pg/mi)wasadded to one setofcultures(arrow). Bothsetsof cultures were
pulsedwith[3H]uridine(250yiCi/ml)at37°Cfor
30-minperiodsbetween2and5.5hp.i. and were
har-vestedimmediatelyafter the pulse. (A) Total [3H]-uridineincorporationinto viral RNA. Acid-insoluble radioactivitywasdetermined by counting a sample
aftertrichloroacetic acidprecipitation. The values
shown represent the total
[3Hluridine
in eachsam-ple; duplicate experiments gave similar results. (B)
Total[3H]uridine incorporation into minus-strand RNA. RF RNA in each sample was isolated by CF-11cellulosechromatography as described in the text, denaturedby heatingat1000C,and allowed to rean-neal in the presenceof an excess of unlabeled SIN HR 42S virion RNA. RNase-resistant,radiolabeled
RNAformedduring the hybridization reaction was
taken as labeled minus-strand RNA. Symbols: 0, untreatedcultures;0,cultures treated with cyclohex-imide.
cycloheximidein amanner identical to that
ob-served in BHK-21,HeLa, or Vero cells infected
withSemlikiForest virus (25). Thesynthesisof
alphavirus minus-strand RNA, therefore,
ap-pears torequirethecontinued synthesis of
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.503.275.430.66.372.2]SINDBIS VIRUS MINUS-STRAND 351
teins. To determine whether aviral protein is
responsible for minus-strand transcription, we
screenedseveraltsmutantsof SIN HRto
iden-tifymutantspossessingatsdefect thatmanifests
itself in the failure to synthesize minus-strand
RNA, butnotplus-strand RNA, atthe
nonper-missivetemperature.
Screening for SIN HRts mutantsthatare
defective in minus-strand RNA synthesis.
We screened sevenrepresentatives of SIN HR
tsmutantsthathavebeenassignedtofour
com-plementationgroupsthat giveanRNA-
pheno-type. These RNA- mutants fail to synthesize
detectableamountsofviral RNAwhen the
in-fection isinitiated and maintainedat390C,the
nonpermissive temperature. To identify ts
mu-tantsofSIN HR thatareselectively unableto
synthesize minus-strand RNAatthe
nonpermis-sive temperature, the cells mustbe infected at
the permissive temperature. The infected cells
arethenshiftedtothenonpermissive
tempera-ture at a time when minus-strand RNA
tran-scription is detectable. Thus, the RNA-mutants
canbecharacterizedby their abilityorinability
to continue synthesizing plus-strandor
minus-strand RNA. Nascent viralRNAand all
minus-strand RNAsarefoundaspartofthe replicative
intermediates. During apulse-label with
[3H]-uridine,theproportion of radiolabel incoporated
intominus-strand RNA relativetothat in
plus-strand RNA in the replicative intermediates
in-dicates the relative level ofsynthesis of
minus-strand RNA. During the period in which minus
strandsarebeingaccumulated, 40to50% of the
[3H]uridineincorporated into RF RNA derived
from replicative intermediates is in minus
strands, and this proportion isconstantforpulse
periods of 15 minto4 h(24, 25).
Monolayers of CEF cells were infected with
either SIN HR orRNA- tsmutantsand were
maintainedat280Cfor 3 h beforebeing shifted
to39°C.After30minat390C,
[3H]uridine-con-tainingmediumwasadded to thecultureswhich
weremaintainedat390Cforanadditional 2.5 h
before harvest.Viraldouble-stranded RFRNA
wasisolated, RNAwasdenaturedby heatingat
1000C, and the proportion of labeled
minus-strand RNA in RF RNA was determined by
hybridization in the presence of an excess of
unlabeled SIN HRvirion RNA.Table 1 shows
three tsmutantsthatappearedtobe unableto
continue synthesis of minus-strand RNA after
the shift to 390C: ts4, ts6, and tsll. RF RNA
isolated from tsll-infected cells contained the
least proportion of labeled minus-strand RNA
(about 6%);RFRNA isolated from ts4- and
ts6-infected cells containedonlyabout 10%labeled
minus-strand RNA. Theseresultswere in
con-trastwiththoseobtainedwith RF RNA isolated
TABLE 1. Analysis of SIN RNA- mutants for defects in minus-strandsynthesisa
Complementa- RN
ineedR
Virus°tiopn
grmoeunp
RNA inmu'nus-tionroup strand RNA
SIN HR 40.4
ts4 A 9.8
ts6 F 8.8
ts7 G 20.3
tsll B 6.2
tsl5 A 24.6
tsl8 G 31.6
ts24 A 36.2
aMonolayers of CEF cells in 50-mm petri dishes
wereinfected with either SIN HR or the ts mutants asdescribed in the text. An MOI of 50 was used for the B and G group mutants, and an MOI of 100 was used for the A and F group mutants. SIN HR was tested at both MOIs and gave equivalent results for minus-strand synthesis. The different MOI was used to maximize incorporation into ts4 and ts6 RNAs because of the reported effect of the shift to390Con RNAsynthesis (16). At 3 h p.i., the monolayers were shifted to 39°C and were further incubated at 39°C in fresh medium. Beginning 30 min after the shift to
390C,[3H]uridine-containingmedium was added (500
,uCi/2ml per petri dish), and the incubation continued for2.5h.Theinfectedcellextracts wereprepared, and RNase-resistant RF RNA was isolated and hybridized to an excessof unlabeled 42S virion RNA. The hy-bridization values shown represent the average of from two to five determinations. A control preparation of RF RNA from SIN HR-infected cells labeled with
[3H]uridinefrom1 to 5hp.i.at370Ccontained 49% of
thelabeled RNA in the RFs in minus-strand RNA.
from theremainingmutants (20to36% labeled
minusstrands) and SINHR(40%labeled minus strands).
