JOURNAL OF VIROLOGY, Jan. 1994,p. 390-399 Vol. 68, No. I 0022-538X/94/$04.00+0
Copyright ©D 1994,AmericanSociety for Microbiology
Apoptosis
Induced in
CD4+
Cells
Expressing
gpl60 of Human
Immunodeficiency
Virus
Type 1
YI-YU LU,' YASUHIROKOGA,'* KAZUOTANAKA,2 MASAFUMI SASAKI,2 GENKI KIMURA,2 AND KIKUO NOMOTO'
Departments ofImmunology' and Virology,2 MedicalInstituteof Bioregulation, Kyushu University 69, Maidashi,
Higashi-ku,
Fukuoka 812,Japan
Received 26 July 1993/Accepted 5 October 1993
Ina previous study (Y. Koga, M.Sasaki, H. Yoshida, H.Wigzell, G.Kimura,and K.Nomoto, J.Immunol.
144:94-102, 1990),we demonstrated that the expression ofgpl60,aprecursor formofenvelopeglycoproteinof human immunodeficiency virus type 1, inCD4+ cells causes thedownregulationofsurface CD4 andsingle-cell
killing by forming intracellulargpl60-CD4complex. In the presentstudywe investigatedthe events that lead to cell death inCD4+ cells expressinggpl60.We found thatapoptosisis induced in cellsundergoingsingle-cell death. Moreover, even the cell clone, which expresses so littlegpl60 that it does not exhibit any apparent cytopathic effects, such as the inhibition of cell growth, was found to be highly susceptibleto theapoptosis
induction by the anti-Fas monoclonal antibody.
Infection withhumanimmunodeficiency virus (HIV)causes anextensivedepletion of CD4+ helper T cells,resultinginthe destruction of the host defense mechanism composed of
cellular and humoral immune responses (6, 7). HIV exerts a
cytopathic effecton single Tcells in culture through a mech-anism independent of syncytium formation (25, 26). Although the mechanism of single-T-cell killing as a result of infection withHIV still remains to beelucidated, it mayplay acrucial role inthe depletion of CD4+ T cells in patients withAIDS. Recentestimatesof the viral burden in the lymphoid organs of
HIV-infected individualsalso support thepossibilitythat virus-mediated cell killing itself, without the participation of any
immune responses against infected cells or syncytium
forma-tion,accounts for the CD4+T-cell depletion in vivo (5, 20). The mechanism by which the virus directly kills infected CD4+ cells has not yet been precisely defined. An unusual featureof HIVinfection is the accumulationof alargeamount
of unintegrated viral DNA in the infected cells, and this phenomenon hasalso beensuggestedtobeanimportant factor in the cytopathic effect ofHIV(24). Massive viral buddingmay result incell death by injuringthesurface cell membrane (26).
It is also speculated that infection with HIV may induce terminaldifferentiation of the infected CD4+Tcells, resulting
incell death (30).
On theotherhand, there is accumulating evidence that both
theCD4moleculeand the virus envelope glycoprotein play an
important role inthe cytopathic effect of infected CD4+ cells. In previous studies (10, 14), we showed that the intracellular
expression ofgpl60, a precursor form ofthe HIV envelope
glycoprotein, in CD4+ cells causes the downregulation of
surface CD4 and single-cell death. An analysis of those cells
demonstrated the intracellular accumulation ofanaggregated form ofgpl60bound to CD4 and the direct participation of
suchagpl60-CD4complexinsingle-cellkilling following CD4
downregulation.
In the present study, we attempted to determine exactly which events lead to cell death in the CD4+ cells expressing
*Corresponding author. Mailing address: Department of Infectious
Diseases, School of Medicine, Tokai University, Bohseidai, Isehara, Kanagawa 259-11, Japan. Phone: 463-93-1121, ext. 2591. Fax: 463-94-2976.
gpl60 andfoundthatsuch cellsundergo apoptosis,which is an
internally programmed cell death pathway characterized by
cleavage ofDNA at the internucleosomal regions. Moreover,
eventhe cellclone, inwhich gpl60 expressionis so low that it is unable to either killcells or inhibit cellgrowth, was highly susceptible toapoptosis inductionby the anti-Fasmonoclonal
antibody (MAb).
MATERIALS ANDMETHODS
Construction of HIV env expression plasmids. Plasmid
pSMTE7wasconstructedbyinserting thepromoterportion of the human metallothionein IIA gene and the sequence of whole HIV env (BH1O clone [22], nucleotides 5496 through 8474) into plasmid pSV2neo, which has a gene encoding
resistance toG418.Agpl60-expressing plasmid, pSMTE7-160,
and a gpl2O-expressing plasmid, pSMTE7-120, were
con-structed by introducing mutagenesis into the env region of pSMTE7 bytheoligonucleotide-directed mutagenesis method of Kunkel (23), using mismatched 24-mer synthetic DNA
primers 5'TAT1CCCACTGCGC7 Fri--iCTCT3' and 5'T
ATTCCCACTGCTCATTTTTCTCT3', respectively. These primers correspondtonucleotides7323 to 7346of theenvgene
ofBH1O, whilechanging Arg-518 (BH10 numbering) ofenvin
pSMTE7 into Ser in pSMTE7-160 and into a stop codon in pSMTE7-120, respectively. Suchchanges of nucleotides were confirmed by nucleotidesequence analysis.
