Vol. 58, No. 3 JOURNALOFVIROLOGY, June 1986,p.805-816
0022-538X/86/060805-12$02.00/0
Copyright C) 1986, American Society for Microbiology
Mapping of Phosphorylation Sites in Polyomavirus
Large T
Antigen
MARTIN HASSAUER, KARL HEINZSCHEIDTMANN,* ANDGERNOT WALTERt
Institutfur Immunbiologie der Universitat Freiburg, Stefan-Meier-Str. 8, D-7800 Freiburg, Federal Republic of Germany Received 28 October 1985/Accepted 24 February 1986
The phosphorylation sites of polyomavirus large T antigen from infected or transformed cells were investigated. Tryptic digestion of large T antigen from infected, 32P1-labeled cells revealed seven
major
phosphopeptides. Five of these were phosphorylated only at serine residues, and two were phosphorylated at serine and threonine residues. The overall ratio of phosphoserine to phosphothreonine was 6:1. The transformed cell line B4 expressed two polyomavirus-specific phosphoproteins: large T antigen, which was only weakly phosphorylated, and a truncated form of large T antigen of 34,000 molecular weight which washeavily phosphorylated. Both showed phosphorylation patternssimilarto thatof large Tantigen from infectedcells. Peptide analyses of large T antigens encoded by the deletion mutants d18 and d123 or ofspecific fragmentsof wild-type large T antigen indicated that the phosphorylation sites are located in an amino-terminal region upstream of residue 194. The amino acid composition of the phosphopeptides as revealed by differential labeling with various amino acids indicated that several phosphopeptides contain overlapping sequences and that all phosphorylation sites are located in four tryptic peptides derived from a region between Met7l and Argl91.Two of thepotential phosphorylation sites were identified as Ser8l and Thrl87. Thepossible role of thismodification oflarge T antigen is discussed.
The early region of the polyomavirus genome encodes threeproteins,thesmall(sT), medium-sized (mT),andlarge T(LT)antigens, whichshare theamino-terminal 79 residues but differ in their
carboxy-terminal regions.
Among these proteins,LTplaysamajor regulatory role.Inlytic infection, it is involved in initiation ofreplication
andregulation
of transcription ofthe viral genome(forareview,seereference 16). These functions are probably mediated by specific interactions with regulatorysequences onthe viral chromo-some(9,31). Other biochemicalproperties of LT,such as its ATPase activity (13) ornucleotide binding (7), mayalso be relatedtoDNAreplication (6). Inmalignant transformation, bothLTand mT arerequired. LTalonemayconfer immor-talityuponprimary cellsand may reduce therequirement of transformed cells forgrowth factors (33, 35);mTisrequired
toobtainfull transformation (33, 34, 47).
Somefunctions ofLT canbeascribedtodistinct structural domains (28, 33, 35). Anadditional elementinregulation of its various activities may be phosphorylation (36). In the case of simian virus 40 (SV40), phosphorylation of LT appears to affect its DNA-binding properties (26, 38) and, perhaps, oligomerization and interaction with the cellular transformation-specific
p53
protein (4, 11, 15). SV40 LT exists inatleast twodistinctstates ofphosphorylation that may be characteristic for different functional states(38).Tostudy phosphorylation of polyomavirusLTis interest-ing from two points of view. First, the close homology between SV40 and polyomavirus suggests that similar cor-relations between phosphorylation and other biochemical propertiesmayexist forpolyomavirusLT.Second, fromthe
evolutionary standpoint,
it isinteresting
to see whether specific phosphorylation sites are conserved between these two proteins. This, in turn, would be an indication of the biologicalrelevanceofphosphorylationin thesesystems.In this study,we present data on the complex phosphor-ylation pattern of LT from polyomavirus-infected and
* Correspondingauthor.
tPresentaddress:DepartmentofPathology,School ofMedicine, University ofCalifornia, SanDiego, LaJolla, CA92093.
-transformed cells. Tryptic fingerprinting revealed seven major phosphopeptides, five of whichwere phosphorylated atserine residues andtwoof which werephosphorylatedat both serine and threonine residues. These phosphopeptides were localized onthepolypeptide chain by analyzing either fragments of LT generated by partial proteolysis or LTs encoded by thedeletion mutantsd18 and d123. In addition, the amino acid
compositions
of thephosphopeptides
were determinedby differential labeling with various amino acids. The dataindicatethatallphosphorylation sitesareclustered infourtrypticpeptides
derived fromaregion betweenMet7l andArgl91.MATERIALS AND METHODS
Cells and virus strains.3T6mousecells
(originally
obtained from W. Gibson, San Diego, Calif.) and the polyomavirus-transformed rat cell line B4 (1) were grown in Dulbecco modified Eagle minimal essential medium containing 10% calf serum.Polyomavirus wild-typeA2(originally obtained from T. Friedman) and deletion mutants d18 and d123 (kindly provided by B.Griffin [18])werepropagated in 3T6cellsas describedpreviously
(17).Infection and radiolabeling of cells. For experiments, subconfluent 3T6cells growingon petri dishes (diameter,9 cm) wereinfected with
wild-type
polyomavirus ordeletion mutants d18ord123 at amultiplicity of infection of about 5 PFU per cell and afterwards kept in Dulbecco modified Eagle minimal essential mediumcontaining 5% calfserum. Radiolabeling was usually carried out from 24 to 27 hpostinfection.
B4 cells were labeled under subconfluent conditions. '4C-labeled amino acidswereapplied
at125,Ci,
[35S]methionine
or[35S]cysteine
wasappliedat250,Ci,and32p;
(carrierfree) wasappliedat 1 mCiperplateinmedium lackingthe corresponding amino acid orphosphate. Long-termlabelingwascarried out with 250,uCiof32p,
perplatein complete medium from 10 to 27 hpostinfection.
The radiochemicals were obtained from Amersham Buchler, Braunschweig, FederalRepublic of Germany.Preparation of cell extracts and purification of LT. Cell extracts werepreparedasdescribed
previously
(39);briefly,
805
on November 10, 2019 by guest
http://jvi.asm.org/
m
a b
c d
LT- - 92 K
rnT-
VP1-_S -42K
UM - 34 K
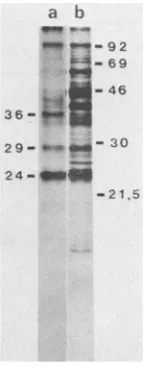
sT--FIG. 1. T antigens from infected and transformed cells. Poly-omavirus-infected 3T6 mouse cells (lanes a and b) or the trans-formedratcelllineB4(lanes c and d) was labeled with [35S]meth-ionine (lanes a and c) or32p, (lanes b and d). Cell extracts were prepared, and the T antigens were isolated by immunoprecipitation and SDS-PAGE and visualized by fluorography as described in Materials and Methods. The molecular weight standards (lane m)
were 14C-labeled phosphorylase b (92K), bovine serum albumin (69K), ovalbumin (46K), carbonic anhydrase (30K), and lysozyme (15K). LT, mT, and sT seenin lanes a and c are labeled.
after labeling, the cells were washed with cold phosphate-bufferedsalineand lysed withisotonicphosphate buffer (10 mM NaPO4, 10 mM Tris hydrochloride [pH 9], 140 mM NaCl, 3 mM MgCl2, 1 mM dithiothreitol) containing 0.5% Nonidet P-40 and 10 ,ug of aprotinin (Boehringer GmbH,
Mannheim,
FederalRepublic of Germany) perml; pH9 was usedinstead ofpH 8because pH9providedhigher yields of LT. Thenucleiwerepelleted by low-speedcentrifugationat 800 x gfor3min,andthe cellextracts werefurthercleared by centrifugation at 13,000 x g for 10 min. LT was then isolated by immunoprecipitation with serum from tumor-bearing rats (RaT serum) (43) and with Staphylococcus aureus as describedpreviously
(25); it was furtherpurified by electrophoresis through 12.5% sodium dodecyl sulfate-polyacrylamide gels (SDS-PAGE) (23).Generation ofspecific fragments of LT by partial tryptic digestion. After immunoprecipitation, the immune com-plexeswere washedand digestedwith 20
jIg
oftrypsin per ml (treated with L-1-tosylamide-2-phenylmethyl chloro-methyl ketone [TPCK])at0°C
for 20min,bythemethodof Schwyzeretal. (42).Thedigestion productswereseparated by electrophoresison 15% SDS-polyacrylamide gels.Two-dimensionalpeptide analysis. LT orfragmentsthereof werelocalizedonthe dried gels by autoradiography and then excisedandeluted;they wereprecipitatedwith20% trichlo-roacetic acid, with 10 ,ig ofbovine y-globulin per ml as
carrier,
and oxidized with 3% performic acid as describedpreviously
(37, 39).Theprotein samplesweredigested
either8
j
i
11
n
4
1
7
9 7a
I13 9
la:
..1..
b
7 3
Ix
x
6 5 4
c
7 7a
6
I..
