JOURNAL OFVIROLOGY, Oct.1980,p.189-203
0022-538X/80/10-0189/15$02.00/0 Vol. 36, No. 1
Fine-Structure
Mapping and Functional Analysis of
Temperature-Sensitive
Mutants in the
Gene Encoding the
Herpes
Simplex
Virus Type
1
Immediate Early Protein VP175
RICHARD A.F. DIXON'2AND PRISCILLAA.SCHAFFER'*
TheSidney Farber CancerInstitute,HarvardMedical School, Boston, Massachusetts02115,1and
Department
of
Virology,
Baylor
CoUege of
Medicine,
Houston,
Texas770302Herpes simplex virus (HSV)-specific proteins fall into at least three kinetic
classes whose synthesis issequentially and coordinately regulated.
Temperature-sensitive (ts) mutants of one complementation group (1-2) are defective in the
transition from immediate early to early and late protein synthesis. To elucidate thefunctionof the 1-2 gene product in the HSV type 1 replicative cycle, nine ts
mutants in this group were mapped by fine-structureanalysis and characterized
biochemically. Physicalmapping by homotypic marker rescue has shown thatall
members of the group lie within thetenninally repeatedsequencesof the S region
of the genome. Fine-structure geneticandphysical mapping permitted the
mu-tations to be ordered within these sequences.Because it has been shown that the
messagefor VP175 and the DNAtemplate
specifying
thisprotein extend beyondthe limits of thephysical map of the mutations, it follows that the mutations
mustlie within thestructuralgenefor VP175.Sodiumdodecyl
sulfate-polyacryl-amide gel electrophoresis analysis showed that most members of the group
overproduced the immediate early proteins VP175, -136, -110, and -63 and
markedlyunderproduced earlyandlateproteinsatthenonpermissive
tempera-ture. Intemperatureshiftup experiments,itwasfound that thesynthesisofearly
andlateproteinsceased, whereas the synthesis of immediateearlyproteins began
again. Thus, it is postulated that VP175 is (i) involved in the transition from
immediateearlytoearly protein synthesis, (ii) requiredcontinuouslytomaintain
early proteinsynthesis,and (iii)autoregulated, actingtoinhibit immediateearly proteinsynthesis.
The genome ofherpes
simplex
virus type 1(HSV-1) is large
(molecular weight,
96 x106)
andstructurally complex (1,
10, 11, 18, 38,44).
Despiterapidprogress in the
genetic analysis
ofHSV, little is known about theregulationofviral
geneexpression. The observation that the
syn-thesis of three classes of viral polypeptides is
sequentiallyand
coordinately regulated
wasfol-lowedbythedemonisrationthat coordinate
reg-ulation isatleast
partly
controlledatthelevel oftranscription (7, 8, 13, 14, 17, 22, 24,25,32, 43,
47). More
recently,.a
viral gene has beeniden-tifiedwhichactsvery
early
in the HSVreplica-tivecycle andcontrols the transition from im-mediate
early
toearly
and lateprotein synthesis
(8, 24, 25,32).Thisgeneislocated in theterminal
repeats of S (20, 29, 33,
42)
and isthought
toencodeaviral
polypeptide
of molecularweight
175,000,
designated
ICP4(13)
orVP175(8).
Anumber of
temperature-sensitive (ts)
mutantsinthisgene havebeenisolated(5, 28,
36).
Studieswith these mutants have revealed that VP175
controls a
transcriptional event(s)
which ises-sential forthetransition fromimmediate
early
functionstoearlyandlatefunctions(28, 32, 47).
Additionally, the gene is diploid (20, 27, 33),
presentinganunique situation for genetic
anal-ysis.
It is currently believed that VP175 is the
product of gene 1-2; however, most of the
evi-dencetosupport this contention is
circumstan-tial. Thus,mutantsdefective in this gene
over-produce VP175, the mutant protein exhibits
al-tered intracellular compartmentalization and
migrationinsodium dodecyl sulfate(SDS)-gels,
and thetemplate
specifying
VP175 maps in thesame region of the genome as do mutants in
group 1-2(27,29, 41,42).Todetermine whether
the lesions in mutants in group 1-2 are in the coding sequence for VP175, we have mapped nine ts mutants in group 1-2 by physical and
geneticmethodsandcorrelated their order and
map locations with the known location of DNA
sequences whichspecify VP175 (27, 33) and the
mRNA which encodes thisprotein (48).
Other investigators have demonstrated that
onefunctionoftheproductofgene 1-2 involves
transcriptional regulation (32, 47). To further 189
on November 10, 2019 by guest
http://jvi.asm.org/
190 DIXON AND SCHAFFER
define therole of thisproteininregulation,we
have examined viral polypeptide synthesis by one mutantin the group,tsB21u,intemperature
shiftup experiments. Our results demonstrate
thatVP175 is involved inmediatingthe
transi-tion fromimmediateearlytoearly protein
syn-thesis, that it is required continuously for the
synthesis of some early and late proteins, and
that itacts toinhibit itsownsynthesis.
MATERLALS AND MErHODS Cellsandcellculture.Serially propagatedhuman embryoniclungfibroblasts(HEL), primaryrabbit kid-neycells(RK),andAfrican greenmonkeykidneycells (Vero) wereused. Cellsweregrown inEagleminimal essential medium(Autopow;FlowLaboratories,
Rock-vile,Md.) containing10% fetal bovineserum, 0.03% glutamine, and 0.025% NaHCO3. Cells were main-tained in thesamemediumcontaining5%fetalbovine serum.Virusstocksweregrown,andcomplementation
andrecombinationtests wereperformedin HELcells;
RKcellswereused for the isolation of viral DNA and for markerrescuestudies.Virusinfectivityassayswere performedin Vero cells.
Viruses and virus assays. The KOS strain of HSV-1 was used as the wild-type virus (40). The isolation ofmutants tsB2b, tsB21u, tsB27h, tsB28h,
tsB32h,tsJ12g,ts90l,tsLB2, tsD,and tsc75 has been
reportedpreviously (4,5, 12,15,20,34).The last four
mutants wereobtained from M.Levine,I.Halliburton,
J. Subak-Sharpe, and R. Honess,respectively. Virus stocks were prepared and infectivities were deter-mined as previously described (36). In all tests, the
permissivetemperaturewas34°Cand the
nonpermis-sivetemperaturewas390C.
Complementation tests and mapping proce-dures. Mutants tsB2b, ts9Ol, tMD, and tsLB2 were showninapreviousstudytodefinecomplementation
group 1-2 (36). Complementation analysis with the othermutantsused inthisstudyis describedherein.
Complementationtestsandrecombination tests using two-factorcrosses werecarriedout asdescribed
pre-viously(34,37).Markerrescueexperimentswere per-formedasreportedbyChuetal.(5). The restriction maps of strainKOS used todetermine the physical locationsof mutations in markerrescuestudies were taken from Skare and Summers (EcoRI) (39) and Wagner andSummers (HincII) (45) or weregenerated
in thislaboratory(BamHI).
Isolation ofviralDNA, digestion with
restric-tionenzymes, and agarose gel electrophoresis.
DNAwasisolated frompartially purifiedcytoplasmic andextracellular virionsasdescribedby Chu et al. (5). Restrictionenzymes were purchased from New Eng-land Biolabs (Boston, Mass.), and DNA was digested with2U ofenzyme per
jig
ofDNA. Fragments were separated byelectrophoresis on horizontal slab gels andisolatedaspreviously described (5).Analysisofinfected cellpolypeptides.HEL cell
monolayerswereinfectedat amultiplicity of 100PFU/
cell andincubated in water bathsat either 34 or
390C
under the conditions describedin the text. Polypep-tideswerelabeled with 5pCiof
'4C-amino
acids perml (Schwarz/Mann, Orangeburg, N.Y.; amino acid
mixture, 50 to 60 mCi/matom) in maintenance
me-dium containing 10% of the normal amounts of all amino acids except arginine and glutamine, which wereused in normal amounts. Immediateearly pro-teins (31) were induced in infected cultures treated with 50 ug ofcycloheximide (Calbiochem, LaJolla,
Calif.) per ml from 0 to 7 hpostinfection. Thedrug wasremoved after7h,monolayerswerewashed five times withTris-saline, and mediumcontaining5 uCi of'4C-amino acids and 15pgofactinomycinD (Cal-biochem)permlwasadded.Cellswereharvestedat8 h postinfection by scraping into the medium. They
werewashed threetimes in Tris-buffered slineand suspended in water, and the resulting suspensions
werefrozen.Sampleswerelaterthawed,sonicatedfor 2min,adjustedto0.05MTris-hydrochloride (pH
6.7)-1%SDS-1%2-mercaptoethanol-0.5M urea, andboiled
for 2 min.Electrophoresiswascarriedout on7to15%
lineargradientSDS-polyacrylamideslabgels (23).The
gelswere fixed in7% acetic acid and 50%methanol, impregnated with Enhance (New England Nuclear
Corp.,Boston,Mass.),and dried. The driedgelswere
thenfluorographedat-70°ConKodakSB-5film.