A closer examination of these mutants was
undertaken.Pulses of[3H]uridineweregivenfor
30min atthe time oftheshiftto390Candat
30-minintervalsthereaftertocultures infected with
SIN HRmutantsts4,ts6,tsll,andtsl5and with
theparentalSINHR. Therateof RNA
synthe-sis inSIN HR-infected cells increased
approxi-mately 20-foldoverthe 3-hperiodafter the shift
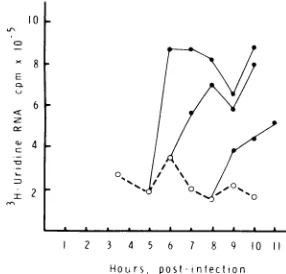
to390C (Fig.2). However, inallof the
mutant-infected cultures, raising the temperature to 390C at3 hp.i.resulted inafailuretoincrease
substantially the rate of RNA synthesis. The
rate of RNAsynthesisintsll-infected cultures
increasedonlytwo-tothreefoldoverthis
period,
remained essentially constant in ts4- and
tsl5-infected cultures, and slowly decreased in
ts6-infected cultures.Weexamined theother
RNA-mutantsshown inTable1,and allof them failed
toincreasesignificantlytherateofRNA
synthe-sisafterthe shiftto390Cat3 hp.i.;their rate of
synthesis either was constant or slightly
de-creased over the 3-h period after the shift to
on November 10, 2019 by guest
http://jvi.asm.org/
352
20
15
L-I
2n
I0
5
3 4 5 6
[image:5.503.64.226.55.402.2]Hours,
post-infectionFIG. 2. Relative RNA synthesis in SIN HR- and mutantvirus-infected culturesat39°C. CEFcells in 50-mmpetridisheswereinfected with SINHRor one
ofthets mutants atanMOIof50andmaintainedat
28°Cuntil 3h p.i.,atwhich timetheywereshiftedto
39°C. [3H]uridine incorporation intoRNAat39°C
wasdetermined with pulses of [3H]uridine(250 ,iCi/ ml) for30min between3and 6 h p.i. The cellswere
harvestedasdescribed inthetext,and sampleswere
takenfor acid-insolubleradioactivity. Theamountof incorporation observedin cultures pulsedbetween3 and3.5hp.i. (shownbelow in parentheses)wastaken
as1.0, andtheamountof incorporationobserved in thesubsequentpulse periods foreachmutantand the parental virus is expressedrelativeto thisamount.
During the infectious cycle, [3H]uridine incorpora-tion intoSINHR RNAduringa30-minpulse-label
increasedbyanaverageof 22-fold (fiveexperiments ranging from 9- to 33-fold). Cultures wereinfected
with: A,ts4(133, 181 cpm);A,ts6(183, 848 cpm);0,
tsll(273,600cpm);x,ts15(230,854cpm);*,SIN HR
(241, 270 cpm); and EL SINHR incubated in the
presenceofcycloheximidefrom the timeof the shiftto
39°C (241,029cpm).
390Cat3hp.i. (datanotshown).The inability
ofthe RNA- mutants to increase significantly
thelevel ofviral RNA synthesisafter the shift
to 390C at 3 h p.i. was not unexpected. The
failuretoincrease therateof viral RNA
synthe-siscouldhave resulted fromafailure to
synthe-sizeadditionalminus-strandtemplates,afailure
to accumulate functional polymerase
compo-nents,and theinactivation ofpreviously formed
polymerase components that are temperature sensitive inthese mutants. Therefore, the rela-tive level of minus-strand RNA synthesis was
determinedfromRF RNA isolated from each of
the datumpointsshown in Fig. 2
(Fig.
3).Be-tween 40 and 50% of RF RNA
synthesized
inSIN HR-infected cells between1 and 2 hafter
theshiftto390Chybridizedtounlabeled virion RNA anddecreasedtoless than 1%by3h after
theshift.Therefore,theseresults
again
showed that theincreasingrateof SIN HRplus-strand
RNAtranscriptioncorrelated with thesynthesis
ofminus-strand RNA. Cells infected with SIN
tsl5 of theAcomplementiongroup continuedto
synthesizeminus-strandRNAforanadditional
3hafter the shiftto390C(Fig. 3).Similar results
havebeen obtained for cells infected with SIN
ts18 of the G complementation group and for
SINts24of theA
complementation
group(data
notshown). Unlike the resultsobtained forSIN
HR,tsl5, ts18,andts24,
only
asmallproportion
60
5 50
40
3
40
E
C - 20
E
u 0
aI 0 A
0
3 4 5 6
[image:5.503.276.423.358.580.2]Hours, post- i nfection
FIG. 3. Minus-strand RNA synthesis in infected CEF culturesshifted to390C.RF RNA wasisolated fromextractsof thecultures shown in Fig. 2 byCF-I1 cellulosechromatography. The percentage of [3H]-uridineincorporatedintoRF RNA thatwas in minus' strands was determined by hybridization as de-scribedin thetext. RF RNA was isolated from cul-turesinfected with: *, SIN HR;A,ts4;A, ts6;0,tsll; U, tsl5.