5 2,A1 g
1b 3
3a 4
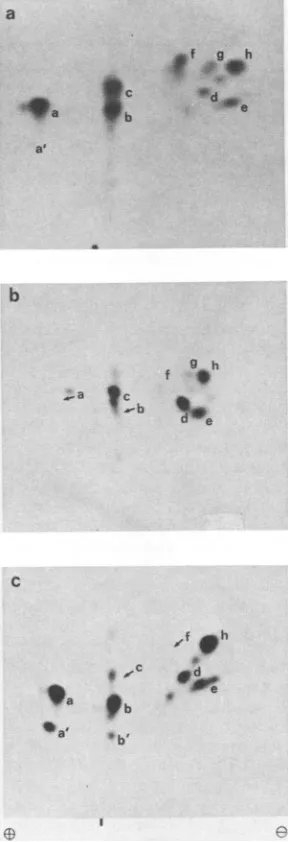
FIG. 2. Two-dimensionalseparationoftrypticphosphopeptides ofTantigens from infectedortransformed cells. Cellswerelabeled with
32Pi
andthe Tantigenswereisolatedbyimmunoprecipitation andSDS-PAGE andprocessed forpeptidemappingasdescribed inthe text. The phosphopeptides were separated onTLC plates by electrophoresisatpH1.9 and 1 kVfor40min(cathodeontheright)
andascendingchromatography inisobutyric acid buffer (37) for5h. Thephosphopeptideswerethen visualizedbyautoradiographywith intensifyingscreens. The vertical dashindicates theorigin. (a)LT
frominfected cells; (b)92K protein (= LT) from transformedB4
cells; (c) 34K proteinfromtransformed B4cells(see thelegendto
Fig. 1).Thepeptide designatedxin(b)wasunique forLTfromB4
cells;it was notfurtheranalyzed.Peptides lband3aseeninpanel
c probablyarise from incomplete digestion; theywere repeatedly observedin LTpreparations from varioussources.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.94.269.77.333.2] [image:2.612.368.511.98.578.2]PHOSPHORYLATION SITES OF POLYOMAVIRUS LARGE T ANTIGEN
with trypsinalone orsequentially with trypsin and pronase; digestionwas performed with 20 ,ug of enzyme per ml in 50 mM NH4HCO3, pH 7.8, at 37°C for 10 h (37). The digestion products were separated oncellulose thin-layer chromatog-raphy (TLC) plates byelectrophoresis and ascending chro-matography (14). Electrophoresis was usually carried out at pH 1.9inbuffer containing(vol/vol) 6%formic acid, 1.25% aceticacid, and 0.25% pyridine; insome cases,pH4.7buffer (5% butanol, 2.5% acetic acid, 2.5% pyridine [vol/vol]) was usedinstead. Ascending chromatography wasperformed in isobutyric acid buffer (isobutyric pyridine-acetic acid-butanol-water [65:5:3:2:29,vol/vol])(37).35S-or
14C-labeled
peptides were visualized by fluorography (2). 32P-labeled peptideswere visualized by autoradiography with intensify-ing screens.
RESULTS
Separation ofthephosphopeptides of LT. Toinvestigate the phosphorylation sites of LT, cells wereinfected with wild-typepolyomavirus A2and labeled with32p;. Thecellswere lysed, and theextract wasimmunoprecipitated and analyzed by SDS-PAGE as described in Materials and Methods. LT was the major phosphoprotein obtained (Fig. 1, lane b). Longer exposures revealed additional protein bands which were notfurtheranalyzed. Forcomparison,an immunopre-cipitate of
[35S]methionine-labeled
cellextractwasanalyzed (Fig. 1, lane a).Inthiscase,LT, mT, sT, and the viralcapsid protein VP1 wereprecipitated. Inaddition, several cellular proteins, ranging in size between70,000 and 30,000 molec-ularweight,wereobserved, withthe mostprominentband at 42K.For subsequent phosphopeptide analysis, 32P-labeled LT was eluted from the gel anddigested with trypsin, and the resulting
phosphopeptides
were subjected to two-dimen-sionalpeptide mappingasdescribed in Materials andMeth-P-Ser- P-Thr
I 0
la,b *
2
0
3,3a
5
0p
6 *
7 9
4 ,
."""mq
e
e
FIG. 4. Phospho-amino acid analysis of individual phos-phopeptides(listed tothe left of thefigure). The tryptic phospho-peptides shown in Fig. 2a were eluted from the TLC plates, hydrolyzed, and analyzed as described in the legend to Fig. 3, except that chromatography was omitted. The cathode is on the right. 0,Origin.
m1P
IP-
Set-0..
P-Tyr
[image:3.612.348.517.71.356.2]P-ThI 0
FIG. 3. Two-dimensional analysis of phospho-amino acids of LT. 32P-labeled LT was purified andhydrolyzed in 5.6N HCl at 110°C for 2 h in sealed glass tubes. Unlabeled phosphoserine phosphothreonine,orphosphotyrosinewasadded,andthe mixture
was separated by electrophoresis at pH 1.9 and 1.3 kV for 1 h,
followed by ascending chromatography in isobutyric acid-0.5 M
NH40H (5:3, vol/vol) (10). The markers were visualized with
ninhydrin,andthe labeledphospho-aminoacidswerevisualizedby autoradiography.The dottedspotindicates thepositionof unlabeled
phosphotyrosine.The cathode isontheright. 0,Origin.
ods. Seven
major
phosphopeptides
and two minor phosphopeptides wereresolved and designated 1through 7 and laand7a,respectively (Fig.
2a).Peptides
la, 3, and5 sometimessplitted
intodoublets,
probably because of in-completedigestion (Fig. 2c). Phosphopeptide
7a was notalways detected.All
peptides
hadanetpositive charge
atpH
1.9,asthey migrated to the cathode duringelectrophoresis.
With theexception
ofphosphopeptide
7, theyappeared
tobe very polar, since they did not migrate in conventionalchromatography
buffers(datanotshown).Phospho-amino acid analysis. The
phosphorylated
amino acidswere identifiedby
acidhydrolysis
of total 32P-labeled LT. Thehydrolysis products
wereseparated
onTLCplatesby
electrophoresis
andchromatography
toallowseparation
ofthreoninephosphate
andtyrosinephosphate.
Phosphoryla-tion occurredexclusively at serine and threonine residues, thusconfirming previous
results(Fig. 3) (36).
The ratio ofphosphoserine
to phosphothreonine was about 6:1, both after short- (3h)
or long-term (20 h)labeling.
Thus, weexpected about seven (or
multiples thereof)
differentphos-phorylation
sites.Toidentify the phosphorylated residues in the individual phosphopeptides, the peptides were separated by electro-phoresis and chromatography on TLC plates, eluted, and hydrolyzed.The
phosphoamino
acidswerethenanalyzedby
electrophoresis (Fig.
4).Phosphopeptides
1, la,lb,
2,3,3a, 5, and 7contained only phosphoserine. Phosphopeptides 4 and6contained bothphosphoserine
andphosphothreonine
inapproximate ratiosof 0.55 and1.5,respectively.Thesmall amount of
phosphothreonine
inpeptide
5 wasprobably
VOL.58, 1986 807
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.93.257.446.637.2]a
7
1
1t
.1s1
Oat 2'
75a
9~~~
dl
237
2 9
4 5
[image:4.612.157.465.75.241.2],
13
49
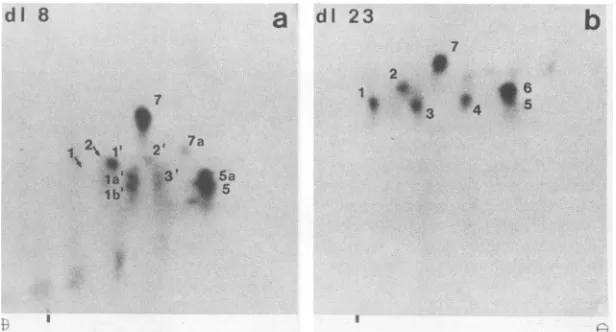
FIG. 5. Phosphopeptide analysis ofLTsencoded by d18(a)andd123 (b). Cells wereinfectedwith thedeletionmutantsd18ord123 and
labeled with 32p;. The LTswereisolated and processed for peptide mappingasdescribed for wild-typeLT inthe legendtoFig. 2. Thearrows
indicate thepositions of peptides1 and 2 ofwild-typeLTasassessedby analyzingamixture ofphosphopeptides from wild-typeandd18.The
cathode is onthe right.Theverticaldashes indicate theorigin.
causedby contamination from peptide 6 because of thepoor separation of thetwopeptides (Fig. 2) (see Discussion).