RESULTS
Complementation studies with ts
mu-tants. Intwoseparatetests, eightof theninets
mutantsusedin thisstudywereshowntobelong
tocomplementation group 1-2 (5, 36).To
con-firn the assignment of these mutants and of
tsc75 (20) togroup 1-2,aquantitative
yield-of-progeny complementation test was performed
(Table 1); tsJl2g, a memberofgroup 1-9, was
used as the positive control (36). None of the
ninemutantstested complementedeach other,
whereas all complemented tsJ12g (Table 1).
Thus, all ninemutantsbelongedtogroup 1-2.It
should be noted that theninemutantsincluded
in this study behave as if they possess point
mutations inasinglegene;i.e.,theyrevertwith
lowtomoderatefrequency(datanotshown) and
exhibitnonoverlappingpatterns of
complemen-tation intestswith members ofothergroups (5,
36).
Physical mapping experiments. Marker
rescuestudieswereundertakentodeterminethe
physical map locations of the nine ts mutants.
We have reported previously that tsB21u lies
withinthe "c"sequencesin theterminal repeats
of the Sregionof thegenome(29).Thislocation
is in agreement with thefindings of Stow and
Wilkie(42) andKnipeetal. (20,21), whoplaced
tsLB2,tsD, andtsc75withinEcoRIfragment K,
which contains S-terminal sequences "c" and
Sta."Ps
Table 2 shows the results of marker rescue
experiments using purified EcoRI fragments
shown inFig. 1.tsLB2,tsD,and tsc75hadbeen
shown previously to bein fragment K (20, 21,
42). Efficientrescuewith the remaining six
mu-tants was observed with fragment K (0.966 to
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
HSV-1 ts MUTATIONS IN VP175 191
TABLE 1. Complementation between members ofgroup 1-2
Virus B2 B21 B27 B28 B32 901 LB2 tsD tsc75 tsJl2g
tsB2b 1.5a 0.70 0.81 0.37 0.40 0.55 0.48 0.70 170
tsB21u 0.31 0.27 0.86 0.76 0.88 0.33 0.32 4.9
tsB27h 0.28 0.18 1.9 1.5 0.61 0.68 19
tsB28h 0.20 0.42 0.68 0.13 0.50 72
tsB32h 1.4 0.84 0.28 0.38 580
ts901 0.45 0.10 0.20 570
tsLB2 0.44 0.91 22
tsD 0.35 444
tsc75 30
aComplementation index=[yield (A +
B)]/[yield
(A)+(B)]; infected cultures wereincubated at 39°C, and yields were assayed at 34°C; values greater than 2.0 (in italics) were consideredpositive(34).TABLE 2. Marker rescuetests with mutants in group 1-2: rescue with EcoRI fragments
Fragment' Virus
A,B,C D,E,F,G H I,J K None tsB2b 4.0 0.0 0.0 0.0 0.7 0.0
tsB21u 10.0 0.0 0.0 0.0 9.6 0.0
tsB27h 1.5 0.0 0.0 0.0 1.0 0.0 tsB28h 0.7 0.0 0.0 0.0 0.6 0.0 tsB32h 7.0 0.0 0.0 0.0 41.0 0.0 ts901 22.0 0.0 0.0 0.2 1.0 0.0
aValues represent the ratio of the yield of progeny
virusfrom transfectedcells when assayed at 39°C as compared with the yieldat340C(xlOO). Virus titers
at34°C were always greater than 105 PFU/ml.
1.000) and with amixture offragments A, B, and
C; fragments B and C are joint-spanning
frag-ments, both of which contain sequences in K.
TheEcoRI Kfragmentlies
entirely
withintheterminallyrepeated sequences of the Sregion.
Althoughwehavenotdetermined whether each
mutation is present within both repeats, it is
likely that this is thecase, sincefragments
rep-resenting both ends of S have been shown to
rescuetsB21u and thepresenceof the lesion in
only one end would permit self-rescue of the
mutant(29). Moreover,like
tsB21u,
theterminalfragments of the L region (J and E) failed to
rescue,indicatingthatnoneof the mutationslies
within"a"sequencespresentatthe
joint
andatthe terminiof themolecule.
Rather,
themuta-tions lie within the "c"sequences
adjacent
toUs
(20, 29). In the case of ts901, the low level of
rescueobservedwithcombined
fragments
IandJ wasprobably duetocontamination of theI,J
mixturewithfragmentK,becausealthough
frag-mentsJ andKsharesome commonsequences, rescuewas notobserved withfragmentE.
To further refine the map positions of the
mutants, EcoRI fragment K was isolated and
redigested with BamHI and
HincII;
each of these enzymesmakesasingle
cutwithin theKfragment,resultingin threefragments
(2.0,
1.85,and 1.25 kilobases
[kb])
which areeasily
re-solved(Fig. 1).When these fragmentswereused
in marker rescue tests, the results shown in
Table 3 were obtained. With tsB2b, tsB21u,
tsB27h, tsB28h, and tsB32h, rescue occurred
maxrimally
with the tenninal HincII fragmentK2 (0.988 to 1.000), whereas with ts901,rescue
occurred only with the middle fragment, K3
(0.979 to 0.988). Fine-structure mapping of
tsLB2 and tsc75 demonstrated that with the
formermutant rescue occurredonlywith
frag-mentKl,whereas with thelattermutantrescue
occurred with both K2 and K3.
Asummaryof thephysicalmap locations of
all nine mutants is presentedin Fig. 2. Inthis
figure, the limits of fragments which exhibited
rescue areshownasbars. Thewhiteportion of
each bar indicatessequencesinwhichmutations
are thoughtnot to lie bythe process of
elimi-nation;e.g.,failure of the termini of Ltorescue
tsB32h, tsB28h, tsB27h, tsB2b, and tsB21u
ex-cludestheirlyingin the "a"sequences,whereas
rescue oftsc75 by all four terminal fragments
excludesits lying in the "c"sequences. Because
direct tests of the sequences excluded by the
processof elimination havenotbeenconducted,
theoutermostlimitsof each barshould be
con-sidered definitive. The location of tsD (0.966to
0.900) was taken from Stow et al. (41, 42). It
should be mentioned that Wilkie and his
col-leagues haverecently refined the limits of tsD
and of tsK, another member of group 1-2, to
coordinates, 0.979 to 0.990 and 0.966 to 0.978,
respectively (personal communication).The
se-quence of mutations on the
physical
map istherefore tsK-tsLB2,tsD-ts901,
tsB32h-tsB28h-tsB27h-tsB2b-tsB21u,andtsc75.
Thelocation of thesequencespecifyingVP175 asdeternined byheterotypicrecombinationand marker rescue (27, 33) and the 4.7-kb mRNA
which codes for VP175 (48) are also shown in
Fig.2.Becausethemapcoordinatesof the mu-tantsliewithinthelimitssetforthe mRNAand
VP175 structural polypeptide and because no
evidence for
splicing
ofthegene forVP175 hasbeen reported, it is assumed that each ofthe
VOL. 36, 1980
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.510.51.242.233.324.2]192 DIXON AND SCHAFFER
FIG. 1. EcoRI restriction map of HSV-1, strain KOS. (1) L andS componentsofthe viral genome; (2) sequenceorganization; (3)fractionaldistance; (4)EcoRI restriction map in the P orientationofthe genome
(27)(fragments B andCarejoint-spanningfragments containingEplusK and JplusK, respectively); (5)
BamHI/HincIIrestriction mapofEcoRIfragmentK. TheterminiofK,aredeterminedbyEcoRI andBamHI
sites, theterminiofK3aredeterninedbyBamHI andHincII, and theterminiofK2aredeterminedbythe
HincIIsite and thephysicalendofthe Sregion. Thegel profileontheleftis apreparative gel of EcoRI-cleavedHSV-1 DNA fragments. The Kfragmentwasexcised and digestedwith BamHIandHincII. The
resultingfragmentsareshownin thegelprofileontheright.
mutants is defective inthe structural gene for
VP175.