on November 10, 2019 by guest
http://jvi.asm.org/
SINDBIS VIRUS MINUS-STRAND MUTANT 353
oflabeled RF RNA obtained from ts4-,ts6-,and
tsll-infected cells after the shift to390Cwasin
minusstrands(Fig. 3). Thus, minus-strand
syn-thesis ceased in ts4-, ts6-,and tsll-infectedcells
soon after the shift to the nonpermissive
tem-perature.
Analysisofplus-strandRNAsynthesis in
toll-infected cells.Wefocused our studies on SIN HRtsll,thesolemember of the B
comple-mentation group, that is, the only mutant in
whichminus-strand synthesisappears to be
se-lectively temperature sensitive. Although
mi-nus-strandsynthesis failed to continue at 390C
ineither ts4- orts6-infectedcells (Fig. 3), the ts
defectdoes not appear toberestricted to
minus-strand synthesis, but also directlyaffects
plus-strand RNA synthesis. Ineither ts4- or ts6-in-fected cells, thesynthesis of42Sand 26S
plus-strand RNAs is reducedafter the shift to390C atlate timesintheinfectiouscycle when
minus-strand transcription has already ceased (16).
This is incontrast totsll-infected cells,in which
42S and 26Splus-strandsynthesescontinueafter
the shiftto390C atlate timesin theinfectious cycle (16). We analyzed the pattem and the
amountsof42Sand26Splus-strandRNAs
syn-thesized in tsll-infected cells after the shift to 390Cat atime (3hp.i.) intheinfectious cycle
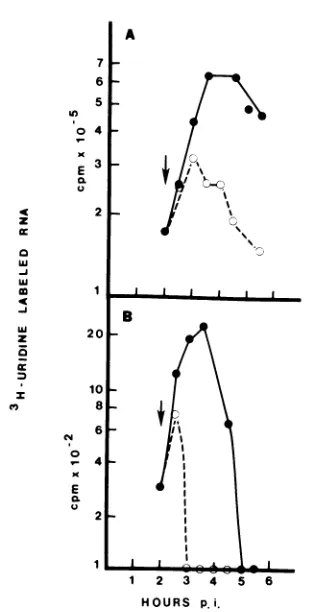
whenminus-strand synthesiswasjustbeginning. CulturesofCEF cellswereinfectedat280Cwith
tsll andwere either maintained at
280C
(Fig.4A) or shiftedat 3hp.i. to390C (Fig. 4B) and
labeled with[3H]uridine.At bothtemperatures,
full-length42S and 26Splus-strandRNAs were
synthesized. The cultures that hadbeenshifted
to390C incorporated 18% as much [3H]uridine
into42S and26S RNAsasdid the culture
main-tained at 280C, but an amount equal to that
obtained in a culture maintained at 280C but
labeled at 3 h p.i. (data not shown).
Further-more,the ratio oflabeled 26S RNAto42S RNA
did notchangesignificantly withashift in
tem-perature; at280Cthe ratiowas1.7,andat390C
the ratio was 1.2. Therefore, plus-strand RNA
continued to be synthesized at
390C
intsll-infectedcells, although minus-strand
transcrip-tion ceased when the cultures were shifted to
thenonpermissivetemperature at 3h p.i. Analysis of viral RNA synthesis at the
permissivetemperature. Wenextdetermined
whethertheregulationofminus-strand
synthe-sis inSINHRtsll-infectedcellswasnormalat
the permissive temperature. Cultures of CEF
cells were infected with tsll at280C and were
pulsedfor 60 min with
[3H]uridine
atintervalsthroughout infection (Fig. 5A). An increasing
amount of
[3H]uridine
incorporation into RNAwasdetected between2and 6 hp.i.,followedby
several hours of a more orlessconstant rateof
x
E 8
Z 6
wc
E 4
t'l 2
As"s.
2
B 42 265
IU0 U0 U0 U0 30
IRACTION NUMBER
FIG. 4. Sucrosegradientanalysis of RNA synthe-sized intsll-infectedcultures at thepermissive and nonpermissive temperatures. Monolayers of CEF cellswereinfected withtsllat28°C. Certain of these culturesweremaintainedat28°C,receivinga60-min pulse of [3H]uridine atvarious intervals; other cul-tures wereshiftedto39°Cat3 hp.i. andwere main-tained at39°C. Cellswereharvestedafter the pulse into ETbuffercontaining 2% sodium dodecyl sulfate, and theextracts werelayeredonto15 to30%sucrose gradients in NETbuffer (0.1 MNaCI,0.01M EDTA, 0.01 MTris-hydrochloride[pH 7.4]) containing 0.2% sodiumdodecyl sulfate and sedimented inanSW27.1 rotor at 93,000xgfor18hat20°C. Samples of 0.5 ml were collected, and the radioactivity in each was determinedby counting the entirefraction afteracid precipitation and collectionon aglassfiberfilter. (A) tsll-infected culture maintainedat28°C and pulsed with [3H]uridine between 7and8 hp.i.; (B) tsll-infected culture shifted to 39°C at 3 hp.i., subse-quently maintainedat39°C, andpulsed with [3H]-uridine between 7.5 and8hp.i.
incorporation.RF RNAwasisolated and showed asimilarpatternofsynthesis(Fig.5B).As shown inFig.5C,theperiodofincreasingRNA synthe-siswasassociatedwithanincrease in the
incor-poration of radiolabel into minus-strandRNA.