Cells.U937 clone2(U937-2),ahumanmonocytoid cellline
expressingahighlevel of surfaceCD4(1),wasused asthe host for transfection with each of the HIV env expression vectors
by the electroporation method (21). The cells were then selected for resistance to I mgof G418 permlin the absence
of any heavy metal ions to get clones stably integrated with
those vectors. Such cell clones were maintained in RPMI 1640 medium supplemented with 10% fetal bovine serum, 5 mM N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid (HEPES), 100 U of penicillin G per ml, and 100 p.g of streptomycin sulfate per ml in humidified air containing 5%
CO2. The number of cells was determined with a Coulter counter(model ZB;CoulterElectronics Inc., Luton,England).
Cell viability was examined by the exclusion of 0.1% trypan blue in the culture.
390
on November 9, 2019 by guest
http://jvi.asm.org/
APOPTOSIS BY gp160 of HIV-1 391
1
2
_ f
3
kDa
I-.
b3
-160P -120
-40
FIG. 1. Immunoblot analysis by anti-Env MAb. UE160-16 cells (lanes 1), UE160 cells (lanes 2), and UE120 cells (lanes 3) were cultured for 4 h eitherwith (+ lanes) or without (- lanes) 10 ,uM CdCl2.A100-p.gsample of the celllysateprotein prepared from such cell samples was run on an SDS-polyacrylamide gel electrophoresis (10% polyacrylamide) gel, blotted onto a nitrocellulose membrane, reactedwith0.5f MAb,and developed by the ECL system.
Reagents. A murine anti-Fas MAb, clone CH-1 1 (29), was
purchased from MBL, Nagoya, Japan. A recombinant human tumornecrosis factor alpha (TNF-ac), which is derived from
thefull-length human TNF-o gene, waspurchasedfrom Wako Pure Chemical Industries, Osaka, Japan. The definition of 1
unit used in thepresentstudy is theamountofTNF-otrequired
tomediate the half-maximal cytotoxicityof L929 cells.
Immunoblot analysis. The cells were washed once with phosphate-buffered saline (PBS) and lysed by sonication in
lysis buffer consisting of 0.5% Nonidet P-40, 0.5% deoxy-cholate, 20 mM Tris-HCl (pH 8.0), 120 mM NaCl, 2 mM EDTA, 2 mMphenylmethylsulfonylfluoride, 10 F.Maprotinin,
and 10,uwMleupeptin.Bythisprocedure, 106cellsyieldedacell
lysatecontaining about 200 pgofprotein.A 100-,ugsample of each cell lysate was run on a sodium dodecyl sulfate (SDS)-10% polyacrylamide gel, blotted onto a nitrocellulose
mem-brane, reacted with 0.5 ,ug of0.5,B,an anti-envelope glycopro-tein MAb (17), per ml at room temperature, and then was developed by the ECLsystem (enhancedchemiluminescence;
Amersham Corp., Amersham, England) as specified by the
manufacturer. The relative density of the protein bands ex-posed on X-ray film was measured quantitatively with a densitometer (dual-wavelengthchromatoscannerCS-930; Shi-madzu, Tokyo, Japan). To measure the amountsofenvelope
glycoproteins detected in the immunoblot analysis, adefinite
amount of purified gpl20 or gpl60 was serially diluted and loaded intoparallellanesasthestandard,andthe amountsof envelope glycoproteins in the sampleswere then assessed by
using a densitometer within the range overwhich the linear dose relation is available. Standard gpl20 and gpl60 were purchased from MicroGeneSys, Inc., Meriden, Conn.
DNA fragmentation assay. DNA fragmentation was
deter-TABLE 1. Level ofenvelope glycoproteinin cell clones
Treatmentwith Amt
(,ug/10`
cells) in:CdCl,
UEI60-16
UE160)
UEl20
No ND"' 0.06 ND
Yes 0.03 0.41 0.33
"ND,notdetected.
mined by the method of Newell et al. (18). Briefly, 5 x l0" cells were washed once with PBS and lysed in 0.5 ml of hypotonic lysing buffer (5 mM Tris-HCl [pH 7.4], 5 mM EDTA, 0.5% Triton X-100). The lysates were centrifuged at 13,000 x g for 15 min. Their supernatants were incubated with RNase at 50 p.g/ml for 2 h and deproteinized by extraction
once in phenol-chloroform and once in chloroform-isoamyl
alcohol(24:1).The supernatant was then precipitated at 10,000 x gafterbeingleft overnight at -20°C in 50% isopropanol-130 mM NaCl.Afterelectrophoresison 2%agarose slabgels, the DNA wasstainedwith ethidium bromide.The low-molec-ular-weight DNAs in the supernatant were quantitated by a
diphenylamineassay (2).
Detection ofapoptosis by flowcytometry. The cells
under-going apoptosis were examined by flow cytometry by the
methodofLyons etal.(16).Inbrief, the cellswerestained with
1 ,ugof ethidiumbromide per ml inPBSand were kept for 30
min onice in the dark. Afterbeing washedwith cold PBS, a
total of 10,000 cells were analyzed by using an EPICS Elite
(Coulter, Hialeah, Fla.) with the logarithmic amplification of red fluorescence detection and linear amplification of the forward scatter.
Electronmicroscopy.Thecells (2 x 107)weresuspendedin 5ml of PBS,combined with5 mlof2 x fixing buffer (1x fixing buffer is2.5%glutaraldehyde, 0.1 M sucrose, and 3 mMCaCl,
in 0.1 M cacodylate buffer [pH 7.4]), and kept for 10 min at
37°C. Thecellpelletswereprocessed through 1%OS04for30 min at 4°C, dehydrated in graded ethanol and in propylene oxide, and embedded in Epon 812 resin. Thin sections were stained with2% uranylacetatefor60 minandlead acetatefor
5 min and thereafter were examined under a JEM-2000EX electron microscope.