Phosphorylation patterns of T antigens in polyomavirus-transformed cells. To investigate whether phosphorylation of lToccursatthesamesites in infected and transformed cells, the polyomavirus-transformed cell line B4 (1) was labeled with
32Pi
and analyzed by immunoprecipitation and SDS-PAGE as described above. In this case, the major phosphorylated polypeptides appeared as a34,000-molecular-weight doublet (34K doublet) (Fig. 1, lane d). Two minor bands were seenat92K(the size of LT) and at42K. When B4 cells were labeled with [35S]methionine (Fig. 1,
lane c), the 34K doublet occurred only at weak bands, whereas the 92K and 42Kproteins werereadily visible. The 42Kprotein presumably represented the same cellular pro-teinasthatseenininfected cells(compare Fig. 1 lanesaand
c). It was not investigated further. The 92K and 34K phosphorproteins were isolated and subjected to phospho-peptide analysis as described above. The 92K protein gave rise to the same phosphopeptides as did LT from infected
cells, though with different intensities (compare Fig. 2a and b), thus it represented LT. An additional phosphopeptide, designated "x", was reproducibly observed; it was not
investigated further. The 34K protein (Fig. 2c) showed the
same phosphorylationpatternasdid LTfrom infected cells. This latter resultindicated that the 34K polypeptide repre-sentedatruncatedform ofLTand thatthephosphorylation
sitesareclustered inasmall region of LT.
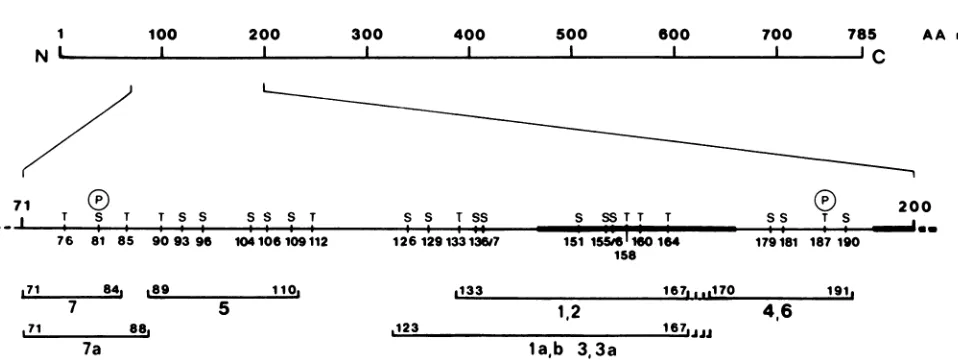
Mapping of phosphopeptides ofLT. To localize the phos-phorylation sites of LT within its primary structure, we
performed three different analyses: (i) analysis of LTs of various deletion mutants; (ii) phosphopeptide analysis of specific fragments of LT; and (iii) determination of theamino
acid composition of the phosphopeptides by radiolabeling with various amino acids.
Phosphopeptide analysis of LTs encoded by deletion
mu-tantsd18andd123. 3T6 cellswere infected with the deletion
mutantsd18 andd123,carryingdeletions from amino acid 145
to174orfromamino acid 195to228, respectively (45). The cells were labeled with 32p;, and LT was isolated and
analyzed by tryptic peptide mapping as described above.
The resultsareshown inFig. 5. LT encoded by d123 revealed a phosphorylation pattern indistinguishable from that of
wild-type LT (compare Fig. 5 with Fig. 2a). In contrast, LT of d18 lackedpeptide 4. Peptide 6, whichwasnormallymore heavily labeled than peptide 5 appeared to be strongly reduced. In further analyses, in which the amino acid composition of the phosphopeptides was determined, it became evident thatpeptide 5 always appearedas adoublet (5 and 5a) and that peptide 6wassuperimposedupon Sa(see Fig. 9b and d). Thus,onthe phosphopeptidemapof d18, the residual material at the position of peptide 6 represented peptide Sa, whereas peptide 6 was absent. Additional changes appeared in peptides 1 to 3, in that these peptides migrated farther to the cathode than did thecorresponding phosphopeptides of wild type; therefore, they were desig-nated 1', 2', and 3', respectively. The positions to which peptides 1 and 2 of wild-type LT migratedonpeptide maps of mixed samples of d18 and wild-type LTareindicated by
arrows (Fig. 5). These findings suggested that some phos-phorylation sitesarelocated within the sequencedeleted in d18 or that the deletion covers part ofsome of the tryptic phosphopeptides and alters their properties.
Phosphopeptide analysis of fragments of LT generated by partial proteolysis. When SV40 LT immune complexesare treated with trypsin under mild conditions, LT is cleaved only at preferential sites without disruption of the immune complex. The fragments generated can be isolated and further analyzed (37, 42).
Whtn
polyomavirus LT was treated in thesameway,36K, 29K, and 24K phosphorylated fragmentswereobserved(Fig. 6, lane a). No fragmentswere released from the immune complex during the treatment.Partial proteolysis of [35S]methionine-labeled material re-vealed additional fragments (Fig. 6, lane b), some of which
wereprobably derived from the unphosphorylated region of
LT; others might have been generated from cellular proteins. The 36K, 29K, and 24K phosphorylated fragments were isolated, exhaustively digested with trypsin, and submitted
to two-dimensional peptide mapping. The 36K fragment showed the same phosphorylation pattern as did complete LT(compare Fig. 2a and 7a) and the 29K fragment (datanot
shown). This was in agreement with the results obtained with the 34Kfragment from the B4 cells. The 24K fragment lacked phosphopeptides 4 and 6, as did LT ofd18; but in
contrast to the d18, phosphopeptides 1 to 3 were present (Fig. 7). We concluded that the phosphorylated 36K, 29K,
dl
8b
on November 10, 2019 by guest
http://jvi.asm.org/
PHOSPHORYLATION SITES OF POLYOMAVIRUS LARGE T ANTIGEN
a b
_0 w f
-92 -69 36 46
36-P$
29- _ so
24
--2 .5
FIG. 6. Analysis by SDS-PAGE of fragments ofLTgeneratedby partial tryptic digestion. LT was immunoprecipitated from 32P-labeled (lane a)or[35S]methionine-labeled(lane b) cellextracts and
digestedwithtrypsinunder mildconditions(for details, see
Mate-rials andMethods). The resulting fragmentswereseparatedon15% SDS-polyacrylamidegels and visualized by autoradiography.
Num-bersontheright(inthousands)refertomolecularweightstandards,
asinFig. 1; 21.5= soybean trypsin inhibitor.
and24Kfragmentswerederivedfrom overlapping regions of LT and that the larger ones cover at least part of the sequence deleted in d18.