The insert to Fig. 5C shows the proportion of
the totalincorporation in RF RNA thatwasin
minus strands. The synthesis of minus-strand
RNA occurred at an increasing rate at 280C
until after 4 h p.i., at which time it declined:
between 2 and 3 h and 3 and 4 hp.i.,about 50%
of the radiolabel in RF RNA was in minus
strands;by6p.i.,less than 20% of labeled RNA
was in minus strands; and only 3% was found
between 6and 7hp.i. Thus, atthepermissive
temperature, the synthesis of minus-strand
RNA intsll-infected cellswastemporally
reg-ulatedandwassimilartothatseenin the
paren-tal SIN HR-infected cells (Fig. 1). Since
plus-strand RNA continuedtobesynthesizedfor at
least 5 h after the shift to 390C (Fig. 4), the
previously formed minus strands continued to
function astemplates for
plus-strand synthesis
I11--1 -Ld baft..AgnoL1-D -AL
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.503.262.460.58.224.2]354 SAWICKI ET AL.
0
E
aL
z
-J
z
CO) 0
E
0
E
,'so
_,so
h.0
I *E20
.1
03 4 5 * 7 9
hours p.i.
HOURS p.j.
FIG. 5. Rateof[3H]uridine incorporation into SINtsll RNA at28°C andaftertheshiftto39°C. CEF
monolayers were infected with tsll (MOI, 50) andmaintained at28°C. Beginningat2hp.i., apulseof
[3HJuridinewasgiven for60-minperiodsat28°C. Duplicate cultureswereshiftedto39°Cat 3hp.i. andwere
pulsed with[3HJuridinefor 30-min periods beginningatthe timeof the shift. (A) Total incorporation into viraltsllRNA; (B) total incorporation into RF RNA thatwasisolatedfromthe extract of each ofthe samples shown in(A); (C) total incorporation intotsll minus-strand RNA. RF RNA was denatured and allowedto reannealin the presenceofan excessof unlabeled 42S virion RNAasdescribed in the text. The insert shows thesameresultsas those shown in(C) expressedas apercentageof the total labeled RF RNA that is in minus-strand RNA. Symbols: 0, cultures maintainedat28°C; 0, cultures shiftedto39°Cat3hp.i. and maintainedat39°C.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.503.152.372.70.559.2]after tsll-infected cellswere shifted from 28 to
390C
at 3 h p.i. For a direct comparison withevents at280C, duplicatecultures wereshifted
to 390C at 3 h p.i. and similarly analyzed. An
initial two- tothreefold rise in plus-strand
syn-thesiswasfollowed by a slowdecline, so that 7
h after the shift, 40% of the initial rate was
observed (Fig. 5A).Aconstant amount of
incor-porationwasdetectedin RF RNA (Fig.5B),and
as seenpreviously(Fig. 3), minus-strand
synthe-sisceased about 60minafter the shift to 390C
(Fig. 5C).
Resumption of minus-strand synthesis.
Since raising thetemperature to390C resulted
in the cessation of minus-strand synthesis in
tsll-infected cells, we next asked whether
mi-nus-strandsynthesis wouldresume after a return tothepermissivetemperature. The requirement
fortranslation ofnew viralproteins for the
re-sumption of minus-strand transcriptionupon a
shift down from39 to
280C
wasinvestigated todistinguish between the reactivation of previ-ously formed proteins and the requirementfor
newly synthesized ones. Cultures that were
shiftedup to390C at 3 h p.i.werereturned to 280C 1 hlaterandincubatedin thepresence or
absence of cycloheximide (100 ,ug/ml). Viral
RNAsynthesized inthese cultureswas labeled
during 60-min pulse periods with [3H]uridine,
and the amountsoflabeled viral RNA andRF
RNA andtheproportion of labeled RF RNAin
minus strandsweredetermined.Areturn to the
permissivetemperature inthe absenceof
cyclo-heximide ledtothe resumption ofanincreasing
rateof RNAsynthesis that eventuallyreached
about 60%of that in infected cultures thatwere
maintainedat280C (Fig. 6). However, areturn
to280Cinthepresenceofcycloheximide ledto
only a small increase over the rate of RNA synthesis occurringat390C.After thereturn to
280C,
the amount oflabeled RF RNA in the untreated culturesincreased, but essentiallynoincrease withtime inthe amountof labeled RF RNA occurredinculturestreated with cyclohex-imide(Fig. 6B).Correspondingto anincreasein
synthesis of totalRNAand RF RNA afterthe
return to280C,therewas aresumption of
minus-strandsynthesis(Fig.6C).Asimilarresumption of minus-strandtranscription aftertheshift back down to280C didnot occur inthe presence of cycloheximide.Active minus-strandpolymerase
musthave been present in all ofthesecultures
at the time of the shift to the nonpermissive
temperature, since minus strands were
accumu-lating at 3 h p.i. at
280C.