RESULTS
Expression of envelope glycoproteins by transfectant cells. The cell clones, precultured for 4 h either with (+ lanes) or
101
2
-J -J
LU. U
51
1
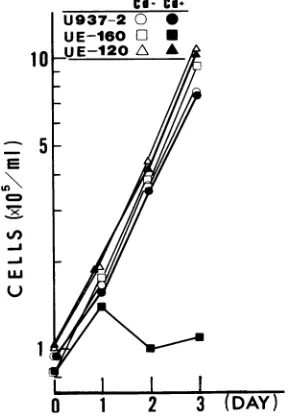
FIG. 2. Cellgrowthafter theexpressionofenvelopeglycoprotein. U937-2, UE160, and UE120 cellswereadjusted to105 cells in1 ml of RPMI 1640mediumonday0 andwereculturedeither withorwithout 10,uM CdCI. Ondays0, 1,2,and3, thecell number in each culture wasdetermined withaCoultercounter.
VOL. 68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.121.230.75.214.2] [image:2.612.364.508.468.676.2]392 LU ET AL.
[image:3.612.74.544.88.666.2]U937-2
UE-
160
UE-120
FIG. 3. Microscopyof the cellculture. U937-2, UE160,and UE120cellswereadjusted to 105 cells in 1 mlofRPMI 1640medium andwere
cultured inthepresence of 10,uM CdCl2.At0,8,24,and 48hof culture, theviability ofthecells ineach culturewasexaminedby the exclusion
of0.1%trypan blueby usinganinverted microscope.
Oh
8h
24h
48h
J. VIROL.
11
on November 9, 2019 by guest
http://jvi.asm.org/
APOPTOSIS BY gpl6O of HIV-1 393
a
U937-2
M
0 4 8 24
UE120
0
4
8
24
UE160
0
4 8 24M
b
JME4
0
8 24 M
bP _-1353
_-311 -281
bp _281
--234
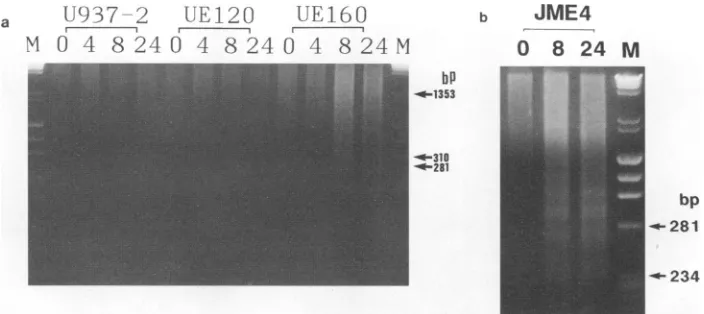
FIG. 4. DNAfragmentation in the cells after induction. (a) U937-2,UE120,andUE160 cells were cultured in the presence of 10 ,uM CdC12 for0, 4,8,or 24h(indicatedatthe top of each lane). Small supernatantDNA was isolated from 5 x 106cells of such cellsamples and then separated on a 2% agarose gel. Lane M contained a standard mixture ofHaeIII-digested4X174DNA andHindIll-digested X DNA. (b) JME4 cells were cultured in the presence of 201iMCdCl2for 0, 8, or 24 h (indicated at the top of each lane).The samples were then treated as described above.
without(- lanes) the addition of 10 ,uM CdCl2 as an inducer, were examined for their expression level of envelope glyco-protein by an immunoblot analysis with 0.5, (Fig. 1). The UE160-16cellsexpresseda160-kDaprotein band correspond-ing togpl60at 4hafter the addition of inducer; however, no detectable amount of gpl60 without induction was observed (lanes 1). The UE160 cells already contained easily detectable amounts of gpl60 without induction, whereas at 4 h after induction, the expression level of gpl60 increased 6.8 times morethan thatwithout induction and reached a peak (lanes 2). The UE120 cells expressed a significant amount of 120-kDa protein corresponding to gpl20 after induction, whereas no gpl20 was detectable without induction. The amounts of envelope glycoproteins in these cell clones were estimated quantitatively and are shown in Table 1.
Cytopathic effect by gpl60. As reported by us elsewhere (14), CD4+ cell clones,including those of monocyte and T-cell origins such as U2ME7, JME2, and JME4, demonstrate an
extensive cytopathic effect leading to single-cell death when gpl60 is expressed in the cells. In the present study, UE160 cells stopped proliferation and exhibited marked single-cell death after 48 h whentheywerecultured in the presence of 10 ,uMCdCl2 (Fig.2and3). AlthoughUE160cells expressasmall
amount ofgpl60 (0.06 [.g/106 cells) without induction, they proliferatedaswellasdidparental U937-2 cells in the absence ofan inducer(Fig.2). Thus it is thought that if the amountof
gpl60is 0.06
pLg
orless,thetransfectant cells of U937-2origincan tolerate the cytotoxic effect exerted by gpl60 and their growth doesnotsuffer any inhibition. Actually, UE160-16 cells, another U937-2-derived cell clone integrated with pSMTE7-160, express 0.03 ,ug ofgpl60afterinduction;they grewaswell
asU937-2cells foratleast4daysevenin the presence of the inducer(see Fig. 10C andD). UE120cells,which express 0.33 ,ug ofgpl20per
106
cellsbutnogpl60afterinduction,did notshow anygrowth inhibitionorapparentcytopathiceffect(Fig. 2 and 3), whereas the Leu3a/OKT4A but not the OKT4 epitopesonall the surface CD4s of these cellswereoccupied
bygpl20asindicated by examinationby flow cytometry(data not shown).