Since d123 did not affect any phosphorylation sites, the 36K(and 29K)fragment(s)mustbelocated eitherupstream orbothupstream and downstreamofthedeletionind123,in either case covering the sequence deleted in d18. The two possible locationsareshownin Fig.8. It was notpossibleto directly distinguish these alternatives
by
partial digestion, because themutated LTs were moresensitivetotrypsinand yielded phosphorylated fragments smaller than 7,000 (data notshown).Instead,
wetookadvantage of the factthat the threeTantigens share their amino-terminal 79 residues and hence three tryptic methionine peptides. If the fragments were derived from the amino terminus ofLT they should36
K
a
N
785
-J C
dl8d123
a) _____ 24K
a) ___________ 36 K
b) 224K
36 K
FIG. 8. Possiblealignments ofthe 36Kand24Kfragmentsonthe
LTmolecule. Theupperlinerepresents LTwith the deletions of d18
anddl23 indicated bybars. The lines below indicate the possible
locations ofthephosphorylatedfragments.
have contained themethionine peptidescommon tomTand sT. The methioninepeptides of the 36K and 29K fragments ofLTwerecompared withthose of complete mTby peptide mapping of [35S]methionine-labeled T antigens. To avoid a possible contamination of the LTfragments with fragments of mT which might also be generated during partial diges-tion, mT was first isolated from the cell extract with a monoclonal antibody specific for mT (19); LT was then immunoprecipitated with RaT serum and partially digested asdescribed above. Upon comparative peptideanalysis, the LT fragments did show three methionine peptides in com-monwith mTand one methionine peptide derived from the splice junction of LT which comigrated withphosphopeptide 7 (data not shown). Thus, the fragments containing the phosphorylation sites were derived from the amino terminus of LT(possibility (a)inFig. 8).
Amino acid composition of the phosphopeptides. To char-acterize the phosphopeptides more precisely, their amino acid compositions were elucidated by differential labeling with various amino acids as described previously (37, 39). A set ofradiolabeled amino acids was chosen thatwould allow clear distinction oftryptic peptides with potential serineor threonine phosphorylation sites, as deduced from the pre-dicted amino acid sequence of LT (12, 46). Infected cells were individually labeled with [35S]methionine or [35S]cysteineorwithvarious 14C-labeledamino acids (Table 1)orwith 32p;. Intact LTwasisolated by immunoprecipita-tion and SDS-PAGE and digested with trypsin. Peptide mappingwascarried out in two ways, either withtheamino
24
K
7
I
la
lb
2
b
7a
5a .i5 3
3a
I Ie
FIG. 7. Phosphopeptide analysis of fragments ofLTgeneratedbypartial tryptic digestion. 32P-labeledLTwasimmunoprecipitated and
partiallydigestedasdescribed inthelegendtoFig.6. 36K, 29K, and 24Kfragmentswereisolatedandanalyzed furtherby peptidemapping
asdescribed inthelegendtoFig.2.Thepatternofthe29Kfragmentwasidenticaltothat of the 36Kfragment.The cathode isontheright.
Theverticaldashesindicatetheorigin.
VOL. 58,1986 809
2
t
. 7a
il..-.Alk-t.
64 5
on November 10, 2019 by guest
http://jvi.asm.org/
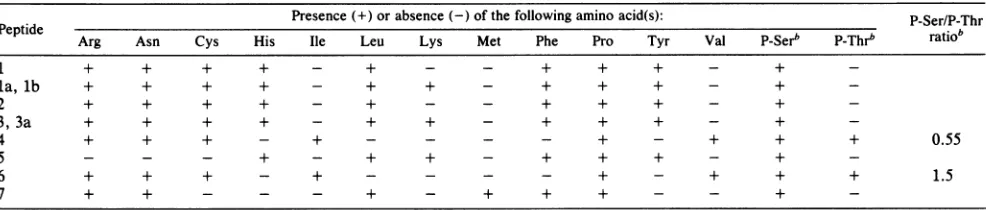
[image:5.612.146.219.74.259.2] [image:5.612.319.560.76.152.2] [image:5.612.154.464.521.686.2]TABLE 1. Amino acidcompositionofthe phosphopeptides ofLT'
Presence(+) orabsence (-) of thefollowing amino acid(s): P-Ser/P-Thr
Peptide Arg Asn Cys His Ile Leu Lys Met Phe Pro Tyr Val P-Serb P-Thrb ratiob
1 + + + + - + - - + + + - +
-la,lb + + + + - + + - + + + - +
2 + + + + - + - - + + + - +
-3, 3a + + + + - + + - + + + - +
4 + + + - + - - - - + - + + + 0.55
5 - - - + - + + - + + + - +
-6 + + + - + - - - - + - + + + 1.5
7 + + - - - + - + + + - - +
-a Theamino acid composition was elucidated bytwo-dimensionalpeptide analyses of mixtures of32P-labeledand
"S-
or14C-labeledaminoacid-labeledLT asdescribedin the text. bp_,Phospho-.
acid-labeled samples alone or with a mixture of the amino acid-labeledand32P-labeled peptides.The mixture served to allow clear identification of peptides withbothkindsof label. On the mixed fingerprints, the 32P-labeled peptides were distinguished by shielding the 35S or 14C radiation with aluminum foil; the amino acid-labeled peptides alone were visualized by fluorography after decay of32P. Samples of arginine, histidine, leucine,andlysineare shownin Fig.9. It is evident that arginine (Fig. 9a and b) was present in peptides 1 to 3, 4, 6, and 7 but absent from 5 (and 5a); histidine (Fig.9c andd)was presentin peptides1to 3, 5, and 5a but absent from the others; leucine (Fig. 9e and f) was present inpeptides 1 to 3, 5, 5a, and 7; andlysine (Fig. 9g andh) was presentinpeptides la, lb,3, 3a, 5, and 5a, but was absent from peptides 1, 2, 4, 6, and 7. The amino acid composition ofthe individual phosphopeptides, as derived from these experiments, is shownin Table 1. Accordingto their compositions, the phosphopeptides were aligned with predicted tryptic peptides ofLT as shown in Fig. 10. The
peptides
aredescribed in theorder in which they appearin thepolypeptide chain. For easeof discussion, the sequenceof
theamino-terminal region ofLTis shown inFig.
11. The overallcomposition of phosphopeptide7 was compat-ible with a peptide, extending from Met7l to Arg84 and spanning the splice junction ofLT. Since itwas phosphory-lated onlyat serine and theidentified peptide contains only one serine residue at position 81,Ser8l
must be the phosphorylated residue.Phosphopeptides 5 and 6 separated only poorly during peptide mapping. Moreover, peptide 5 appeared as a dou-blet, 5 and 5a, with peptide 6 partially superimposed upon
peptide
5a.
Butthesepeptides could be clearlydistinguished
onthebasis of their amino acid
compositions. Peptide
5(and
5a)
containedhistidine, leucine, lysine (Fig. 9, panelsc toh), phenylalanine, and tyrosine (data not shown), which were absent in peptide 6. On the otherhand, peptide 5 (and5a)
containedno arginine (Fig. 9aandb), asparagine, cysteine, isoleucine, or valine (data not shown), whereas peptide 6 did.Thecomposition of phosphopeptide5wasonly compat-ible with a peptideranging
from Gly89 to LysllO and containing five serine residues and one threonine residue. Thephosphorylated residue(s) could not beidentified.Phosphopeptides 1through 3 showedidenticalamino acid compositions except for lysine, which was presentonly in peptidesla,lb,3, and 3a (compareFig. 9 aand bwith g and h). Thus, these peptides were related to each other. The amino acid composition of peptides 1 and 2 pointed to a peptide extending from Thrl33 to oneofthe arginine resi-duesatpositions 167 to 169. The presence oflysine (andthe
absence of isoleucine and valine) in peptides la and 3 indicates that these peptides overlap with peptides 1 and 2 and are extended upstreamof Thrl33 to Trpl23 orLys122. Peptides 1 and 2contain fiveand peptides la and 3 contain seven serine residues. Again, a precise localization of the phosphorylation site(s) in peptides1 to 3 was notachieved. Phosphopeptides4and 6alsoshowed identicalamino acid compositions, indicating that they contained overlapping sequences. Their amino acid compositions wereonly com-patible with apeptide extending from Asn170to Argl86or Argl9l. Since both peptides werephosphorylated atserine and threonine and Thr187 is the only threonine residue in this region, we conclude that trypsin did not cleave at Argl86 and that both peptides extend to Argl9l. Hence, Thr187 is one of the phosphorylated residues. The phosphorylated serine residuewas notidentified.