Since minus-strandpolymerase activity in SIN HR-infected cells
wasnormally short-lived,as wasdemonstrated
byitsrapid disappearanceafter the inhibition of
T
x=
CE
-C
II
x E
Vb
a
E
z
CK
Hours. post-infection
FIG. 6. Effect of cycloheximide addition on the
temperature-dependent resumption ofminus-strand
synthesis. tsll-infectedcultureswereshiftedto39°C
at3hp.i.,and RNAsynthesiswasmonitoredby the
addition of[3H]uridinefor 30-minpulses at 39°C (x). Two setsof these cultureswerereturnedto28°C at 4hp.i. Oneset wasincubatedcontinuouslyin the presence of100pgofcycloheximideperml (A); the secondset was notfurthertreated and servedasthe 28°C control(0).Incorporationof radioactivityinto theacid-insolubleform bythese cultureswas deter-minedafterthe culturesweregiven60-minpulsesof
[3H]uridinebeginningfromthe timeofthereturnto
28°C.Inaddition,severalcultureswhichwere main-tainedat28°C throughouttheexperiment (0) were
pulsed for60-minperiodswith[3Hluridinebetween
2and3hp.i.andbetween 8and9hp.i. tomonitor theamountofRNAsynthesis occurring in infected
culturesnotsubjectedtothesetemperatureshifts. (A) Totalincorporation into viralRNA; (B) incorpora-tion into RFRNA;(C)percentageoftotal labeled RF RNAinminus strands.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.503.284.435.70.448.2]356
proteinsynthesis (Fig. 1), the failure to
demon-strate aresumption of minus-strand RNA
syn-thesis in SIN HR tsll-infected cells that had
been shifted to390C at 3 h p.i. and then returned
to 280C 1 h later in the presence of
cyclohexi-mide couldhave resulted fromeither therapid
degradation or therapidfunctional inactivation
at390C ofaviral polypeptide required for mi-nus-strand RNA synthesis. In either case, the
minus-strand polymerase activity that
reap-pearedin SIN HRtsll-infected cellsupon the
return to28°Crequiredthesynthesis of proteins.
Cultures shifted to 39°C at 3 hp.i. were
re-turned to28°C atvarious times (Fig. 7). In all
cases, there was an immediate increase in the
[3H]uridineincorporationascompared with that detected in infected cultures not returned to
280C.Increasing thetime atthenonpermissive
temperaturebefore thereturntothepermissive
temperaturedecreased theextent towhichtotal
RNAsynthesisrecovered, but, nevertheless, mi-nus-strand synthesis resumed in tsll-infected cultures if the cultures were shifted back to
280C.
DISCUSSION
Previously,we have shown that minus-strand
transcription in Semliki Forest virus-infected
cellsisshut off selectivelywhen protein
synthe-sis isinhibited with puromycinor cycloheximide
(25).The data presented in this study extended
this observation to include SIN and demon-strated that when SIN HR tsll-infected cells
10
x 8
caE
6
C=4
4I2
2 3 4 5 6 7 8 9 10 11
Hours, post-i nfection
FIG. 7. Temperature-dependent increase in tsll RNA synthesis. CEF monolayers infected with tsll
wereshiftedto39°Cat3hp.i. At the indicatedtimes, certainof these cultureswerereturnedto28°Cand labeled with[3H]uridineat28°C for 60-minperiods; culturesmaintainedat39°Cwerepulsed for30-min
periods. Cultureswereharvestedinto ET buffer with
2% sodium dodecyl sulfate immediately after the pulse, and theamountof incorporation into the acid-insolubleform was determined. Symbols: 0, 39°C; 0,280C.
wereshifted to the nonpermissivetemperature
during the early phase of the viralreplication
cycle, when therateof minus-strand RNA
syn-thesis was increasing, minus-strand transcrip-tion ceasedselectively. Temperature sensitivity
of a component of the viral RNA-dependent
RNA polymerase that is required for minus-strand transcription would account for the
RNA-phenotype of SINmutant tsll.The
prod-ucts ofatleast four cistrons (complementation
groups A, B, F,andG) of SIN HR arerequired
forviral RNA synthesis (5, 6, 22, 29). SINHR tsllistheonlymember of theB
complementa-tiongroup(22, 28,29). The results ofourstudies
on ts mutantsofSINHR suggestthat the
prod-uctof theBcistronof SIN HR isapolypeptide
thatfunctionsinminus-strandtranscriptionand demonstrated that thermal inactivation of
mi-nus-strand transcription occurred in tsll-in-fected cellswithoutinactivation ofplus-strand
transcription.