Genetic mapping experiments. Because
markerrescue testsdidnotsucceed inseparating
all mutations physically within the repeat, a
linkage map was constructed by two-factor
crosses in an effortto order the mutantswith
respect to one another (Table 4, Fig. 2). The
following observations concerning the linkage
map are notable. First, the mutants clustered
into four groups, each separated by 1.4 to 1.9
recombination units. The firstgroup contained
mutantstsB2b, tsB21u,andtsB27h; the second
contained mutantstsB28h, tsB32h, and tsLB2;
the third contained tsD; and the fourth
con-tainedts9Ol.Second, although themutants
clus-tered into groups, no mutational hot-spotting
wasevident(i.e.,allmutationsinducedwithone
mutagen were not located in one place). Both
thymidine-specific (bromodeoxyuridine and
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.510.103.437.57.473.2]VOL. 36, 1980
UV) andcytosine-specific (hydroxylamine) mu-tagens wereusedformutantinduction, and
mu-tationsderivedwitheithertypeofmutagen were
randomlyarrangedonthemap.Third, although
the physicalmap is notsufficiently detailed to
establish the order ofmutantswithin eachof the
groups, thegenetic andphysicalmaps are
coli-nearwith respect tothe order ofseven of the nine mutants tested: tsD-ts901 and
tsB32h-tsB28h-tsB27h-tsB2b-tsB21u.Only tsLB2 isout
of orderonthegeneticmap.Fourth, the
recom-TABLE 3. Marker rescue tests with mutants in group 1-2: rescue withBamHI,HincII-cleaved
EcoRI-K Fragmenta Virus
Ki K2 K3 None
tsB2b 0.0 2.1 0.0 0.0
tsB21u 0.0 4.5 0.3 0.0
tsB27h 0.0 0.3 0.0 0.0
tsB28h 0.0 0.1 0.0 0.0
tsB32h 0.0 1.4 0.0 0.0
ts901 0.0 0.0 5.4 0.0
tsLB2 7.3 0.0 0.0 0.0
tsc75 0.0 14.0 7.6 0.0
aSee Table 2.
c
0950 0.960 0.9,66 0.970
HSV-1 ts MUTATIONS IN VP175 193
binationfrequencies observed were largerthan
one might expect given the total physical
dis-tanceinvolved (3.1kb). In previous studies, the greatestdistancebetween mutations inasingle
geneinuniquesequencesin L was 3.3
recombi-nation units (37) (unpublished data). The high
frequencyofrecombination between mutants of
group 1-2could reflect (i) auniquepropertyof the repeated "c" sequences in that they may constitute a region of very activerecombination,
(ii) the diploid nature ofthe gene, or (iii) the
large size of thestructural gene for VP175. In
TABLE 4. Recombination betweenmutantsingroup
1-2
Virus B2 B21 B27 B28 B32 901 LB2 D c75
tsB2b 0.36a 0.51 3.2 4.1 4.0 1.4 2.2 0.87
tsB21u 0.91 4.2 2.8 7.7 2.3 2.1 1.6
tsB27h 4.9 2.7 6.1 1.4 1.3 1.8
tsB28h 0.11 3.8 0.73 4.5 0.31
tsB32h 3.4 0.71 1.45 0.55
ts9Ol 2.0 1.9 0.89
tsLB2 3.5 0.40
tsD 0.0
tsc75
a D~omomaun irequency=! r /IA
-Recombmation
frequency
=[yi4
39°C]/yield (A+B)at34°C]x2 x ]
a
0.9790.980 0.9880.990
B32B28,B27,B2,B
901
D
eld (A + B) at
100. SEQUENCE ORGANIZATION
VP175CODING SEQUENCE
4.7kb mRNA
1.000 FRACTIONAL
DISTANCE 121
c75 PHYSICAL MAP LB2
GENETIC MAP
FIG. 2. Physicalandgeneticmapsofmutantsincomplementationgroup 1-2. TheVP175codingsequence wastakenfromPrestonetal. (33) andMorseetal. (27). Thelimitsofthe4.7-kb mRNA anddirectionof transcriptionaretakenfrom Watsonetal.(48)andClementsetal.(6). Thefractionaldistance barindicates thepositionsof cleavagesitesofrestrictionenzymesusedin thisstudy:0.966=EcoRI,0.979=BamHI,and
0.988=HincII. The maximum limitsofthesequencesinwhich themutationsmaparepresentedassolid bars
(indicatingsequencesin whichthemutation isthoughttolie)andopenbars(indicatingsequences inwhich themutationprobablydoesnotlieasdeducedbytheprocessof elimination; seetextfor explanation). The geneticmapwasconstructedfromthe datapresentedin Table3.
moor.=
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.510.49.448.196.584.2]194 DIXON AND SCHAFFER
contrast to most mutants, low recombination group1-2. Todetermine whether all members of
frequencies wereconsistentlyobservedbetween the group examined in this
study
exhibit thistsc75 and other mutants (20) (Table4). More- property,cellswereinfectedwith eachmutant,
over, tsc75 appearsto becloselylinkedto
tMD,
incubated at39°C,
andlabeled from5 to 24 hyet it is located in the "a"sequencesatthe end postinfection. All of the mutants exhibited a
of thephysical map. It may be thatmutanttsc75 similarbutnotidentical
polypeptide
phenotypecontains multiplemutations,thepossibleorigins at
390C
(Fig. 3).Thus,for allmutants,immedi-ofwhich will be discussed later. ateearlypolypeptides
VP175,
-68,and-63 werePolypeptide synthesis in ts mutant-in- overproduced. These
polypeptides
correspondfected cells. We have shown
previously
that toICP4,
-22, and-27,
respectively,
of HonesstsB2b andtsB21uoverproduce immediate
early
andRoizman(13).
Although
VP136(ICP6)
waspolypeptides VP175, -136, -110, and -63 (8, 9). overproducedbyallmutantsexcept tsB28h and
Similarfindingshave beenreportedby Marsden tsB32h, VP110
(ICPO)
was overproduced onlyet al. (26) and Knipe et al. (21) with
tsD, tsK, by
mutants not derived from strainKOS,
i.e.,tsT, andtsLB2, all members ofcomplementation
tsLB2,
tsc75, andtsD.Additionally,
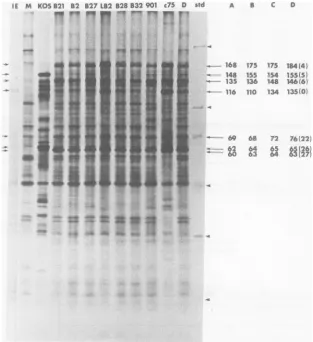
someearlyF M KOS82 B2 B27 LB2 B28 B32901 C75 D ;Std
A&MIL ~ | 168 175 175 184i4;
148 155 154
153,515
135 136 148 146(6- '16 P0 *34 '135O.
968 76762F
62 64 65 g 26;
60 63 64 27
_U
00:
o6
, *$ , ;l
FIG. 3. SDS-PAGE analysis of viral proteins synthesized in cells infected with ts mutants in complemen-tationgroup 1-2. HEL cells wereinfected with100PFU/cellat39°C, labeled with14C-amino acids from 5 to
24hpostinfection, andprocessed for PAGE as described in Materials and Methods.Abbreviations: M, mock-infected cells; IE, immediateearlypolypeptides; KOS, wild-type virus; std, molecular weight standards in
decreasingorder(220,000,93,000, 69,000, 44,000,30,000,and12,000). Themolecular weights of viral proteins presented are thosereported here and by otherinvestigators: A, this study; B, Marsden etal.(26)andPreston (31); C, Courtneyetal. (9);D,Honess andRoizman (13)andPereira etal. (30). Valuesindicatemolecular weightsx103.Numbers inparentheses are ICP designations ofHonessandRoizman (13).