LTofd18 showed an altered phosphorylation pattern in that phosphopeptides 4 and 6 were absent and peptides 1 through 3 were altered (Fig. 5). The deletion in d18 (from residue 145 to 174) covers the carboxy-terminal part of peptides1 and2, including threeoutof fiveserine residues, and the amino-terminal portion of peptides 4 and 6, but excluding the potential phosphorylation sites in peptides 4 and6 (see scheme in Fig. 11). Consequently, tryptic diges-tion of the mutated LT would generate altered peptides composed oftheamino-terminalpartofpeptides 1and2and the carboxy-terminal partofpeptides 4 and 6. These
com-posite
peptideswerepresumably represented by peptides 1' through 3' in Fig. 5. To investigate which of the sites remaining inthecomposite peptideswerephosphorylated ind18,
the LTs ofwild-type and ofd18 were digested sequen-tially with trypsin and pronase with the assumption thatdigestion
withpronasewouldgenerate smallpeptides which would not be affected by the deletion. The relationship between thetryptic
andthepronasepeptides
waselucidated by digestingindividualtryptic peptideswith pronase andby subsequent two-dimensional peptide mapping. It should be noted that all tryptic peptides gave rise to two or more, probably overlapping,pronasepeptidesand that theratiosof thedigestion productsvaried to some extent. The results are summarized in Table 2. Pronase digestion of wild-type LT gave rise to eight majorand several minorpeptides desig-nated "a" to "h" (Fig. 12a). In LT ofd18,
the pronase peptides aand b were missing. These are derivedfrom the tryptic peptides1 to3(Table 2).On the otherhand, peptides cthrough h, whicharederived from thetryptic peptides4to 7, were present, except for peptide f, which was hardly detectable (Fig. 12b). As control, a pronase digest of the amino-terminal 24K fragment of wild-type LT which, likeon November 10, 2019 by guest
http://jvi.asm.org/
a
7
7a d
As
4!&
II*, q
C
1 2
'g :4
IV"3a
e
p
* ,
_~
9 *|~~~7a a
9
7 e
7a
12 4 4 5a
1a,b 3,3a
d
7 v
2 ,7a
lab3,3a tI
4
f
U
._
*1 2
. 'I 7
7
7.
2 4
l.ob3,3a w
7a
9
. *
* *
e01
Ae6
S.
Ia,b33a * ew
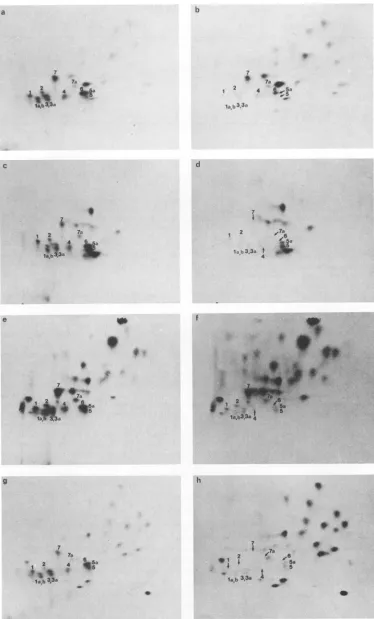
FIG. 9. Peptidemapsof LTlabeled with 32p_orvarious '4C-labeledamino acids. Polyomavirus-infectedcellswereindividually labeled
with32p;or['4C]arginine (a and b), histidine (cand d), leucine(eand f), orlysine (g and h), LTwasisolated by immunoprecipitation and
SDS-PAGE anddigested with trypsin; each amino acid-labeled samplewasmixed witha32P-labeledsample and subjectedtopeptidemapping asdescribed in thetext.Thepeptideswerevisualized by fluorography(2)before(a,c,e,g)andafter(b, d,f,and h) decay of the32Pradiation.
Only thephosphopeptidesaremarked with the respective numbers; arrowsindicate those phosphopeptides thatare nolongervisible after
decay of the32Pand hence donotcontain the respective amino acid. The cathode isonthe right of each panel. The vertical dashes indicate
theorigin.
811
it
p
h
I
b
I
a
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.121.495.51.670.2]I N L
100 200 300 400 500 600 700 785
I--
c
AA
no
71 (
I T S T T S S S S S T S S T SS S SST T T
I
I-I I: :,. e
76 81 85 90 93 96 104 106 109112 126 1291331367 151 15516 160 164 158
171 84, La9 7
171 88,
7a
110.
5
0133 167L,,1i70
1,2
.123 167,
191.
4,6
[image:8.612.68.547.59.239.2]la,b 3,3a
FIG. 10. Alignment of phosphopeptides of LT. The upper line represents the polypeptide chain with the region containing the
phosphorylation sitesshownenlarged in thesecondline.The positions of allserine and threonine residuesareindicated, and thetwoidentified
phosphorylation sites (Ser8l and Thrl87)aremarked with(P).The barsrepresentthe deletedsequencesof d18 and d123. The lower lines show
theextensions of tryptic phosphopeptides 1through7(labeledwithlarge numbers); the first and the last residues areindicated.
d18, lacks the tryptic phosphopeptides 4 and 6 (see above) (Fig. 6), wasanalyzed. In this case, pronasepeptidesaand
bwerepresent,butpeptidescand f(whicharederivedfrom
the tryptic peptides 4 and 6) were missing (Fig. 12c). (The
minorpeptide seenatthe position of peptide c is probably
related to peptides a and b, since it was also observed in
some pronase digests of the tryptic peptides 1 to 3.) The presence of pronase peptide c in dl8 indicated that the
mutation of d18does notaffect phosphorylation outside the deletion(the sites contained in the carboxy-terminal portion oftryptic peptides 4 and 6). On the other hand, the absence ofpronasepeptidesaand bsuggested thatone ormoreof the
deleted serine residues (Serl5l, Serl55, and Serl56) are
phosphorylated in wild-type LT.
In summary, we have identified four tryptic phospho-peptides in a region between Met7l and Argl91 of the
polypeptide chain of LT. Three of these peptides appeared on two-dimensional peptide maps in two or more variants with overlapping sequences. Inpeptide 7, Ser8l was
identi-fiedasphosphorylated residue. In peptides 5 and 1 (and the variants of peptide 1), containing five serine residues, the phosphorylated residues were not identified. In peptides 4 and 6, Thrl87 was identified as the only phosphorylated threonine, but the serine site was again not identified. Considering the phosphoserine:phosphothreonine ratio of
6:1, and Thrl87 being the only phosphorylated threonine,we assume that about six serinesarephosphorylated.
DISCUSSION
Rough localization of the phosphorylation sites ofLT. In thisinvestigation, we analyzed the phosphorylation sites of
polyomavirus LT. Two-dimensional peptide mapping
re-vealed seven major phosphopeptides which were further characterized and localized on the polypeptide chain. The phosphorylationpatternsofmutated LTs encoded by d18or
d123 and ofatruncatedTantigen from transformed B4 cells indicated that all phosphorylation sitesarelocated within a
34Ksegmentaround the deletion in d18(residues 145to174 [45]). This conclusionwasfurthersubstantiatedby analyzing fragments of LT which were generated by partial proteolysis. The 36K and 29K fragments contained all phos-phorylation sites of intact LT, whereas a 24K fragment lackedtwo phosphopeptides thatwere also affected in dl8.
These fragments were derived from the amino terminus of
LTsincethey contained those methionine peptides that are sharedby LT, mT, and sT because of theircommonamino termini(46). The 24K and 29K fragments probably extendto
the clusters of basic residues at positions 167 and 191, respectively, which might be preferentially cleaved during limiteddigestion (42). The carboxy-terminal extension ofthe
MET-ASP-ARG*VAL-LEU-SER-ARG*ALA-ASP-LYS-GLU-ARG*LEU-LEU-GLU-LEU-LEU-LYS*LEU-PRO- 2U ARG*GLN-LEU-TRP-GLY-ASP-PHE-GLY-ARG*MET-GLN-GLN-ALA-TYR-LYS*GLN-GLN-SER-LEU-LEU- 40 LEU-HIS-PRO-ASP-LYS*GLY-GLY-SER-HIS-ALA-LEU-MET-GLN-GLU-LEU-ASN-SER-LEU-TRP-GLY- 60
THR-PHE-LYS*THR-GLU-VAL-TYR-ASN-LEU-ARG*MET-ASN-LEU-GLY-GLY-THR-GLY-PHE-GLN-GLY- 80
SER-PRO-PRO-ARG*THR-ALA-GLU-ARG*GLY-THR-GLU-GLU-SER-GLY-HIS-SER-PRO-LEU-HIS-ASP-100 ASP-TYR-TRP-SER-PHE-SER-TYR-GLY-SER-LYS*TYR-PHE-THR-ARG*GLU-TRP-ASN-ASP-PHE-PHE-120
ARG*LYS*TRP-ASP-PRO-SER-TYR-GLN-SER-PRO-PRO-LYS*THR-ALA-GLU-SER-SER-GLU-GLN-PR0-140
ASP-LEU-PHE-CYS-tyr-qlu-glu-pro-leu-leu-ser-pro-asn-pro-ser-ser-pro-thr-asp-thr-160
pro-ala-his-thr-ala-gly-arg*arg*arg*asn-pro-cys-val-ala-GLU-PRO-ASP-ASP-SER-ILE-180 SER-PRO-ASP-PRO-PRO-ARG*THR-PRO-VAL-SER-ARG*LYS*ARG-PRO-arg-pro-aia-giy-ala-thr-200 gly-gly-gly-gly-gly-gly-val-his-aia-asn-gly-gly-ser-val-phe-gly-his-pro-thr-gly-220
FIG. 11. Amino acidsequenceof the amino-terminalregionof LTasdeduced from the DNAsequence(16, 46). Cleavagesites fortrypsin
areindicatedby asterisks;serine and threonine residues indicatedaspotential phosphorylationsitesareunderlined. Thesequencesdeleted
ind18 and d123(residues145to174 and 195to228, respectively [45])arewritten in lowercase letters.