Apoptosis induced in UE160 cells. To examine whether apoptosis is the death pathway of UE160 cells when the expression levelofgpl60is increased in thesecells,theDNA
of cells was extracted and examined by agarose gel electro-phoresis (Fig. 4). At 8 and 24 h after induction, a DNA fragmentation pattern in multiples of about 200 bp of oligo-nucleosome length unit was shown in the UE160 cells but not in either the U937-2 or the UE120 cells (Fig. 4a). Such fragmentation ofgenomic DNAwasalsoseenin JME4(14),a
human T cell cloneexpressinggpl60under thetranscriptional control of hMTIIA promoter(Fig. 4b). Although the fragmen-tation ofDNA wasalready apparentas earlyas 8h after the induction in UE160 cells, thesecellswerescarcely stained with the trypan blue dye, which penetrates the injured plasma membrane butis excluded from the intactplasma membrane
(Fig. 3). This indicates that the surface cell membrane is not
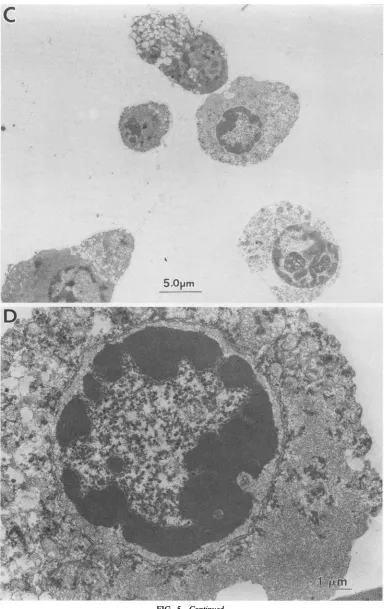
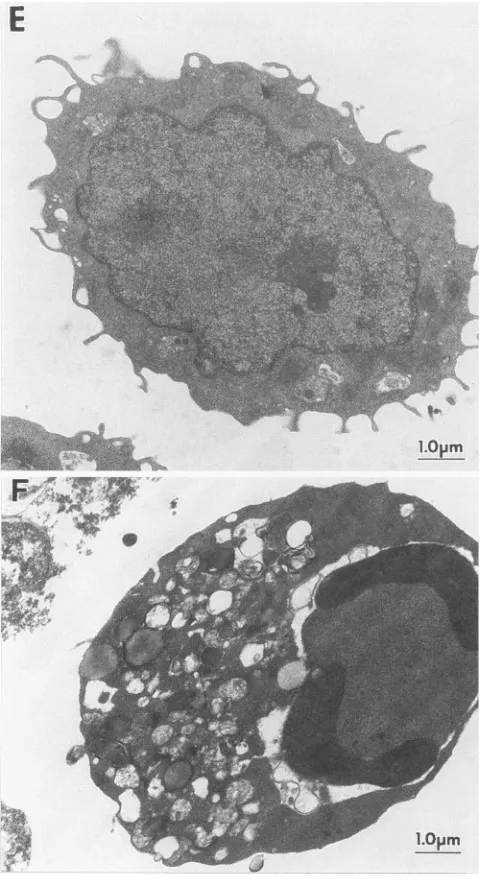
theprimary target of cytopathic effect raised in UE160 cells. These cell samples were examined by transmission electron microscopy at 24 h after induction (Fig. 5). The nuclei of UE160 cells exhibited typical morphology of apoptosis; i.e.,
their chromatin became very dense and formed crescents in tight apposition to the nuclear envelope (Fig. 5C and D). Likewise, the JME4 cells showed thick and crescent chromatin when their cells were treated with inducer (Fig. SF). On the otherhand, the UE937-2 cells didnotshow anymorphological
hallmarks ofapoptosis (Fig. 5A andB).Using flow cytometry, Lyonset al.(16) reported that the cellssufferingapoptosisat
its early stages stained faintly with ethidium bromide and exhibited a lowered light scatter. Such an early-apoptosis
patternwasobserved notonly in 22.7% of UE160 cellsat8h after induction but alsoin5.0%of thesecellswithout induction
(Fig.6, gateC). On the other hand, almost all the U937-2 cells showedanonapoptosis patternconsistingofnonstainingwith ethidium bromide and also exhibited a higher forward light scatterboth before and afterinduction(gateB).These results
suggestedthat the UE160cells,which expressasmallamount ofgpl60withoutinduction,havegoneto apreapoptosisstage in the absence ofinduction, although theygrewaswellasdid
parental U937-2 cells (Fig. 2).
Cytopathic effect by anti-Fas MAb. The susceptibility of
UE160 cells to thecytocidaleffect of anti-Fas
MAb,
which isknown to induce apoptosis, was examined by
measuring
the cell growth in the absence of inducer(Fig. 7).
As mentioned above,theUE160cells(Fig. 7B) proliferateaswellasdo theU937-2 cells(Fig. 7A)in the absenceof inducer. When these VOL.68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.130.482.72.229.2]394 LU ET AL.
I:
"
I 8 1
...
..
....u.-
.... . . ...FIG. 5. Electron microscopyof the cellsundergoing apoptosis.U937-2 cells(AandB)andUE160cells(CandD)were cultured with 10K.M
CdCl2for 24handexaminedbytransmission electronmicroscopy.PanelsB and D arehigher magnificationsofpanelsA andC,respectively.JME4
cellswerecultured for24 h with(F) orwithout (E)20 ,uMCdCl2beforebeing sampledfor electronmicroscopy.
B'd.
.*X
t.f,.g
>
t
X8
_
t@<i X,
a,:->ky,, _
70if
;}Ng4.ts'#S
iD,iS
'*-
;!_
t0f, tE}SHA
f; '-S7X:s:<'9.g5_
tS WX ': "0. id
t; ;0'.ff'_
fffff0000 t00 at00S_
f: 'A:0"'' s
w
.t:
1
um
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
APOPTOSIS BY gp160 of HIV-1 395
,
^ts,
*+
,sY'"SS>
.