Minus-strand RNA synthesis was also ob-servedtobe temperature sensitive in SIN HR
ts4 (complementation group A)-infected cells
and SIN HR ts6 (complementation group
F)-infected cells. This observation was consistent withthe results reported by Pfefferkorn et al. (21) andPfefferkorn andBurge (20), who dem-onstrated that inputts4andts6viral RNAsare notconverted intoanuclease-resistant, double-stranded form at the nonpermissive
tempera-ture. However,since we (16;unpublished data)
and others (22) have shown that ts6 has
diffi-cultysynthesizingplus-strandRNAatthe
non-permissive temperature, we conclude that the
product of the F cistron is required for both
plus-strand and minus-strand RNA syntheses. Cells infected withts4of theAcomplementation
groupsynthesize viralRNApoorly atboth the
permissive and nonpermissive temperatures,
suggesting atemperature-independent defectin
the viral RNApolymerase (16). Another mem-ber of the Acomplementation group, tsl5, did not stop synthesizing minus-strand RNA upon the shift to the nonpermissive temperature.
Most ts mutants of the A complementation
group show an alteration in the ratio of 26S-to-42S plus-strand synthesis after the shift to the
nonpermissivetemperature(29), with the
excep-tion ofts4,which does notsignificantlychange
the ratio of 26S-to-42S plus-strand synthesis
(16). Therefore, we conclude that only tsll of
the Bcomplementationgroupunequivocally
ap-pears to be defective selectively in minus-strand RNAsynthesis. Ashift of tsll-infectedcellsto the nonpermissive temperature at a late time
afterinfection does notaffectplus-strand RNA
synthesis (16). When tsll-infected cells were
shifted to the nonpermissive temperature at
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.503.77.220.413.550.2]early times in the infectious cycle, plus-strand RNA synthesis continued atthe rate that was
occurringatthetime of the shift.
Our results with SIN HRtsll suggest that the
product of theBcistron isa polypeptide which
is required selectively for minus-strand RNA
synthesis. Because minus-strand, but not
plus-strand, RNA synthesis is short-lived and
re-quires continualprotein synthesis, it is not
pos-sible fromourresultstodetermine whether the
productof the B cistron istemperature sensitive
in tsll orwhetherthe cleavageof the
polypep-tide encodedby theBcistronis defective at the
nonpermissive temperature. It has been
sug-gested (29) from the results reported by Waite
(30) that one ofthe polypeptide intermediates inthe processing of the SINnonstructural
pro-teins accumulates in tsll-infected cells at the
nonpermissivetemperature.The isolationof
ad-ditionalts mutantsbelongingtotheB
comple-mentationgroupwhichalsoshare the phenotype
of SIN HR tsll is necessary to conclusively demonstrate thatthe Bcistron is responsible for
minus-strand synthesis.
To date, only three SIN nonstructural
pro-teins have been identified (4, 14). Their gene
order has been determined to be 5'-ns6O-ns89-ns82-3' (4). These proteins are translatedfrom
the 42SpolycistronicmRNAwith a single
initi-ation site(1, 4, 15). A recent study byFullerand
Marcus (14) with increasing doses ofUV
irradia-tion, which resultsinthe blockingof translation
of polypeptides beyond the site of damage on
the mRNA, confirmed the gene order of the
nonstructural proteins of SINand also assigned twoof theseproteinstoparticular complemen-tationgroups;theN'-ns6O proteinwasidentified
astheproduct of the G complementationgroup,
and thens89proteinwasidentifiedas the prod-uctoftheAcomplementationgroup. The third
protein (ns82) could not be assigned to either
the B or F groups because of the verysimilar dose of UV irradiationresulting in loss of
com-plementation withmutantsof boththe F(tsllO)
and theB(tsll)groups.The authorsconsidered
thepossibility that the fourth complementation
group represents anoncoding region of the
ge-nomeRNA and thusfunctionsatthe nucleotide level rather thanatthepolypeptide level.Ifthe
Bcistronrepresentssuchanoncoding regionof
the genome, its ability to function in
minus-strandtranscription wouldnotbeexpectedtobe
sensitive to protein synthesis inhibition. After
the return to
280C
ofSINtsll-infectedcellsthatwere shifted to
390C
earlyininfection,thesyn-thesis of minus-strand RNA recovered only in
the absence ofcycloheximide.Wewould suggest
that, if there areonly three nonstructural
pro-teins,the ns82proteincontainstwofunctionally
distinctdomains,oneof which functionsin
mi-nus-strand RNA synthesis and thus can be
in-dependently mutated, resulting in theB andF
complementationgroups.