0
200ss T S a
179181 187 190
on November 10, 2019 by guest
http://jvi.asm.org/
PHOSPHORYLATION SITES OF POLYOMAVIRUS LARGE T ANTIGEN
TABLE 2. Relationships between phosphopeptides generated by digestion of tryptic peptides withpronasea
Tryptic peptide(s) digested Resultant pronase peptides
1-3 a,b, cb
4,6 c,f
5 d, h
7 e, g
a All tryptic peptides gave rise to two or more, probably overlapping, pronase peptides; the ratios between pronase peptides c and f, d and h, or e and gvaried to some extent.
bIn some cases, pronase digestion of tryptic peptides 1 to 3 revealed a peptide migrating to the position of c.
36K fragment is less clear; it might extend to Arg254 or
Arg258.
Properties of the phosphopeptides. A more precise local-ization of thephosphopeptides wasachieved by differential
labeling of LT with various amino acids. From the amino acid composition, and in accordance with the predicted amino acid sequence, the phosphopeptides were alignedas
shown in Fig. 10.
Peptide 7 extends from Met71toArg84 and is phosphory-latedatSer8l, the only serine residue in this peptide. Peptide 7a showed the same amino acidcomposition as peptide 7, indicating that the two peptides are related. This was
con-firmed by partial acid hydrolysis (24, 41)orpronasedigestion
of the isolated peptides, which revealed identical products from both peptides (data not shown). Peptide 7a had an
additionalpositive charge, sinceatpH 1.9 it migrated closer to the cathode than did peptide 7 (Fig. 2a). At pH 4.7, however, both peptides showed similarelectrophoretic
mo-bilities (datanotshown), indicating thatpeptide 7a contained anadditional acidic residue which is negatively charged at
this pH. These findings suggested that peptide 7a repre-sentedanextended form ofpeptide 7. The absence of valine and tyrosine, which would be contained in an amino-terminal extension, and the presence ofan acidic residue, which would be expected inacarboxy-terminal extensionat
position 87, indicated that phosphopeptide 7a overlaps pep-tide 7 andextends from Met7ltoArg88. Both peptidesare phosphorylatedatSer8l. It should bementioned that, during peptide mapping at pH 1.9, an unphosphorylated peptide containing isoleucine, tyrosine, and valine was superim-poseduponphosphopeptide 7. This peptide could be clearly separated from the phosphopeptideatpH 4.7 or8.9 during electrophoresis (datanotshown).
Phosphopeptide 5 extends from Gly89 to LysllO and contains fiveserine residues andonethreonine residue. Acid hydrolysis revealed phosphoserine and traces of phospho-threonine. Thus, Thr9O could theoretically be phosphoryl-ated. However, the small and varying amount of phosphothreonine suggeststhat itwasderived frompeptide 6because of insufficientseparation of thetwopeptides. The phosphorylated serine residue(s) in peptide 5 could not be identified. The electrophoretic mobility of this peptide sug-geststhat it containstwopositive chargesmorethanpeptide 7, which is compatible with the two histidine residues in peptide 5. Since these positive chargesarenotcompensated by negatively charged phosphates,we assumethatpeptide5 has the same phosphorylation state as peptide 7 and is
phosphorylated only once. Since SerlO4 and SerlO6 are
surrounded by aromatic residues, which is unusual for phosphorylation sites, weassume ratherthatSer93, Ser96, orSerlO9servesasphosphate acceptor.
a
f g,h
49a
b*d;,
b
f
9*h
, c
*a 9b
ik,
FIG. 12. Comparisonofphosphopeptides generated by pronase digestionofwild-typeord18 LTs. LTswereisolatedfrominfected and
32Pi-labeled
cells and digested sequentially with trypsin and pronase; the resulting phosphopeptides were separated on TLC plates byelectrophoresis and chromatographyas described in the legend to Fig. 2. (a) wild type; (b) d18. In (c) the 24K fragment generated by partial proteolysis of wild-type LTwasanalyzedin thesame way. Thearrows pointtopeptides ofwild-typeLT that are
missingind18orthe24Kfragment; peptidesa'and b'areprobably related to a and b, respectively, and were not reproducibly ob-served; the ratios betweenpeptides candf, d and h,or eand g, which are related to each other (see Table 2), varied in different preparations.Thevertical dashesindicate theorigin.
Phosphopeptides 1 through 3 are related to each other, since
they
showed almostidentical amino acidcompositions
(except forlysine)
and gave rise to the same cleavage products upon partial acidhydrolysis
ordigestion
with pronase(datanotshown). Theywere allphosphorylated
atserine residuesonly.Peptides1and 2extend fromThrl33to the
arginine
clusteratpositions
167to 169 andcontainfive serine residues.Peptides
la,lb,
3, and 3aoverlap
withVOL.58, 1986 813
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.612.58.301.95.149.2]SV 40
N
200 400
600
Py
~~~~~~~~400
81 123 148152 16 269
(~P)
0
600 _ 708
C
785
FIG. 13. Homology between the LTs of SV40 and polyomavirus. The linear polypeptide chains of the LT ofSV40 (upper line) and polyomavirus (lower line) were aligned according to homology (29), SV40 LT contains a unique region at its carboxy terminus, and polyomavirus LT contains unique sequences in its amino-terminal region asindicated by the inserts. The numbers refer to amino acid residues. The locations of thephosphorylation sites (P) of both proteins (37) (this study) are indicated.
peptides 1 and 2 but are extended towards the amino terminus up to Trpl23 or Lysl22, as indicated by the presence of lysine in these peptides. The extended peptides contain two additional serine residues. These are not phosphorylated, however, since thecleavage products gen-eratedby partial acid hydrolysis oftheextended peptides(la and3) wereidenticaltothosegenerated from peptides1and 2. The number and positions ofthe phosphorylated serine residues in peptides1and 2couldnotbe identified.
Consid-ering
the overallratio of phosphoserinetophosphothreonine of6:1 and thatpeptides7, 5, and 4 (seebelow) seem tobe phosphorylated only at one serine residue, it is likely that some ofpeptides 1 to 3 contain multiple phosphorylation sites. This assumption is compatible with the apparent acidity ofthese peptides duringelectrophoresis. The phos-phorylation patternofd18 differedfrom thatof wild-type LT in that thetryptic phosphopeptides 1 to 3 were altered and the pronase peptides a and b derived from these tryptic peptides were missing. Since thedeletion of d18 covers the three serine residues SerlSl, Serl55, and Serl56, one or moreof these residues mustbe phosphorylated.The heterogeneity of peptides 1 to 3 is not understood. Part of the heterogeneity may derive from incomplete or alternate cleavage bytrypsinat theamino terminus(Argl2l, Lysl22, or Lysl32), as for peptides la and 3, or at the carboxy terminus (Argl67, Argl68, or Argl69). Another
possibility
isthatCysl44 exists in different oxidationstates because of incomplete oxidation.Asimilar phenomenonhas been observed inSV40LT(37). Lastly, thepeptidesmay be differentially phosphorylated. The finding that partial acid hydrolysis generated identical products from all of these peptides indicates that theyarephosphorylatedatthesamesites,
butnotnecessarilytothesameextent. Peptides 1and la(which
appear more acidic) could bephosphorylated
at tworesidues, andpeptides2and 3 couldbephosphorylated randomly at only one of the two. The two variants ofthe phosphorylation would be indistinguishable, since they would have identical overall compositions and would mi-grate to the same position during peptide mapping. More-over, furthercleavage ofamixtureof thetwovariantswould generate the same products aswouldcleavage of thedoubly phosphorylatedform.Phosphopeptides4and 6 represent anotherpairofrelated peptides. They bothextend fromAsnl70toArgl9l and are phosphorylated at serine and threonine residues. Since Thrl87 is the only threonine residue in these peptides, it mustbephosphorylated. Whichof the threeserine residues
arephosphorylated could notbedetermined.Thedifference between peptides 4and 6 is not clear. Peptide 6 contained almost three times as muchphosphoserineasdidpeptide4. Ontheother
hand,
peptide
4wasmoreacidic,indicatingthatit contained one negative charge (perhaps one phosphate residue)more thanpeptide6.Thus, we assume that thehigh phosphoserine content ofpeptide 6 was due to contamina-tion with peptide5 (seeabove). Infact, peptide 4might be phosphorylated at Thrl87, and one serine residue and pep-tide6 might be phosphorylated only atThrl87. Partial acid hydrolysisorpronasedigestion generated identical products frombothpeptides (datanotshown),indicatingthatthey are phosphorylated at the same sites. However, as already discussed forpeptides 1and 3, peptide6 might existin two versions, onebeingphosphorylated only atThrl87 and the otheronly at oneserine residue.