;
07
Et
Se<WS-.
eSw
H;0-.
+
w
*,,.
-0
.iCw,.tS,.
l,
'f'-'s;';0*-';r
...s
0
't''
|;0
9WiV;0
0
'
f:
0
;
,+Wtet Disk
stU
,,
7
+,X:,5
Fz04,t,
7
0
0'X
05
7
'
<
§
*
,- -' :< ;a >; # r
w*,, ,, 0:0 L' C:: i. sLff
.''
'
A
'to_
t
X
d
s1s!_
r t'w_
*; < _tf
50
s3
yk
.
pm
s
si£x|j5_ >DD_
FIG. 5-Continued.
,W*
-,,,- ,
VOL.68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.113.498.90.699.2]396 LU ET AL.
E
ml I-0
a
4c
0 U.6
FIG. 5-Continued.
cellswere culturedwith CH-11 anti-Fas MAb, the growth of
theUE160cellswasmarkedly inhibited only with 0.5ngof this
MAbperml(Fig. 7B); however,thegrowthoftheU937-2 cells
wasnotblockedevenwithasmuch as 10ngofthis MAbper
ml(Fig. 7A).Both theagarosegelelectrophoresis(Fig. 8)and
diphenylamine assay (Table 2) demonstrated that extensive
apoptosis occurred in the UE160 cells after the addition of
CH-1 1 but that it was less dramatic in the U937-2 cells. In
contrast,thecytotoxic effectofrecombinantTNF-otwasnotso
marked inUE160 cellsevenwith the additionof 100 Uof this
cytokineperml(Fig. 7D),and itwasnegligible in the U937-2
cells (Fig.7C).The expressionlevel of surfaceFasantigenwas
almost thesamefor theUE160(Fig. 9A) and U937-2 (Fig. 9B)
cells by flowcytometry analysis with CH-11.
Susceptibility to anti-Fas MAb is gpl6O dependent. To determinewhether the susceptibility of cells to the cytocidal effect of anti-Fas MAb is related to the amount of gpl60
contained in thecells,weexaminedthe effect of CH-11onthe
U937
UE160
Cd(-)
Cd(+)
LOGRED FLUORESCENCE
FIG. 6. Flow cytometry analysis of cells undergoing apoptosis. U937-2andUE160cellswereculturedfor 8 h eitherwith[Cd(+)]or without [Cd( -)] 10 ,uMCdCl2. Then cellsampleswerestainedwith ethidium bromide and analyzed by flow cytometry. Horizontal and vertical axes denotelogredfluorescence from ethidium bromideand forward scatter, respectively. The cells in gate B in each panel demonstrate viable cells. The cells in gateC,whichareshrunk andgive off higherredfluorescence,representpreapoptotic cells.
growth of UE160-16 in the absence and the presence of
inducer(Fig. 10). Theamountof gpl60inUE160-16 cellsafter
induction with10 pLM CdCl2 was 0.03 p.g/106 cells,which was
almost thesame as that ofgpl60 in the UE160 cellswithout induction(Table 1). In the absenceofinducer, the inhibitory
effect of CH-11 on cell growthwas also not so significant in
either U937-2(Fig.10A)orUE160-16 (Fig.10B) cells; i.e., 1.0 ng of CH-11 per ml did not block the proliferation ofboth typesofcell, whereas this MAb retarded their growth rateto
about half ofthegrowthratewithout CH-11. Inthe presence
of inducer, however, a marked inhibition of cell growth by CH-11wasfound in the UE160-16 cells(Fig. 10D) while the
growth inhibition bythis MAb was still minimal in theU937-2
cells (Fig. 10C), indicating that the susceptibility of cells to anti-Fas MAb isdependentonthe amount ofgpl60 expressed in the cells.
DISCUSSION
Apoptosishas beendemonstratedtooccur intissues
under-going processes asdiverse as embryogenesis, metamorphosis, andimmunological development,inwhich the selective
deple-tion of certain cells without concomitant inflammation is
beneficial (4). The cells which had suffered apoptosis were
identified microscopically by the characteristic formation of condensed bodies of nuclear chromatin. Later it was recog-nized that a major hallmark of this death process was the fragmentationofthegenomicDNAintooligomersof180- to
200-bp multiples, corresponding to the unit length of DNA
coiled aroundahistonecomplex(3).Thisprocesswasshownto
be due to the activation ofa Ca2+-dependent endonuclease and to be ultimately responsible for apoptosis-mediated cell death (28).