Thataspecific function notneededforviral
plus-strand transcription is involved in
minus-strandsynthesis isnotsurprising. The initiation of minus-strand synthesis may be unusual in
that the5'-initiating nucleotidemay be a
pyrim-idine: 5'-ppUp and 5'-pUp residues have been detected in a polyuridylate sequence which is
locatedatthe5' terminus of Semliki Forest virus
minus-strand RNA (23). A polyuridylate
se-quencehas beenfound alsoinSIN HR
minus-strand RNA (12).Althoughasimilar 5'-terminal polyuridylate sequence is presentin poliovirus minus-strand RNA(27, 33, 34), its 5' terminus is covalently coupled to a viral protein (10, 18). Since alphavirus plus-strand RNA synthesis probably initiates with apurine residue (9, 19,
32),anadditional functionmayberequired for pyrimidine initiation. Also, the ability to
regu-late theamountof minus-strandRNA, which is only needed inatemplate capacity and is stable
once synthesized (3, 24-26, 31), confers the
added advantage of the efficient utilization of
available substrates for single-stranded virion
RNA and mRNAformationoncesufficient
mi-nus-strand templates areformed. Since regula-tion ofthe initiationofminus-strand transcrip-tionwould controldirectly the number of
tem-platesproduced, translational control of the
pro-teinrequired for this initiation would bea
sen-sitiveregulatorymechanism.Theactualprocess
of initiation mayrequire the function ofmore
than asingle protein. Therole ofoneviral and
several host cell proteins in QB and f2 RNA phage transcription and replication has been
recently reviewed (1), and in vitro evidence
in-dicates that several picornaviruses, e.g.,
polio-virus (7) and encephalomyocarditis virus (8), mayutilizeahostcellproteininadditiontoviral proteins fortranscriptionon aplus-strand
tem-plate,possiblyfor the initiation oftranscription
(7). Whether this is true for alphaviruses also remainstobedetermined.
ACKNOWLEDGMENTS
We thankRaija Lahdensivu and Ritva Rajala for excellent technical assistance. WeareendebtedtoH.vonBornsdorffor hisgenerousgiftofpurifiedSIN.
Thisinvestigationwassupported byfundsfrom theSigrid Juselius Foundation andfrom the Finnish Academy,from PublicHealthServiceresearchgrantAI-15123from the Na-tionalInstitute ofAllergyandInfectiousDiseases,and from Biomedical Research Support grant 5S07-RR-05700-09 awardedbytheNationalInstitutes of Health.
LITERATURE CITED
1. Blumenthal, T., and G. G. Carmichael. 1979. RNA replication: function and structure of QB-replicase.
Annu.Rev. Biochem. 48:525-548.
on November 10, 2019 by guest
http://jvi.asm.org/
358 ET AL.
2. Bracha, M., A. Leone, and M. J. Schlesinger. 1976. Formation of a Sindbis virus nonstructural protein and its relation to42S mRNA function. J. Virol. 20:612-620. 3. Bruton, C. J., and S. I. T. Kennedy. 1975. Semliki Forest virusintracellular RNA: properties of the mul-tistranded RNA species and kinetics ofpositive and negativestrand synthesis. J.Gen.Virol. 32:413-430. 4. Brzeski, H.,and S. I. T.Kennedy. 1977.Synthesisof
Sindbis virus nonstructuralpolypeptidesinchicken em-bryofibroblasts. J. Virol.22:420-429.
5. Burge, B.W., and E. R. Pfefferkorn. 1966.Isolation and characterizationofconditional-lethal mutants of Sindbis virus. Virology 30:204-213.
6. Burge, B. W., and E. R.Pfefferkorn. 1966. Comple-mentation between temperature-sensitive mutants of Sindbis virus.Virology 30:214-223.
7. Dasqupta,A.,P.Zabel,and D.Baltimore. 1980. De-pendenceof theactivity of thepoliovirus replicaseon ahostcell protein. Cell 19:423-429.
8. Dmitrevia, T. M., M. V.Schleglova, and V. I. Agol. 1979.Inhibition ofactivity ofencephalomyocarditis vi-rus-induced RNA polymerase by antibodies against cel-lular components.Virology 92:271-277.
9. Dubin, D. T., K. Timko, S.Gilies,andV.Stollar. 1979. The extreme5'-terminal sequences of Sindbis virus 26S and 42S RNA.Virology98:131-141.
10.Flanigan, J. B., R. F. Pettersson,V.Ambros, M. U. Hewlett,and D. Baltimore.1977.Covalentlinkageof aprotein to a defined nucleotide sequence at the 5'-terminus of virion and replicativeintermediate RNAs ofpoliovirus. Proc. Natl. Acad. Sci. U.S.A. 74:961-965. 11.Franklin, R. 1966. Purification and properties of the replicative intermediate of the RNA bacteriophage R17. Proc. Natl. Acad. Sci. U.S.A. 55:1504-1513.
12.Frey, T. K., and J. H. Strauss. 1978. Replication of Sindbis virus. VI.Poly(A) and poly(U) in virus-specific RNAspecies.Virology 86:494-506.
13.Friedman,R. M., and P. M.Grimley. 1969. Inhibition of arbovirus assembly bycycloheximide. J. Virol. 4: 292-299.
14.Fuller, F. J.,and P.I. Marcus. 1980. Sindbis virus. I. Gene order of translation in vivo.Virology107:441451. 15.Kaariainen,L, and H. Soderlund. 1978. Structure and replication of alphaviruses. Curr. Top. Microbiol. Im-munol. 82:15-69.
16.Keranen, S., and L.Kaariainen.1979.Functional de-fects ofRNA-negative temperature-sensitive mutants ofSindbis andSemliki Forest viruses. J. Virol. 32:19-29.
17. Lowry,0.H., N. J. Rosebrough, A. L. Farr, and R. J. Randall. 1951. Protein measurementwith the Folin phenolreagent.J. Biol. Chem. 193:265-275.