Characteristics of the phosphorylation sites. Phosphory-lation sitesareoften foundin sequences with ahigh potential for ,B-turn formation (44). The two identified sites (Gly-Ser8l-Pro-Pro and Pro-Arg-Thrl87-Pro) and six ofthe po-tential phosphorylation sites (Gly-His-Ser96-Pro, SerlS1-Pro-Asn-Pro-Ser155-Serl56-Pro,
Pro-Asp-Asp-Serl79,
and Serl8l-Pro-Asp-Pro)alsofollow this rule (5).Thesequences surrounding the phosphorylated residues show no obvious homology to any of the known recognition sequences for protein kinases, thus we cannot even speculate which kinasesmight be responsibleforphosphorylation ofLT. In the case of SV40, several sites can be phosphorylated in vitro by casein kinases which recognize sites in acidic environments (3, 20), suggesting that this type ofenzyme mightparticipate in some of the phosphorylation reactions in vivo (F. Graesser, P. T. Tuazon, J. A. Traugh, K. H. Scheidtmann, and G. Walter,manuscript in prepara-tion).Comparison ofphosphorylation sites ofpolyomavirus and SV40 LT. The structuralandfunctional homology between
polyomavirus
and SV40 LT suggested that some of the phosphorylation sites might be conserved between the two proteins (40). Alignment ofthe twopolypeptidesonthebasis ofthehighest degree of homology (29) reveals fourregions
(Fig. 13). The amino-terminal region down to residue 79 shows closehomology;inthenextregion,
polyomavirus
LT contains large inserts unique to polyomavirus which areinterrupted byshort stretchesofhomology;thethirdregion, extending to the carboxy terminus of
polyomavirus
LT, shows moderate homology between the twoproteins;
the fourth region,extending
to the carboxy terminus of SV40 LT, isunique to SV40. Surprisingly, mostof the phosphor-ylation sites are in the nonhomologous regions, i.e., in regions2and 4 inSV40 and inregion2inpolyomavirus.
The onlyphosphorylation
sites that might be conserved areserineresidues 155 and 156 in
polyomavirus,
corresponding
toserine residues 111and 112 inSV40 LT. Thus, phosphor-ylation might servecompletelydifferentpurposes in thetwo
proteins.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.612.66.553.71.161.2]PHOSPHORYLATION SITES OF POLYOMAVIRUS LARGE T ANTIGEN
Possible role of phosphorylation. Phosphorylation ofLT
mightmodulateits interactions with the viralgenome orwith cellular components involved in virus replication or malig-nanttransformation. InSV40 LT, the DNA-binding activity seems to be modulated by differential phosphorylation of Serl23 and Thrl24(38), whicharelocatedwithin the origin-binding domain of the molecule (22, 32). In polyomavirus LT, the DNA-binding region has not been identified. The deletionmutantd11041, which carriesadeletion fromresidue 136 to 260 and hence covers most ofthe phosphorylation sites, is defective in origin binding (28). In d18,which lacks the potential phosphorylation sites Ser151, Serl55, and
Serl56, DNA replication is diminished (18). Thus, similar correlations between DNA binding and phosphorylation might exist for polyomavirus LT. We are presently testing this hypothesis.
T antigens in polyomavirus-transformed cells.The B4cells expressed two phosphorylated T antigens, a normal-sized 92K LT and a truncated 34K LT. Both showed phosphory-lation patterns similar to that of LT from infected cells. However, the92Kprotein was onlyweaklyphosphorylated and showed an additional phosphopeptide which was not further characterized. This additional phosphorylation site might reflect a mutation; alternatively, this site might be specificallyrecognized in ratcells but notin mousecells. In the latter case, this site must be located in the carboxy-terminal regionof LT, sinceitwasnotfound inthe truncated 34K LT. The generation ofthe 34K fragment is not under-stood. It is probably not adegradation productof LT, since it was more heavily phosphorylated than was LT itself. Alternatively,the 34Kpolypeptide mightbeexpressedfrom defective genomes (1).
It is not clear which of the two proteins fulfills the transforming functions ofLT inthese cells. These functions can be exerted by an amino-terminal fragment comprising only40% ofcomplete LT (33, 35). On the otherhand, intact LT mayeventuallyinitiate replication of theintegrated viral genomes, resulting in their excision and in the loss of the transformed state (8, 21, 27, 30). Thus, there might be a selection in the transformed cells against the replicative functions in favor of the transforming functions of LT, leadingtomutations (as inthe 92Kprotein) or to truncation (as for the 34K protein).
A role forphosphorylation in thetransforming activity of LT is not directly evident. Mutant d18, lackingtwoor three phosphorylation sites,has atransformingcapacitysimilar to that of wild type, whereas d123 is completely defective in transformation, though it contains allphosphorylationsites. However, this defect ofd123 probably resides in mT. The contribution of individual phosphorylation
e'Vents
to the variousactivities ofLT inreplication andtransformation can now be tested bysite-directed mutagenesis.ACKNOWLEDGMENTS
We thank B. Grussenmeyer and A. Haber for their excellent technical assistance. B. Griffin for kindly providing deletion mutants, T. Grussenmeyer for monoclonal antibodies specific for polyomavirus mT, and U. Friedrich and A. Clad for critically reading themanuscript.
This work was supported by the Deutsche
Forschungsgemein-schaft (SFB 31).
LITERATURE CITED
1. Birg,F.,R.Dulbecco, M.Fried, and R.Kamen. 1979. State and organization of polyomavirus DNAsequences in transformed
rat cell lines. J. Virol. 29:633-648.
2. Bonner, W. M., and J. D. Stedman. 1978. Efficient fluorography of3Hand'4Con thin layers. Anal. Biochem. 89:247-256. 3. Brignon, G., B. R. Dumas, J.-C. Mercier, and J.-P. Pelissier.
1977. Complete amino acid sequence of bovine aS2-casein. FEBS Lett. 76:251-276.
4. Carroll, R. B., and E. G.Gurney. 1982.Time-dependent matu-ration of the simian virus 40 large T antigen-p53complex studied by using monoclonal antibodies. J. Virol. 44:565-573.
5. Chou,P.Y.,andG.D.Fasman. 1978. Empirical predictions of proteinconformation. Annu.Rev. Biochem. 47:251-276. 6. Clark, R., M. J. Tevethia, and R. Tjian. 1984. The ATPase
activity of SV40largeT antigen, p. 363-368. In G. F. Vande Woude, A. L. Levine, W. C. Topp, and J. D. Watson (ed.), Cancer cells.2.Oncogenes andviral genes. ColdSpringHarbor Laboratory, Cold Spring Harbor, N.Y.
7. Clertant, P., and F. Cuzin. 1982. Covalentaffinity labeling by periodate-oxidized(y-32)ATP of the large-Tproteins of polyoma andSV 40 viruses. J. Biol. Chem. 257:6300-6305.
8. Colantuoni, V., L.Dailey, G. Delia Valle, and C. Basilico. 1982. Requirements for excision and amplification of integrated viral DNA molecules in polyoma virus-transformed cells. J. Virol. 43:617-628.