There hasbeen accumulating evidence that apoptotic path-ways may account for the depletion of CD4+ T cells in
HIV-infected individuals. These pathways may be mediated J.VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.61.301.74.514.2] [image:7.612.326.544.74.272.2]APOPTOSIS BY gpl6O of HIV-1 397
D
o
t
2 3(DAYS)
FIG. 7. Cellgrowth in the presence of anti-Fas MAb or recombi-nantTNF-ot. U937-2 cells(A and C) and UE160 cells (B and D) were adjustedto105 cells in the RPMI1640 medium and cultured for up to 4daysin the presenceof CH-11 MAb (panelsAandB) or recombi-nant TNF-ot (panels C and D). The different symbols indicate the amountof the reagent addedtotheculture. Panels A and B:0,A, 0 ng/ml; O,A,0.5 ng/ml;C,
4,
1.0ng/ml;e,
A,2.0ng/ml; i,A,5.0 ng/ml; 0, A, 10.0ng/ml. Panels C and D: 0, A, 0 U/ml;OJ,
A, 0.1 U/ml; C, A, 1U/ml;e,
A,&
10U/ml;*, A,100U/ml.either directly by a virus infection or indirectly through the priming of uninfected cells to apoptosis when triggered by different agents (9). The participation of apoptosis in the cytopathic effect of CD4+ Tcells infected with HIV in vitro
was shown by Laurent-Crawford et al. (15). Their study demonstrated that the apoptotic events usually occurbefore thelysis of HIV-infected cells becomes dominant, suggesting
that the apoptosis is the direct cause of cell death by HIV
infection.Inaddition, inourpresentstudy with UE160, which isagpl60-expressing cell clone, the characteristic features of
apoptosis, suchasthefragmentationofDNAand the conden-sation of nuclearchromatin,werefoundas earlyas 8 h after induction, whereas the single-cell death detected by vital
staining with trypan blue dye was found later than 24 h. In
U937-2-derived cell clones such as UE160 and UE160-16, morethan 0.06 ,ugofgpl60per
106
cellsseems toberequiredfor the initiation of the apoptotic process and resultant cell death, because neither the UE160 cells without induction(0.06
,ug/106
cells) nor the UE160-16 cells after induction (0.03p.g/106
cells) showed any apparent cytopathiceffect.Sofarwe have been proposing that the intracellular
accu-mulation of thegpl6O-CD4complexdirectly participates in the
single-cell killing of the
gpl60-expressing
CD4+ cells. The complex formation between gpl60 andCD4 segregates CD4U937
UE160
MO
48 24 0 4 8 24 hr
bp
6033_
3100-FIG. 8. DNA fragmentation in cells treated with anti-Fas MAb. U937-2 and UE160 cells were treated with 1.0 ng of CH-11 per ml for the times indicated above each lane in the absence of inducer. Then small DNA wasextracted from 5 x 106cells of each cell sample and separatedon a 2% agarosegel as described in the legend to Fig. 4. Lane Mcontains a standard mixture ofHindlIl-digested X DNA and HaeIII-digested4X174DNA.
within the cells, resulting in the exhaustion of surface CD4 (14). By contrast, in the present study the disappearance of surface CD4 as a result of the segregation did not occur in the
gpl20-expressing cells, probably because of the secretable property of gpl20 (data not shown), suggesting that the character of retention or secretion kept by these envelope glycoproteins may determine their capability to provoke single-cellkilling. Although it remains unclear howthe intracellular
gpl60-CD4 complex causes the apoptosis of these cells, our analysis demonstrated thataprotein capable of nuclear loca-tion wasbarely transported into the nuclei after its microin-jection into such gpl60-expressing CD4+ cells at 4 h after induction whenDNA fragmentation has notyet occurred in the cells (12, 13). This indicates that the transportation of molecules between cytoplasm and nucleus through nuclear pores is blocked in those cells. It therefore presents the possibility that the occlusion of the nuclear pores activates the endonuclease which is responsible for apoptosis induction in the nucleus.
Using the CD4+ cell culture infected acutely with HIV in vitro, Terai et al. (27) also reported that apoptosis is the mechanism of cell death in the culture. Moreover, in their system the apoptosis is blocked by the addition ofanti-gpl20 neutralizing antibodyin spiteof sustained high levels of viral
replication. This suggests that the interaction of gpl20-CD4
TABLE 2. Quantitationof DNAfragmentation
% offragmentedDNAafter
Amtof CH-1( addedto incubation in the presence of Cell culture(nglml)
CH-1
1 for:Oh 4h 8h 24h
U937-2 0.5 NDa 3.0 4.7 14.5
U937-2 1.0 3.0 3.5 5.0 19.6
UE160 0.5 ND 12.8 13.4 48.0
UE160 1.0 12.4 14.2 15.5 62.1
a ND,notdone.
T-
r--x
0
100_
50
100 1 2 I
0 1 2 3
VOL.68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.79.267.71.377.2] [image:8.612.340.524.75.250.2] [image:8.612.314.557.623.711.2]398 LU ET AL.
FIG. 9. Expression level of surface Fas antigen. The expression level of surface Fas antigenonUE160 cells(A) and U937-2 cells (B) wasanalyzed byflowcytometrywith CH-11MAb. A100-,ul portion of
each cellsuspension (5 x 106 cellsperml) was addedto 100 [lof CH-11solution(20 ,ug/ml) and keptfor 60minonice. The mixturewas
washed with cold PBS, and the bound MAbwasrevealed by fluores-cein isothiocyanate-conjugated anti-mouse immunoglobulin M anti-body and analyzed byflowcytometry. The dotted lines representthe autofluorescence of the cells treated with fluorescein isothiocyanate-conjugated anti-mouse immunoglobulinMantibody only.
plays animportant role in the occurrence ofapoptosis in the
cell killing by HIV. In the present study, however, a
gpl20-expressing cell clone did not show any apparent cytopathic effectorapoptosis, although all the surface CD4son the cell wereoccupiedwithgpl2O.Thediscrepancyinapoptosis
induc-tion between theirstudyandours maybe duetothe difference inthemode ofcytopathic effect,such thatsyncytiumformation will be the dominant feature ofcytopathicity inHIV-infected cell culture while single-cell killing prevails in our
gpl60-expressingcell clones. Anotherpossibility explaining the
fail-ure to induce apoptosis inourgpl20-expressingcells may be
thatgp4laswellasgpl20isproducedinthe cells infected with
HIVwhereas only gpl20is expressedin ourcells.