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.510.113.426.236.578.2]HSV-1 ts MUTATIONS IN VP175 195
andlateviralpolypeptides(e.g.,VP64 and-155,
respectively) were synthesized by all mutants.
That the variations may reflect strain-specific
differences in gene expression cannot be
ex-cluded. However,analtemativeexplanation for
the observed variation concerns the fact that
somemutantsexhibitgreaterleakat39°C than
othersderived from thesamestrain. Thus, for
example, tsB28h and tsB32h synthesizedgreater
amounts oflate proteins at 390C than did the
otherKOSmutants (datanotshown). Figure3
alsoincludes the various molecular weights and
nomenclaturesused in the literaturetodescribe
thesepolypeptides. Inthisreport, wehave used
thenomenclature of Preston (31)(Fig.3,column
B).
Temperature shiftexperiments.To
deter-mine whetherfunctional VP175 isrequiredonly
transientlyor throughout the replicative cycle,
temperature shiftup experiments were
per-formed with tsB21u. In the first experiment,
tsB21u-infectedcellswereincubatedfor various
lengths of time at34°C before shiftupto
390C;
allcultureswerelabeledatthetimeof shift and
harvestedat24h (Fig.4).
When cultures were incubated continuously
at 390C (Fig. 4, lane b), overproduction of
im-mediate early polypeptides VP175, -136, -110,
and -63 wasevident. Late polypeptides VP154
and VP100 and early polypeptide VP64 were
minor species in these cells. In contrast, early
andlatepolypeptides predominatedincultures
incubated at340Cfrom 1,2,4, or 8through24
h(Fig.4,lanesc, e, g,andi).
Inculturesshiftedup at 1, 2,and4h (Fig.4,
lanesd,f, andh),immediateearly polypeptides
VP175, -136, and-63 werepredominant,
consid-erablymorebeingsynthesizedthan in the
340C
controls.Thus,immediateearly protein
synthe-siswascontinuous (i.e.,notshutoff) after
shif-tup. VP110 was also present, but to a lesser
extent than the other three polypeptides. In
cultures shiftedupat4h(lane h), VP175,-136,
-110, and -63 were present in greater amounts
than inthe340Ccontrols(lane g),but increased
amountsof VP154 and-64 were also observed. The synthesis ofVP110 is difficultto evaluate
intheseexperiments. Althoughlittlewasseenin
theshiftat2h andmore was seeninthe shifts
at 4and8h,thisincrease may be duetothefact
that anearlypolypeptide comigrateswithVP110
(30). Thus, theproteinseen atlater timesmay
bethe immediateearly polypeptide,VP110,the
earlypolypeptide,orboth.Clearly,however,the
synthesis of VP175, -136, and -VP63 was not
shut off after shiftup even though many early
andlateproteinswerebeingsynthesized
simul-taneously.Thus,despitethecurrenthypothesis
thatearly viralproteinsarerequiredtoturnoff
immediate early protein synthesis, these
pro-teins continuedtobe synthesized inthe presence
of early proteins. Because the only essential
lesion in tsB21u is in the gene for VP175, these
observationssuggestthe possibility that VP175
is involved in the inhibitionof immediate early
protein synthesisor,alternatively, thatVP175 is
required continuously to produce a factor
re-quiredto shut off the synthesis of this class of
polypeptides. Even whenthe shiftoccurred as
late as 8 h (Fig. 4, lane j), the expression of
VP175wasstillgreaterthan in the
340C
control(lane i).Additional differenceswerenotedwhen
proteins were synthesized after shift to
390C
(lane j) compared with unshifted cultures (lane
i;seedots between lanes i and j). Thus,it appears
thatfunctional VP175 is requiredevenafter8h
for theexpression of these proteins, butnotfor
theexpression of all early and late proteins.
Anadditional observation made fromthisgel
concernsvariationintheelectrophoretic
mobil-ity of VP175.Inallcases,the VP175polypeptide
made at
340C
migrated moreslowly than thatmadeat
390C
(mostclearlyseeninlanese,f,g,and hofFig. 4). This observation is inagreement
with the observations ofPreston, who showed
that incells infected with tsK, different forms of
VP175weresynthesizedatpermissiveand
non-permissive temperature (31), and of Pereira et
al. (30) and Wilcox etal. (49), who foundthree
forms ofwild-typeVP175 inpulse-chase
experi-ments.
Functional VP175 has been shown to be
re-quiredonlytransientlyfor viral DNAsynthesis,
therequirementbeing bypassed bya2-h
prein-cubationat
340C
(35). The results of thepresentstudy demonstrate that continued synthesis of
VP175 isrequiredfor the expressionof certain
late viralgeneproducts (see above).The
require-ment for functional VP175 in the synthesis of
infectious virus, however, has not been
exam-ined.Therefore, the yield of infectious virus in
cultures infectedasforshiftuptests(Fig. 4)was
measured(Table 5).
Inunshiftedcultures,tsB21uwastemperature
sensitive for growth when compared with the
wild-type virus, as expected. When
mutant-in-fected cultureswere shiftedup at1, 2, 4, and8
handyieldswereassayedafter24h, theywere
only10-foldhigherthan theyieldfrom the
cul-tureincubatedat
390C.
Thus,unlike viral DNAsynthesis, functional VP175 is apparently
re-quiredafter8hfor theproductionofinfectious
virus. Moreover,VP175appearedtoberequired
throughoutthe18-hgrowth cycle,since theyield
of infectious viruswasreduced 10-foldcompared
with unshifted cultures when cultures were
shifted up at 18 h (data not shown).
Alterna-tively, given the greatly diminished amount of
VOL. 36, 1980
on November 10, 2019 by guest
http://jvi.asm.org/
196 DIXON AND SCHAFFER
i< b d e f g h
D--p!
Am " ..
_ iF_g~~~~~~~~~~~~~~~~~~~~~~~~~-41
4_ 4m4_
4000 404M 4mm~~~~~~~~~~~~~~~~~~~~~~~~-t-~~~~~ VTl/:X; VP]5sa VP13C3p
VPIL
Vtp r
~~~~s 4
FIG. 4. SDS-PAGEanalysisofviralproteins synthe8izedin cellsinfectedwithtsB2lu aftertemtperature shiftup. (a) Mock-infectedHEL cells labeledfr-om4to24hat3900;(b) tsB2lu-infectedcellsincubatedat 3900 andlabeledfrom4to24h; (c-i)cellsinfected at3400 and held continuouslyatthistemperature or
shiftedupto3900 atdesignated times; (c,e,g,andi)cells incubatedat3400 andlabeledat1, 2, 4,and 8h,
respectively; (d,f, h, andj) cellsshiftedupandlabeledat1, 2, 4,and 8h,respectively; (k and1)wild-type
virus-infectedcells labeledfrom4to24hat34and3900,respectively.Allsamipleswereharvestedat24h and
prepared forPAGE and virusassay(TableS5)asdescribed in Materials and Methods.
VP175 synthesized after shiftup at 8 h (Fig. 4, lanej), it is possible that the same sequences whichspecifyVP175 alsospecify a later function
requiredforinfectivity,and thatamutation in
the sequences specifying VP175 could also in-activate thisputativelater function.
The experiment illustratedin Fig. 4
demon-strated that VP175 synthesis was notshut off J. VIROL.
...,£.,..
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.510.153.386.73.538.2]HSV-1 ts MUTATIONS IN VP175 197 TABLE 5. Yieldof tsB21u and wild-type virus after
temperature shiftup Initial
temp of Time of
Virus tiona shiftup (h) Virusyield
(OC)
tsB21u 34 -a 6 X107
39 - 8x 103
34 1 3x 104
34 2 3x 104
34 4 4x 104
34 8 8x 104
Wildtype 34 - 1 x l08
39 - 1 x 108
a
-,
Not shifted.whencultureswereshiftedup at 1 to 4h(lanes
fandb,respectively; notclear from thisgel,but
seeninotherexperiments),indicatingthat
max-imumsynthesis of VP136occurslater than other
immediate early polypeptides, an observation
thatled Pereiraetal.toclassifythispolypeptide
in the group of early or ,B polypeptides (30).