9. Cowie, A., and R. Kamen. 1984. Multiple binding sites for polyomavirus large T antigen within regulatory sequences of polyomavirus DNA. J. Virol. 52:750-760.
10. Eckhart, W., M. A. Hutchinson, and T. Hunter. 1979. An activity phosphorylatingtyrosine in polyoma Tantigen immu-noprecipitates. Cell 18:925-933.
11. Fanning, E., B. Nowak, and C. Burger. 1981. Detection and characterization of multiple forms of simian virus 40 large T antigen. J. Virol. 37:92-102.
12. Friedmann,T., A.Esty,P. La Porte,and P.Deininger. 1979. The nucleotide sequence and genome organization of the polyoma early region: extensive nucleotide and amino acid homology with SV 40. Cell 17:715-724.
13. Gaudray, P., P. Clertant, and F.Cuzin. 1980. ATP phosphohy-drolase(ATPase) activity of apolyoma virus T antigen. Eur. J. Biochem. 109:553-560.
14. Gibson, W. 1974. Polyoma virus proteins: adescription of the structural proteins of the virion based on polyacrylamide gel electrophoresis andpeptide analysis. Virology 62:319-336. 15. Greenspan, D. S., and R. B. Carroll. 1981. Complex of simian
virus 40 large tumor antigen and 48,000-dalton host tumor
antigen. Proc. Natl. Acad. Sci. USA78:105-109.
16. Grifin, B. E., and S. M. Dilworth. 1983. Polyomavirus: an
overview of its unique properties. Adv. Cancer Res. 39:
183-268.
17. Griffin, B. E., M. Fried, and A. C. Cowie. 1974. Polyoma DNA: aphysical map. Proc. Natl. Acad. Sci. USA 71:2077-2081. 18. Griffin, B. E., and C. Maddock. 1979. New classes of viable
deletionmutants in the early region of polyoma virus. J. Virol. 31:645-656.
19. Grussenmeyer, T., K. H. Scheidtmann, M. A. Hutchinson, W. Eckhart, and G. Walter. 1985. Complexes of polyomavirus mediumTantigenand cellularproteins. Proc. Natl. Acad. Sci. USA82:7952-7954.
20. Hoppe, J., and H. Baydoun. 1981. Substrate specifity of the nuclear protein kinase N II from porcine liver: studies with casein variants. Eur. J. Biochem. 117:585-589.
21. Hutchinson, M. A., T. Hunter, and W. Eckhart. 1978. Charac-terization of T antigens in polyoma-infected and transformed cells. Cell 15:65-77.
22. Kalderon, D., and A. E. Smith. 1984. In vitro mutagenesis ofa
putative DNA binding domain of SV 40 large-T. Virology
139:109-137.
23. Laemmli, U. K. 1970. Cleavage of structural proteins duringthe
assembly of the head of bacteriophage T4. Nature (London)
227:680-685.
24. Light, A. 1967. Partial acid hydrolysis. Methods Enzymol. 11:417-421.
25. Mann, K., T. Hunter, G. Walter, and H. K. Linke. 1977.
Evidence for simian virus 40 (SV40) coding of SV40T-antigen
VOL.58, 1986 815
on November 10, 2019 by guest
http://jvi.asm.org/
and the SV40-specific proteins in HeLa cells infected with
nondefective adenovirus type2-SV40 hybridviruses. J. Virol. 24:151-169.
26. Morrison, B., M. Kress,G. Khoury, and G. Jay. 1983. Simian
virus 40tumor antigen: isolation of the origin-specific
DNA-binding domain.J.Virol. 47:106-114.
27. Muller, W. J., M. A. Naugjokas, and J. A. Hassell. 1984.
Isolation of largeTantigen-producingmousecelllinescapable
of supporting replication of polyomavirus-plasmid
recombi-nants.Mol. Cell. Biol. 4:2406-2412.
28. Nilsson, S. V., and G. Magnusson. 1984. Activities of polyo-mavirus large-T-antigen proteinsexpressed bymutantgenes.J.
Virol.51:768-775.
29. Pawlita, M., A. Clad, and H.zurHausen.1985.CompleteDNA
sequence of lymphotropic papovavirus: prototype of a new
speciesof the polyomavirusgenus. Virology 143:196-211.
30. Pellegrini, S., L. Dailey, and C.Basilico. 1984.Amplification and
excision ofintegrated polyomaDNAsequencesrequirea
func-tionaloriginofreplication. Cell36:943-949.
31. Pomerantz, B. J., C. R. Mueller, and J. A. Hassell. 1983.
Polyomavirus large Tantigenbinds independently tomultiple, uniqueregions ontheviralgenome.J.Virol. 47:600-610.
32. Prives, C.,B. Barnet,A. Scheller,G. Khoury, and G. Jay.1982. Discreteregions of simian virus40largeTantigenarerequired
fornonspecific and viralorigin-specific DNAbinding. J.Virol.
43:73-82.
33. Rassoulzadegan, M., A. Cowie, A. Carr, N. Gleichenhaus, R. Kamen, and F. Cuzin. 1982. The roles of individual polyoma virusearlyproteins in oncogenictransformation. Nature
(Lon-don)300:713-718.
34. Rassoulzadegan, M., P. Gaudray, M. Canning, L.Trejo-Avila,
and F.Cuzin.1981.Twopolyomavirusgenefunctionsinvolved
intheexpression of thetransformedphenotypein FR 3T3rat
cells. Virology 114:489-500.
35. Rassoulzadegan, M., Z. Naghashfar, A. Cowie, A. Carr, M. Grisoni, R. Kamen, and F. Cuzin. 1983.Expressionof thelarge
T protein of polyoma virus promotes the establishment in culture of "normal" rodent fibroblast cell lines. Proc. Natl.
Acad. Sci. USA 80:4354-4358.
36. Schaffhausen,B., and T. L.Benjamin.1979.Phosphorylationof
polyomaTantigens. Cell 18:935-946.
37. Scheidtmann,K.H.,B.Echle,andG. Walter.1982. Simian virus 40large T antigen is phosphorylatedatmultiple sites clustered in two separateregions.J. Virol.44:116-133.
38. Scheidtmann, K. H., M. Hardung, B. Echle, and G. Walter.
1984. DNA-binding activity of simian virus 40 largeTantigen correlates with a distinct phosphorylation state. J. Virol. 50:1-12.
39. Scheidtmann, K. H., A. Kaiser, A. Carbone, and G. Walter.
1981.Phosphorylation of threonine intheproline-rich carboxy-terminal region of simian virus 40 large T antigen. J. Virol. 38:59-69.
40. Scheidtmann, K. H.,A.Kaiser, and G.Walter. 1981. Phosphor-ylation sites in SV40largeTantigen, p. 1273-1279.In0.M. RosenandE.G. Krebs(ed.), ColdSpring Harbor conferences
on cell proliferation, vol. 8: protein phosphorylation. Cold Spring Harbor Laboratory, ColdSpring Harbor,N.Y. 41. Schulz, J. Cleavage at aspartic acid. Methods Enzymol.
11:793-801.
42. Schwyzer, M., R.Weil, G. Frank, and H. Zuber. 1980. Amino acid sequence analysis of fragments generated by partial proteolysis from large simian virus 40tumorantigen. J. Biol.
Chem.225:5627-5634.
43. Silver, J., B. Schaffhausen, and T. L. Benjamin. 1978. Tumor
antigens induced bynontransforming mutants of polyoma virus. Cell15:485-496.
44. Small, D.,P. Y. Chou,andG. D. Fasman. 1977.Occurrenceof phosphorylated residues in predicted P-turns: implications for P-turnparticipationincontrol mechanisms. Biochem. Biophys.
Res.Commun.79:341-346.
45. Smolar, N.,and B. E.Griffin. 1981. DNAsequences of polyoma virus earlydeletionmutants.J. Virol.38:958-967.
46. Soeda, E., J. R. Arrand, N. Smolar, and B. E. Griffin. 1979. Sequence from the early region of polyoma virus DNA that contains theorigin ofreplication and codes for small, middle and (part of) large T-antigens. Cell 17:357-370.
47. Triesman, R., U. Novak, J. Favaloro, and R. Kamen. 1981.
Transformation ofratcellsby an altered polyoma virus genome encoding only the middle-T protein. Nature (London) 292:
595-600.