ThesurfaceFasantigenisassociatedwith the TNFreceptor (29).TNFaugmentsboth thesynthesisofHIV-specificmRNA inHIV-infectedcells and thecytopathiceffect of these cells(8, 19). On the other hand, Kobayashi et al. (11) reported that
anti-Fas MAb does not augment HIV replication but
selec-tively kills chronically HIV-infected cells. Thus the cytotoxic
effect of TNFmaybe mediatedbythe Fasantigen, althoughits
physiological andpathologicalroles remain unclear.Although
inthepresentstudythe UE160 cellsalready expressedasmall
amount ofgpl60in the absence ofinducer, they exhibited no
apparentcytopathiceffect andgrew aswellastheU937-2cells
ifinducerwasnotadded. Thus non-CdCI2-treated UE160cells
may representthe CD4+ cellsinfected chronically with HIV. Aftertreatmentwithanti-FasMAb, likewise, such cells
exhib-itedamarkedcytopathiceffect,leadingtoextensive cell killing due to apoptosis as has been observed for chronically
HIV-infected cells. It therefore suggested that the remarkable
cytotoxiceffectby anti-Fas MAbfound in HIV-infected CD4+
cells is duetothe gpl60 expressed in these cells.
Theextentofsusceptibilitytothecytotoxic effect of anti-Fas
MAb is dependent on the amount of gpl60 because the expression of 0.03 ,ug of gpl60 per 106 cells rendered
100
D,
.50
10Q_
0 1 2 3 4 0 1 2 3(DAYS)
FIG. 10. Effect ofgpl60 expressionon thesusceptibility ofcellsto anti-Fas MAb.Theinhibitoryeffects of anti-Fas MAbonthegrowth of UE937-2cells(AandC)andUE160-16 cells(BandD)wereexamined in the absence(panels A andB)orpresence (panelsC and D)of 10 F.MCdCl2.The cells wereadjustedto105cells in 1 ml ofRPMI1640 medium on day 0 by the addition of various amounts of CH-11 in either the absence orthe presence of 10 F.M CdCl2and then were cultured for 4 days. On days3 and 4, the total number of cellswas counted with a Coultercounter. The different symbols indicate the amountof CH-1 1 addedtothe culture:0,0ng/ml;O,0.5ng/ml; C,1.0 ng/ml;
Q,
2.0 ng/ml;e,
5.0 ng/ml;*, 10.0ng/ml.UE160-16cells farmoresusceptibletoanti-FasMAbthanthe
UE160-16cells notexpressing gpl60.The lack of asignificant difference in the expression level of surface Fas antigen between UE160 and U937-2 cells suggested that putative
apoptosis-inducing machineries located downstream in the surfaceFas are affectedby gpl60,andsuchaneffectmakes the
cells highlysusceptible toanti-Fas MAb.
Inconclusion, gpl60 is cytotoxictoCD4+ cellsas aresult of
forming an intracellular gpl60-CD4 complex, as reported previously(14), and its death process is apoptosis.Evenwitha
minimal amount, not sufficient to inhibit cell growth, gpl60 alreadyexerts ahiddencytopathic effectontheCD4+ cells as can be observed by the presence of anti-Fas MAb-induced apoptosis. Such alatentcytotoxic effect exertedby gpl60can
also be detectedas a preapoptosis pattern by flow cytometry
analysiswith ethidium bromidestaining.
REFERENCES
1. Asjo,B.,I.Ivhed,M. Gidlund, S.Fuerstenberg,E. M.Fenyo,K. Nilsson, and H.Wigzell. 1987. Susceptibility to infectionby the human immunodeficiencyvirus (HIV)correlates with T4 expres-sion inaparental monocytoidcell line andits subclones.Virology 157:359-365.
2. Burton,K. 1956.Astudy of the conditions and mechanism of the diphenylaminereaction for thecolorimetric estimation of deoxyri-bonucleic acid. Biochem. J. 62:315-323.
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:9.612.122.238.73.259.2]APOPTOSIS BY gpl60 of HIV-1 399 3. Cohen, J. J. 1993. Apoptosis. Immunol. Today 14:126-130.
4. Duvall, E., and A. H. Wyllie. 1986. Death and the cell. Immunol. Today7:115-1 19.
5. Embretson, J., M. Zupancic, J. L. Ribas, A. Burke, P. Racz, K. Tenner-Racz, and A. T. Haase. 1993. Massive covert infection of helper T lymphocytes and macrophages by HIV during the incubation period of AIDS.Nature (London) 362:359-362. 6. Fahey, J. L., H. Prince, M.Weaver, J. Groopman, B. Visscher, K.
Schnartz, andM.Petels. 1984.Quantitative changes in T helper or T suppressor/cytotoxic lymphocyte subsets that distinguish ac-quired immunedeficiency syndrome from other immune subsets disorders. Am. J. Med. 76:95-100.
7. Fauci, A. S. 1988.The human immunodeficiency virus: infectivity andmechanisms ofpathogenesis.Science 239:617-622.
8. Folks,T.M.,K. A.Clouse, J. Justement,A.Rabson,E.Duh, J.H. Kehrl, and A. S. Fauci. 1989. Tumor necrosis factor a. induces expression of human immunodeficiency virus in a chronically infected T-cell clone. Proc. Natl. Acad. Sci. USA 86:2365-2368. 9. Gougeon,M.L., A. G. Laurent-Crawford,A.G.Hovanessian, and
L.Montagnier. 1993. Direct and indirect mechanisms mediating apoptosis duringHIVinfection: contribution to in vivo CD4Tcell depletion. Semin. Immunol. 5:187-194.
10. Kawamura, I., Y. Koga,N.Oh-hori, K. Onodera, G. Kimura, and K.Nomoto. 1989.Depletion of the surface CD4 molecule by the envelopeprotein of human immunodeficiency virus expressed in a human monocytoid cell line. J. Virol. 63:3748-3754.