Whencells wereshifted up at4h andlabeled
from4 to8h(laned), immediate early (VP175,
-136, -110, and-63), early(e.g., VP64), and late
(e.g.,VP154)polypeptidesweresynthesized.
Be-causelittlesynthesis of VP175wasobserved at
340C
during this period (lane f), this findingsuggests that continued (or possibly renewed)
expression of immediateearlypolypeptides
oc-curredinconjunctionwith theexpressionoflate
polypeptides.Pulse-labelingforperiodsasshort
as 30 minproduced thesame results (datanot
shown). Shiftingup atlatertimes (i.e.,8 to 12
and 12 to 24h; lanesgandj) didnotresult in
theaccumulation of VP175orVP136; however,
the synthesis of VP63 (and, to alesser extent,
VP110)wasenhancedrelativetoitssynthesisin
34and
390C
controlcultures (laneshand i andlanes k and 1). Asin the case of infected cells
shiftedupfrom4 to8h,thesynthesis of
imme-diate early polypeptides (VP63 and -110)
oc-curredsimultaneouslywith thesynthesisoflate
(VP100 and-154) polypeptides.
Totestthepossibility that theaccumulation
of immediate early polypeptides in cultures
shifted to
390C
was duetotheirincreasedsta-bilityatthis temperature, mutant-infectedcells
werepulse-labeledat
390C
from4 to4.5h (Fig.5, lane m) and chased from4.5 to 8 hat
390C
(lane n)orafter shiftdownto
340C
(lane o).Thelabeled,chasedpolypeptideswereequallystable
atbothtemperatures(lanesnando),supporting
theconcept that theaccumulation ofimmediate
early proteins at
390C
aftershiftup wasduetocontinuedorincreasedsynthesisrather thanto
decreased turnover. Although it appears that
somesynthesis continued duringthe chase
pe-riod(Fig. 5), suchsynthesis was not observed in
other experiments. Therelative stability of the
protein at the two temperatures was identical,
however.
Additional evidence for the reexpression of
immediate early polypeptides after shiftup
comesfrom the observationthat when
tsB21u-infected cellswereshiftedat 4 or 8 h and either
pulse-labeled immediately for 1 h or
pulse-la-beled 2 h after the shiftup, the same level of
synthesis of VP175 was observed (data not
shown). Because the half-life of the early
mRNA's isonly1 h (46), it would benecessary
to synthesizenew mRNA in order to maintain
thesame rateofprotein synthesis.In this
exper-iment, expression of VP175 in cultures shifted
up at 8hwasobserved, whereasno expression
of VP175 was observed in cultures labeled at
340C during the same period, again indicating
that VP175 is reexpressed and therefore
in-volved ininhibiting itsownsynthesis.
Anadditional observation made from the gels
shown in Fig. 5 concerns the active form of
VP175.ItisapparentfromFig.4and5thatthe
form of VP175 whichaccumulatedat340Cwas
ofhigher apparent molecular weight than the
form which accumulated at 390C. After
pulse-labeling, afast-migratingform wassynthesized,
and this form could be chased to a
slower-mi-grating form which accumulatedat390C (Fig.5,
lanesmandn). After shift from39to340Cand
chase (lane o), aslight changeinapparent
mo-lecularweightwasevident. As mentionedabove,
the finding of three electrophoretic variants of
VP175is consistent with thereportsof Pereira
etal. (30), Preston (31), andWilcox etal. (49),
who have described three forms of theprotein
(VP175a, -b,and -corICP4a, -b,and-c) which
shareaprecursor-productrelationship. Preston
noted thatuponshift from34to
390C,
VP175cwasconvertedto VP175b, with loss ofactivity
(31).Consistent with thisobservation,wefound
that VP175b was converted to VP175c upon
shiftdown. Of additional interest is Preston's
findingthatupon shiftdown in the presence of
cycloheximide, no new protein synthesis was
required forearly transcription(32). From these
observations,itappearsthat the "c"forn is the
active form of VP175. Although the nature of
the modification in VP175 isnotknown,Wilcox
etal. (49)havepresentedevidence which
impli-catesphosphorylationinthemodification proc-ess.Whatever themechanismofmodification,it
isessentialforfunctionalactivity.
DISCUSSION
Theresultspresentedheresupport and
com-plement previousfindings concerningthe
loca-VOL. 36, 1980
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.510.48.242.80.237.2]198 DIXON AND SCHAFFER
a b c d e f g h i
I
k I m n o.VP1.4
VP136
.i~~~~~~~~~~~p-.0
* ~ _
.,A
_._Jss
1.
0*. 44
- -
VP4
-4
VPP53
-*m.
FIG. 5. SDS-PAGEanalysis ofviralproteinssynthesizedin cellsinfectedwith tsB21uaftertemperature
shiftup andpulse-labeling. HEL cells wereinfectedwith 100PFU/celland labeledasfollows: (a)
mock-infectedcells labeledfrom4to8hat34°C; (bandc)tsB21u-infectedcellslabeledfrom1 to4hat34and 39°C,respectively; (d)tsB21u-infectedcellsshifted from34to39°Cat4hand labeledfrom4 to8h;(eandf) tsB21u-infectedcellslabeledfrom4to8hat39and34°C,respectively; (g)tsB21u-infectedcellsshiftedup from34 to39°Cat8h and labeledfrom8to12h;(handi)tsB21u-infectedcellslabeledfrom8to12hat39
and34°C,respectively; (j) tsB21u-infectedcellsshiftedfrom34 to39°Cat12h andlabeledfrom 12to 24h;
(kand1)tsB21u-infectedcellslabeledfrom12 to24 hat39and34°C,respectively;(m-o)tsB21u-infectedcells labeledat39°C from4to4.5h andharvestedat4.5h(m)orchased with coldamino acid-containingmedia
from2.4 to 8hat39°C (n)or34°C (o).
tion and function of the gene encoding VP175.
Theoriginaldescriptionofcascaderegulation in
theHSV-1replicativecycle(13) ledtothe
iden-tification of a group of proteins (immediate
early,ora, proteins)whicharesynthesized
im-mediatelyafter infection and whosesynthesisis
required fortheinitiationofsynthesis of a
sec-ond group ofproteins, theearly, or,B,proteins.
Early proteinswereshowntoplay arolein the
shutoff of immediateearly protein synthesis(13,
14).
Amongthe immediateearly proteins first
de-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.510.117.422.72.496.2]HSV-1 ts MUTATIONS IN VP175 199
scribed were VP175, -110, and -63 (13). The
existence ofa mutant, tsB2b, which
overpro-duced VP175 andwasdefectivein thetransition
from immediate early tolate protein synthesis
was reported simultaneously (8). The latter
study also demonstratedthat in tsB2b-infected
cells, VP175wasnottransportedto thenucleus
asefficientlyasin wild-typevirus-infectedcells
at the nonpermissive temperature. Both the
transition fromimmediate early to late protein
synthesisandtransport tothe nucleuscould be
reverseduponshiftdown and bypassed by
prein-cubationatthepermissivetemperature,
indicat-ing that the defective function in this mutant
was bothearly and reversible. Subsequent
ex-periments demonstrated that functional VP175
was essential for viralDNA synthesis and that
therequirementcould be bypassedby
preincu-bation for 2 h at the permissive temperature
(35). More recently, it was shown that upon
shiftdown, new transcription was required for
viral DNAsynthesis in cells infected with tsB2b
(28).Otherinvestigators have isolated additional
tsmutantsofcomplementationgroup 1-2(D, T,
K, and LB2) (36) whichpossesssimilar
pheno-typicproperties. Preston (32) has reported that
at least six immediate early polypeptides
(VP175,-136, -110,-68, -63,and -12) are
synthe-sized in vivo after release of a cycloheximide
block and that in vitro translation of immediate
early mRNA yields thesesameproducts.