18. Nomoto, A., B. Detjen, R. Pozzatti, and E. Wimmer. 1977.The isolation of thepolio genome protein in viral RNAs anditsimplication for RNA synthesis. Nature (London) 268:208-213.
19. Pettersson,R.F., H. Soderlund, andL.Kaariainen. 1980.Nucleotide sequence of the 5'-terminal Tl-oligo-nucleotideof Semliki Forest virus 42S and 26S RNAs isdifferent. Eur. J. Biochem.105:435-443.
20. Pfefferkorn, E.R., and B. W.Burge. 1968. Morpho-genic defects in thegrowth oftsmutantsof Sindbis virus.Perspect. Virol. 6:1-14.
21. Pfefferkorn, E.R., B. W.Burge, and H.M. Coady. 1967.Intracellularconversion of the RNA of Sindbis virus into adouble-stranded form. Virology 33:239-249. 22. Pfefferkorn, E. R., andD.Shapiro.1974.Reproduction oftogaviruses, p. 171-230.In H.Fraenkel-Conrat and R. R.Wagner. (ed.), Comprehensive virology, vol.2. Plenum Publishing Corp., New York.
23. Sawicki,D.L.,and P. J. Gomatos.1976.Replicationof Semliki Forest virus:polyadenylateinplus-strand RNA andpolyuridylate inminus-strand RNA. J. Virol 20: 446-464.
24. Sawicki,D.L.,L.Kaariainen, C., Lambek,and P. J. Gomatos. 1978. Mechanism for control of synthesis of Semliki Forest virus 26S and 42S RNA. J. Virol. 25:19-27.
25. Sawicki,D.L., and S.G. Sawicki. 1980. Short-lived minus-strand polymerase forSemlikiForest virus. J. Virol. 34:108-118.
26. Scheele,C.M.,and E. R. Pfefferkorn.1969.Inhibition ofinterjacent ribonucleic acid (26S) synthesis in cells infectedby Sindbis virus. J. Virol. 4:117-122. 27. Spector,D.H.,and D. Baltimore. 1975.Polyadenylic
acid on poliovirus RNA. IV. Poly(U) in replicative intermediate anddouble-stranded RNA.Virology67: 498-505.
28. Strauss,E.G.,E. M.Lenches,and J. H.Strauss.1976. Mutants ofSindbis virus. I. Isolation andpartial char-acterization of89 newtemperature-sensitivemutants. Virology 71:154-168.
29. Strauss, J. H., andE. G. Strauss. 1980. Mutants of alphaviruses:geneticsandphysiology,p.393-425. InR. W. Schlesinger (ed.), Togaviruses, biology, structure, replication. AcademicPress,Inc.,New York. 30. Waite,M. R.F. 1973.Proteinsynthesisdirected byan
RNA-temperature-sensitive mutantof Sindbis virus. J. Virol. 11:198-206.
31. Wengler, G., andG. Wengler. 1975. Studies on the synthesis of viralRNApolymerase-templatecomplexes inBHK-21 cells infected with Semliki Forest virus. Virology 66:322-326.
32. Wengler,G.,G.Wengler,and H. J.Gross.1979. Rep-licativeformofSemliki Forest virus RNA containsan unpaired guanosine.Nature(London)282:754-756. 33. Yogo, Y., M. Y.Teng,and E. Wimmer.1974.Poly(U)
in poliovirus minus-strand RNA is5'-terminal. Bio-chem.Biophys.Res.Commun.61:1101-1109. 34. Yogo,Y., and E. Wimmer.1973.Poly(A) and poly(U)in
poliovirus double-stranded RNA. Nature (London) NewBiol.242:171-174.
on November 10, 2019 by guest
http://jvi.asm.org/

![FIG. (B) 3 h p.i., 39°C, and pulsed withuridine0.01wereinfectedsizednonpermissiveturesandtainedintoculturescells 8 h andl -infected M ET the [3H]uridine were collected, were at these of CEF at Tris-hydrochloride dodecyl between maintained 93,000 extracts b](https://thumb-us.123doks.com/thumbv2/123dok_us/1475261.100155/6.503.262.460.58.224/withuridine-wereinfectedsizednonpermissiveturesandtainedintoculturescells-infected-uridine-collected-hydrochloride-maintained-extracts.webp)
![FIG. 5.pulsed[3HJuridinemonolayersshownreannealminus-strandmaintainedviralthe Rate of [3H]uridine incorporation into SIN tsll RNA at 28°C and after the shift to 39°C](https://thumb-us.123doks.com/thumbv2/123dok_us/1475261.100155/7.503.152.372.70.559/fig-pulsed-hjuridinemonolayersshownreannealminus-strandmaintainedviralthe-rate-uridine-incorporation-shift.webp)
![FIG. 6.presencepulsed28°C28°C.[3H]uridine2synthesis.secondadditiontemperature-dependentatminedatthetained(x).RNAtheculturestion intoTotal and 4 3 Effect of cycloheximide addition on the resumption of minus-strand tsll-infected cultures were shifted to 39°C](https://thumb-us.123doks.com/thumbv2/123dok_us/1475261.100155/8.503.284.435.70.448/presencepulsed-synthesis-secondadditiontemperature-dependentatminedatthetained-rnatheculturestion-intototal-cycloheximide-resumption.webp)