11. Kobayashi, N., Y. Hamamoto, N.Yamamoto, A. Ishii, M. Yone-hara, and S. Yonehara. 1990. Anti-Fas monoclonal antibody is cytocidaltohumanimmunodeficiencyvirus-infected cells without augmenting viralreplication. Proc.NatI.Acad. Sci. USA 87:9620-9624.
12. Koga, Y.,M. Sasaki,K.Nakamura, G. Kimura,and K.Nomoto. 1990. Intracellular distribution of the envelope glycoprotein of humanimmunodeficiency virus and its role in the production of cytopathic effect in CD4+ and CD4- human cell lines. J. Virol. 64:4661-4671.
13. Koga,Y., M. Sasaki,H.Yoshida, M.Oh-tsu, G. Kimura, andK. Nomoto. 1991. Disturbance of nuclear transport of proteins in CD4+ cellsexpressing gpl60of humanimmunodeficiency virus. J. Virol.65:5609-5612.
14. Koga,Y., M.Sasaki, H. Yoshida, H. Wigzell, G. Kimura,and K. Nomoto. 1990. Cytopathic effect determined by the amount of CD4molecules in human cell linesexpressing envelope glycopro-tein ofHIV.J.Immunol. 144:94-102.
15. Laurent-Crawford, A. G., B.Krust, S. Muller, Y. Riviere,M.-A. Rey-Cuille, J.-M. Bechet,L.Montagnier,and A.G. Hovanessian. 1991. Thecytopathic effect of HIV is associated withapoptosis. Virology 185:829-839.
16. Lyons,A. B., K.Samuel, A. Sanderson,and A. H. Maddy. 1992. Simultaneous analysis of immunophenotype and apoptosis of murine thymocytes by single laser flow cytometry. Cytometry 13:809-821.
17. Matsushita, S.,M.Robert-Guroff, J. Rusche,A.Koito,T.Hattori, H.Hoshino, K.Javaherian, K. Takatsuki, and S. Putney. 1988. Characterization ofahumanimmunodeficiencyvirusneutralizing
monoclonal antibody andmapping of the neutralizing epitope. J. Virol.62:2107-2114.
18. Newell, M. K., L. J. Haughn, C. R. Maroun, and M. H. Julius. 1990.Death of mature T cells by separate ligation of CD4 and the TCR forantigen. Nature (London) 347:286-289.
19. Osborn, L., S. Kunkel, and G. J. Nabel. 1989. Tumor necrosis factorotandinterleukin 1 stimulate the human immunodeficiency virusenhancer by activation of the nuclear factor KB. Proc. Natl. Acad. Sci. USA86:2336-2340.
20. Pantaleo, G., C. Graziosi, J. F. Demarest, L. Butini, M. Montroni, C. H.Fox, J. M.Orenstein, D. P. Kotler, and A. S. Fauci. 1993. HIVinfection is active and progressive in lymphoid tissue during the clinically latent stage ofdisease. Nature (London) 362:355-358.
21. Potter, H., L. Weir, and P. Leder. 1984. Enhancer-dependent expression of human immunoglobulin genes introduced into mouse pre-B lymphocytes by electroporation. Proc. Nati. Acad. Sci. USA 81:7161-7165.
22. Ratner, L., W.Haseltine,R.Patarca,K.J.Livak,B.Starcich, S.F. Josephs, E. R. Doran, J. A. Rafalski, E. A. Whitehorn, K. Baumeister, L. Ivanoff, S. R.Petteway,Jr.,M. L.Pearson, J.A. Lautenberger, T. S. Papas, J. Ghrayeb, N. T. Chang, R. C. Gallo, and F. Wong-Staal. 1985. Complete nucleotide sequence of the AIDSvirus, HTLV-III. Nature (London) 313:277-284.
23. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning:alaboratorymanual,2nd ed., p. 15.74-15.79. ColdSpring Harbor Laboratory Press, ColdSpring Harbor, N.Y.
24. Shaw, G. M.,B. H.Hahn, S.K.Arya, J.E.Groupman,R.C.Gallo, and F. Wong-Staal. 1984. Molecular characterization of human T-cell leukemia (lymphotropic) virus type III in the acquired immunedeficiency syndrome. Science 226:1165-1171.
25. Somasundaran, M., and H. L.Robinson. 1987. Amajor mecha-nism of human immunodeficiencyvirus-induced cell killing does notinvolve cell fusion. J. Virol. 61:3114-3119.
26. Stevenson, M., C. Meier, A. M. Mann, N. Chapman, and A. Wasiak.1988.Envelopeglycoproteinof HIV induces interference andcytolysis resistanceinCD4+ cells: mechanism forpersistence inAIDS. Cell 53:483-496.
27. Terai, C.,R.S. Kornbluth, C.D.Pauza,D. D.Richman,and D. A. Carson. 1991.Apoptosisas amechanism of cell death in cultured T lymphoblasts acutely infected with HIV-1. J. Clin. Invest. 87:1710-1715.
28. Wyllie,A. H.,R. G. Morris,A. L. Smith, and D. Dunlop. 1991. Chromatin cleavage in apoptosis: association with condensed chromatin morphologyand dependenceon macromolecular syn-thesis. J. Pathol. 142:67-77.
29. Yonehara, S., A. Ishii, and M. Yonehare. 1989. A cell-killing monoclonal antibody (Anti-Fas) to a cell surface antigen co-downregulated with the receptor oftumornecrosis factor.J.Exp. Med.169:1747-1756.
30. Zagury, D., J. Bernard, R. Leonard, R. Cheynier, M.Feldman, P.S.Sarin,and R.C. Gallo. 1986. Long-term cultures of HTLV-III-infectedTcells:amodel ofcytopathologyof T-celldepletionin AIDS.Science 231:850-853.
VOL.68, 1994