Pres-tonhasalsocompared the synthesis of
immedi-ate early polypeptidesin tsK andwild-type
vi-rus-infected cells and has obtained evidence
which demonstrates that (i) immediate early
proteins are directlyinvolved in the transition
from immediateearlytoearly protein synthesis,
(ii) three forms of VP175 exist, and (iii) upon
temperature shiftdown, the multiple effects of
themutationsarereversible. Incontrast,Pereira
etal. (30) placed VP136 in the group ofearly
proteinsdespitethe fact that it issynthesizedin
the presence of cycloheximide or canavanine.
Thisclassificationwasbaseduponthe
observa-tion that VP136 wassynthesized slightly later
than the other immediateearly proteins.
To confirm that the product ofgene 1-2 is
indeed VP175 andtofurther define the function
of thisproteinin theregulationof viral
transcrip-tion, studieswereundertakentomapphysically
andgeneticallyaseries ofmutantsingroup 1-2
andtocharacterizetheresidualactivityof
mu-tantgene productsin infected cellsatthe
non-permissivetemperature.
Mapping studies. Previously, studies have shown thattsB21u (29), tsB2b (3), tsD(41, 42),
tsLB2 (21), and tsc75
(22)
lie in therepeated
sequencesbounding Us.Inthisstudy,we have
attemptedtorefine the mappositionsofseveral
mutantsin complementation group 1-2 and to separate them physically within these se-quences.
It is our contention that the data presented
heretogether with thefindingsof other
investi-gators place themutants within the structural
gene for VP175. The pertinent arguments for
thiscontentionare asfollows: the ninemutants
(i) fail to complement each other, (ii) exhibit
similarviralDNAandprotein phenotypes, (iii)
produceanaltered form of VP175, (iv)are
sep-arable by physicaland genetic mapping
proce-dures, and (v) map within the DNAsequences
which encode VP175. Moreover, VP175 is the
only protein which has been shown to be
en-codedwithin thesesequences, and (vi) a4.7-kb
immediateearly mRNA coding foraprotein of
molecular weight 175,000 also maps in these
sequences(6).Therefore,unless extensive splic-ing occurs in production of the message for
VP175, andnoevidencefor such splicing hasyet
beenreported,the ninemutants mustlie in the
structural gene for VP175. Mapping studies
therefore exclude the possibility that the
phe-notypic properties ofmutants ofgroup 1-2 are
due to mutations in a gene which regulates
VP175synthesis.
As described above (Results), mostmutants
map nearertheterminal "a"sequencesthan the
unique region of S. Two possible explanations
for this observation exist: (i)thissegmentof the
VP175 gene, ifmutated, results in significant,
observable phenotypic changes (e.g., this
seg-mentencodesanactivesite),or(ii) some
prop-erty of the DNA makes ithighlymutable. We
cannot testthe forner possibilityat this time;
however, the latter lends itself to speculation.
Knipe et al. (21) have demonstrated regionsof
identity between the terminal "ca"sequencesin
S and have proposed a copying mechanism
whereby,duringDNAreplication,oneterminus
serves asthe templatefor the other. They fur-therpostulatedthat "c"sequencesfurther from
the terminal "a" sequences can tolerate
se-quenceheterogeneitymorereadilythan the "c"
sequences nearer "a." Thus, the former
se-quences have been designated variable "c"
se-quences, and those nearer terminal "a"
se-quences have been designated invariable "c"
sequences. These investigators also
demon-strated that ts lesions in VP175 are recessive. Thus, sequences atonly one terminusneedbe
wildtype togenerate the wild-type phenotype,
whereassequencesatbothterminimustbe
mu-tant to generate the ts phenotype (21, 31, 33).
Our dataareconsistent with thishypothesisin
that a mutation induced in invariable "c"
se-quenceswouldbemorelikelytobecopiedinto the opposite terminus, yielding a mutant phe-VOL. 36, 1980
on November 10, 2019 by guest
http://jvi.asm.org/
200 DIXON AND SCHAFFER
notype, than would a mutation in variable "c"
sequences, which could produce a"silent"
mu-tant phenotype. Thus, perhaps, the copying
mechanism of DNA synthesis constitutes the
underlyingreasonfor the isolation ofa
predom-inance of ts mutations ininvariable,terminal"c" sequences. Further analysis of the remaining
members ofgroup 1-2 and efforts tointroduce
mutations into the variable and invariable
se-quences in oneterminus butnotthe other will
benecessary to prove thishypothesis.
Thegeneticmap of the nine mutantsexhibits
strikinglinearity and additivity, much more so
than linkage maps of markers in unique
se-quences in L (37; Dixon and Schaffer,
manu-script in preparation). Moreover, the
recombi-nation frequencies observedamong mutants in group 1-2 were muchhigherthanthose observed
between markers within a single gene in the
uniqueregion (Dixon andSchaffer, manuscript
inpreparation). Thediploid natureof thegene
may accountfor the high frequency of recom-bination sinceonlyoneterminus need recombine
toyield ats+ recombinant: crossovers ateither
end wouldproducetwice theeffective recombi-nationfrequency.
Another interesting observation which arose
from recombinationstudies concerns the
behav-ior of tsc75. This mutant recombines
ineffi-ciently with other members of group 1-2. In
addition, it behaves as if itwere closelylinked
totsD,yet it hasbeenmapped physicallyin the
terminal "a" sequences (20). A possible
expla-nation for thesefindingsis thatthe"a"sequence
may, infact, berepeated within "c"sequences.
Thissuggestionwasfirst madeby Wadsworthet
al. (44), who demonstrated small lariat
struc-tures at the ends of molecules which had been
digestedwith exonucleaseIII,andproposedthat
the sequencearrangement of the Sregion may be ...a'x'ac'---Us---ca'xa.We will assumefor the
sake ofargument that this arrangement is cor-rect. Ifthe "a"sequence isdiploidin eachrepeat
andif tsc75 doesliewithin the "a" sequence, at
least two copies of the mutant "a" sequence
wouldbepresent in order to preventself-rescue.
Although Knipe et al. (20) have proposed that
thecodingsequences for VP175includethe
ter-minal "a" sequence, this need not be the case. Infact, it may be a mutation in the internal "a" sequence which produces the mutant pheno-type, or perhaps mutations in both "a" se-quences. Ourgeneticandphysicalmapping data support this contention. If the mutation in the
internal "a" sequence were essential for the
expression of the ts phenotype, and ifthis
se-quence were adjacentto tsD, then the position oftsc75 foundbyrecombinationanalysis would
be predicted. Moreover, if "a" sequences were
also present in "c" sequences, one wouldpredict
that tsc75 would be rescued by fragment K2
(containing the terminal "a" sequence) and by
fragment K3 (containing the putative "a"
se-quencein"c"). Indeed,this is precisely the result weobtained.
Alternatively, the low frequency of
recombi-nation observed between tsc75 and other mem-bers of group 1-2 may reflect the fact that recom-bination occurs with decreasedfrequencyatthe ends of molecules.Thus, low recombination fre-quencies with markers in the terminal "a" se-quencecould result inmismapping.This expla-nation is inconsistent, however, with rescue of tsc75by both K2 and K3.
Function of VP175. Having demonstrated that all nine mutants in complementation group 1-2arelocated in thestructuralgeneforVP175,
further functional analysis of this polypeptide
was undertaken.
We first demonstrated thatall nine mutants exhibit similarphenotypes withregardto poly-peptide synthesis at the nonpermissive temper-ature. Minorvariationsintheamounts of certain polypeptides were observed, owing perhaps to strainvariation orleak,but in most casesVP175, -136, -110,-68, and -63 were seen to be overpro-duced. Intsc75-infectedcells,however, increased
amounts ofVP110 accumulated relative tothe
other mutants.Althoughthismaysimply reflect
a strain-specific difference, it may also reflect
someunusualproperties ofVP110.Clementset al. (6) and Watson etal. (48) have shown that VP110 is encodedby the terminalrepeats ofthe
Lregion. Because the "a"sequence is common totherepeatsof bothLandS and because the
mRNAcodingforVP110may contain aterminal
"a"sequence, it ispossible that tsc75 is mutant for both VP175 and VP110 and is,therefore, a
nonresolvable double mutant. If tsc75 contains
a mutation in the gene for VP110, then the
accumulationof VP110 inmutant-infected cells
mayreflectarole forVP110initsownregulation
or,alternatively,itmayreflectafailureto
proc-ess the mutant form of VP110 analogous to mutants in VP175. Because nosingle mutant in VP110 is available, we cannot distinguish be-tween the twopossibilities.
Severalgroups have reported thatfunctional
VP175 is required to switch from immediate
early to early protein synthesis and that the requirement is probably at the level of
transcrip-tion (7, 8, 13, 14,22, 24, 25, 32, 43, 47). What is not clear is the roleofearly proteinsinregulating
immediate early protein synthesis and the role ofVP175 inregulating its ownsynthesis. Tem-perature shiftup experiments have shown that VP175 aswell as otherimmediateearly proteins canbesynthesized in the presence of both early J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
HSV-1 ts MUTATIONS IN VP175 201 and late functional proteins, indicating that
VP175 isinvolved either directlyorindirectlyin
turningoff itsownsynthesis andperhaps that of
other immediateearly proteins. This conclusion was particularly apparent in the case of shifts
lateininfectionwhich resulted in the
reexpres-sion or increased synthesis of immediate early
proteins (Fig.4).If this isindeed the case, then
themodel ofcascaderegulation mustbe
modi-fiedtoinclude autoregulation of immediateearly
proteins in addition to feedback inhibition by
early proteins.
Anotherpoint tobe madeconcernsthe
exis-tence ofmultiple control groups in each
poly-peptide class. Pereiraet al. (30) firstsuggested
that subgroups of immediate early and early
proteinsexisted,aconceptsupported by Preston
(32). Our experiments have defined a group of
proteins whichareactivelateininfection (after
8h) andyetrequirethecontinuedexpressionof
VP175. This is apparent from the differences
seenin polypeptide synthesis aftershiftuplate
in infection and from the observation that no
viruswas producedeven thoughmostproteins
were synthesized (35). Inaddition, VP136, first
classifiedas animmediateearly protein by
Mars-denetal. (26)andanearly protein byPereiraet
al. (30), does not appear to be under VP175
control, yet it accumulates after VP175, -110,
and -63 reach maximum synthesis. Thus, as
pointedoutbyothers, VP136synthesisexhibits
thekinetics ofanearly proteinand the control
requirements ofanimmediateearly protein.
A model consistent with all available data
would require that all promoters on the HSV
genomebeactiveatalltimes but that promoters
differinthefrequencywith whichtranscription
isinitiated. PromotersforVP175, -110,and -63
would have the highest initiation frequencies,
whereas those for VP136 and other early and
lateproteins would havealowerfrequency,but
still berecognized by the
transcriptional
appa-ratus. One function of VP175 would be to
in-creasethefrequencyoftranscriptionfromsome
earlyor even late promotersand decrease the
frequency from its own promoters. Consistent
withthis model arethe
following
observations:(i) certain late proteins (e.g.,
VP154)
can beexpressed in the absence of functional VP175
(this study), (ii) VP175 is
required
for certainlate viralfunctions (thisstudy), (iii)theresident
HSV TK gene in biochemically transformed
cells is expressed in the absence of VP175 (19,
24,25), (iv) functionalVP175isabletoenhance
HSV TK expression in HSV TK-transformed
cells (19, 24, 25), (v) VP175 is
reexpressed
ingroup 1-2mutant-infected cellsafter
shiftup (i.e.,
after inactivation of theautoregulatory function
ofVP175) (thisstudy),and(vi) theactive form
of VP175 has DNA binding properties (49). If
thismodelwerecorrect, onemightexpect VP175
tobind directlyto promoter sequences or
per-haps toform complexes with the RNA polym-erase toalteritsspecificity. Thistranscriptional
model is similarin many ways to theregulatory
mechanism operative during adenovirus
repli-cation inwhich pre-early proteinsare required
toinitiate the synthesis of early proteins,which
in turn initiate later protein synthesis (2, 16).
However, duringtranscription in vitro,the ma-jorlatepromoter(s) is recognized in preference
to the early promoter(s) (J. Manley, personal
communication). This indicates that although
early functionsareneededtoenhance late
pro-moter activity, they can be bypassed by the cellular transcriptional complex.
ACKNOWLEDGMENTS
We thank L. B. Sandner and P. A.Temple forexcellent
technical assistance, J. G. Driscoll forpreparation of the manuscript, and D. M. Coen forhelpful discussions and sug-gestions. We areespecially indebted to R. J. Courtney for constructive comments and continued interest and
encourage-ment.
This investigation wassupported byPublic Health Service research grant CA20260 and programproject grant CA21082 from the National Cancer Institute.
LITERATURE CITED
1. Becker, Y., A. Dym, and Y. Sarov. 1968. Herpes simplex virus DNA. Virology36:184-192.
2. Berk, A. J., F.Lee,T. Harrison, J.Williams,and P. A.Sharp. 1979. Pre-early adenovirus5geneproduct regulates synthesis of early viral mRNAs.Cell
17:935-944.
3. Bookout, J. B., P. A. Schaffer, D. J. M. Purifoy, and N.Biswal. 1978.Marker rescue of temperature-sensi-tive mutants by defectemperature-sensi-tive DNA of herpessimplexvirus type1.Virology87:528-538.
4. Brown, S.M.,D. A.Ritchie, and J. H. Subak-Sharp. 1973.Geneticstudieswith herpes simplex virus type 1: the isolation of temperature-sensitive mutants, their arrangementintocomplementationgroups,and recom-binationleading to alinkagemap. J. Gen. Virol. 18:
329-346.
5. Chu, C.-T.,D. S.Parris, R. A. F. Dixon, F. E. Farber,
andP.A.Schaffer.1979.Hydroxylaminemutagenesis of HSV DNA and DNA fragments: introduction of mutations into selected regions of the viral genome. Virology 98:168-181.
6. Clements,J.B., J. McLauchlan, and D. J. McGeoch.
1979.Orientation ofherpes simplex virus type 1 imme-diateearlymRNAs.Nucleic Acids Res. 7:77-91.
7. Clements,J.B., R. J. Watson, and N.M. Wilkie. 1977.
Temporal regulation of herpes simplex virus type 1
transcription: location oftranscriptson the viral
ge-nome.Cell 12:275-285.
8. Courtney, R. J., and M.Benyesh-Melnick.1974. Iso-lationandcharacterizationof alarge molecular weight polypeptideofherpessimplexvirustype 1. Virology62: 539-551.
9. Courtney,R.J., P. A. Schaffer, and K. L. Powell.
1976. Synthesis of virus-specific polypeptides by tem-perature-sensitivemutantsofherpes simplex virus type
1.Virology75:306-318.
10. Delius,J., and J.B.Clements.1976. Apartial
denatur-ationmap ofherpes simplexvirustype1:evidence for VOL. 36, 1980
on November 10, 2019 by guest
http://jvi.asm.org/
202 DIXON AND SCHAFFER
inversions of the two unique DNA regions. J. Gen. Virol. 33:125-133.
11. Graham, B. L., H. Ludwig, D. L. Bronson,M. Ben-yesh-Melnick, and N.Biswal. 1972. Physicochemical properties of the DNA of herpesviruses. Biochim. Bio-phys. Acta 254:13-22.
12. Halliburton, I.W.,R. E.Randall, R. A.Killington, and D. H. Watson.1977.Someproperties of intertypic recombinants between type1and type 2herpessimplex
virus.J. Gen. Virol. 36:471-484.
13. Honess, R.W.,and B. Roizman. 1974.Regulationof herpesvirusmacromolecularsynthesis.I. Cascade reg-ulation of thesynthesisof threegroups of viralproteins.
J.Virol. 14:8-19.
14. Honess,R.W.,and B. Roizman. 1975. Regulationof herpesvirusmacromolecular synthesis: sequential tran-sition ofpolypeptidesynthesis requiresfunctional viral polypeptides. Proc.Natl. Acad. Sci. U.S.A. 72:1276-1280.
15. Hughes,R.G.,and W. H.Munyon.1975.
Temperature-sensitivemutantsofherpessimplexvirustype1 defec-tive in lysis but not in transformation. J. Virol. 16:275-283.
16. Jones,N., and T. Shenk. 1979. Anadenovirustype 5 early gene functionregulates expressionof other early viral genes. Proc. Natl. Acad. Sci.U.S.A.76:3665-3669. 17. Jones, P.C., G. S.Hayward,and B. Roizman. 1977. Anatomy ofherpessimplexvirus DNA.VII. aRNA is
homologoustononcontiguoussites in both the L andS component of viral DNA. J.Virol. 21:268-276. 18.Kieff, E.D., S.L.Bachenheimer, andB.Roizman.
1971. Size, composition, and structure ofthe deoxyri-bonucleic acid ofherpessimplexvirussubtypes1and 2. J.Virol. 8:125-132.
19. Kit,S., D. R.Dubbs,and P.A. Schaffer. 1978.
Thy-midine kinase activity of biochemically-transformed
mouse cellsaftersuperinfectionby thymidine kinase negative temperature-sensitive herpes simplex virus mutants.Virology85:456-463.
20. Knipe, D. M., W. T.Ruyechan,R.W.Honess, and B.
Roizman.1979.Moleculargeneticsofherpessimplex virus: theterminal a sequences of the L and S compo-nents areobligatorilyidentical andconstitute a part of astructural genemapping predominantlyin theS com-ponent. Proc.Natl. Acad.Sci. U.S.A.76:4534-4538.
21.Knipe,D.M.,W. T.Ruyechen,B.Roizman,and I. W.
Halliburton. 1978.Moleculargeneticsofherpes sim-plex virus: demonstration ofregionsofobligatoryand nonobligatory identity within diploid regions of the genomeby sequencereplacementand insertion. Proc. Natl. Acad.Sci.U.S.A.75:3896-3900.
22. Kozak, M.,andB.Roizman. 1974.Regulationof her-pesvirus macromolecularsynthesis:nuclear retention of
nontranslatedviral RNAsequences. Proc. Natl. Acad. Sci. U.S.A.71:4322-4326.
23. Laemmli, U. K. 1970. Cleavage ofstructuralproteins
during theassembly of the head of bacteriophage T4. Nature(London)227:680-684.
24. Leiden, J.M., R. Buttyan, and P. G. Spear. 1976. Herpes simplex virus geneexpressionintransformed
cells.I.Regulation of theviralthymidine kinase gene in
transformedLcellsbyproducts of superinfecting virus. J. Virol.20:413-424.
25. Leung,W.C.1978.Evidencefor aherpessimplex
virus-specificfactorcontrollingthetranscription of deoxy-pyrimidine kinase. J. Virol. 27:269-274.
26. Marsden, H. S., I. K. Crombie, and J. H.
Subak-Sharpe.1976.Controlofproteinsynthesis in
herpes-virus-infected cells:analysis of the polypeptides induced by wild-type and sixteen temperature-sensitive mutants of strain17.J. Gen. Virol. 31:347-372.
27. Morse,L. S.,L.Pereira, B.Roizman, and P. A. Schaf-fer. 1978.Anatomyofherpessimplex virus DNA. X.
Mapping of viral genes byanalysis of polypeptides and functionsspecified by HSV-1 xHSV-2 recombinants. J. Virol. 26:389410.
28. Parris, D. S., R. J.Courtney,and P. A. Schaffer. 1978. Temperature-sensitive mutantsof herpes simplex virus type 1 defective in transcriptionaland post-transcrip-tionalfunctions required for viral DNA synthesis. Vi-rology90:177-186.
29. Parris,D.S.,R.A. F.Dixon,and P. A. Schaffer. 1980. Physical mapping of herpes simplex virus type 1 ts mutants by marker rescue:correlation of thephysical andgenetic maps. Virology 100:275-287.
30. Pereira,L., M. H.Wolff,M.Fenwick,and B. Roiz-man. 1977.Regulation of herpesvirus macromolecular synthesis. V.Properties of a polypeptides made in HSV-1 andHSV-2 infected cells. Virology 77:733-749. 31. Preston,C. M. 1979. Abnormal properties of an
immedi-ateearlypolypeptide in cells infected with herpes sim-plexvirus type 1 mutanttsK. J. Virol. 32:257-269. 32. Preston,C. M. 1979. Control of herpes simplex virus type
1 mRNA synthesis incellsinfected with wild-type virus or the temperature-sensitive mutanttsK. J. Virol. 24: 275-284.
33. Preston, V.G., A. S. Davison, H. S. Marsden, M. C.
Timbury,J. H. Subak-Sharpe,and N. M.Wilkie.
1978.Recombinants between herpes simplex virus types 1and 2: analysis of genome structure and expression of immediate early polypeptides. J. Virol. 28:499-517. 34. Schaffer,P.A., G. M.Aron,N.Biswal,and M.
Ben-yesh-Melnick. 1973. Temperature-sensitive mutants of herpessimplex virustype 1: isolation,complementation andpartial characterization. Virology 52:57-71. 35. Schaffer,P.A.,D.R.Bone,and R. J.Courtney.1976.
DNAnegativetemperature-sensitive mutants of herpes simplex virus type 1:patternsofviral DNAsynthesis
after temperature shift-up. J. Viro.17:1043-1048. 36. Schaffer,P.A.,V.C.Carter,and M. C.Timbury.1978.
Collaborative complementation study of temperature-sensitive mutants of herpes simplex virus types 1 and 2. J.Virol. 27:490-504.
37. Schaffer, P. A., M. J. Tevethia, and M.
Benyesh-Melnick. 1974. Recombination between temperature-sensitive mutants of herpes simplex virus type 1. Virol-ogy58:218-228.
38. Sheldrick,P., and N. Berthelot. 1974. Inverted repeti-tions in the chromosome ofherpes simplex virus. Cold
SpringHarborSymp.Quant. Biol.39:667-681. 39. Skare, J., and W. C.Summers. 1977. Structureand
function ofherpesvirus genomes II. Eco RI, XbaI,and Hind IIIendonuclease cleavage sites on herpessimplex
type 1 DNA.Virology 76:581-595.
40. Smith, K. 0. 1964.Relationship between the envelope and theinfectivityofherpessimplexvirus. Proc. Soc. Exp. Biol. Med. 115:814-816.
41. Stow, N. D., J. H.Subak-Sharpe, and N. M. Wilkie. 1978.Physical mapping of herpes simplex virus type 1 mutationsby markerrescue.J.Virol. 28:182-192. 42. Stow, N. D., and N. M.Wilkie.1978. Physicalmapping
oftemperature-sensitive mutants of herpes simplex vi-rustype 1 by intertypic marker rescue. Virology
90:1-11.
43. Swanstrom,R.C., and E. K. Wagner. 1974. Regulation ofsynthesis ofherpes simplex virus type 1 viral mRNA during productive infection. Virology 60:522-533. 44. Wadsworth,S.,R. J.Jacob, and B. Roizman. 1975.
Anatomy of herpes simplex virus DNA.II. Size, com-position and arrangement of inverted terminal repeti-tions. J.Virol. 15:1487-1497.
45. Wagner,M.M.,and W.C. Summers.1978.Structure of thejoint region and termini of the DNA of herpes simplex virus type1.J.Virol. 27:374-387.
46. Ward, R. L, and J. G. Stevens. 1975. Lifetimes of mRNA moleculesdirecting the synthesis ofviral pro-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 36, 1980 HSV-1 ts MUTATIONS IN VP175 203
teins inherpessimplexvirus-infected cells. J.Virol.15: 1979.Separationand characterization ofherpes simplex
81-89. virus type 1immediateearlymRNAs. J.Virol.
31:42-47.Watson, R.J.,and J. B. Clements. 1978. Characteri- 52.
zationoftransciption-deficienttemperature-sensitive 49. Wilcox, K.W., A. Kohn, E.Sklynskaya,and B. Roiz-mutants of herpes simplex virus type 1. Virology 91: man. 1980. Herpes simplex virusphosphoproteins. I. 364-379. Phosphatecycles on and off some viral polypeptides 48.Watson, R.J., C. M. Preston, and J. B. Clements. and can alter theiraffinity for DNA. J. Virol. 3:167-182.
on November 10, 2019 by guest
http://jvi.asm.org/