0022-538X/86/040050-09$02.00/0
Copyright© 1986, AmericanSocietyforMicrobiology
Structure and Processing of the p2 Region of Avian
Sarcoma
and
Leukemia Virus
gag
Precursor
Polyproteins
R. BLAKEPEPINSKY,l* ROBERT J. MATTALIANO,' AND VOLKERM. VOGT2
Department of Protein Chemistry, Biogen Research Corp., Cambridge, Massachusetts
02142,1
and Section of Biochemistry, Molecular and Cell Biology, Cornell University, Ithaca, New York 148532Received 25September 1985/Accepted 25 November 1985
We have purifiedtwo low-molecular-weight polypeptides from the Prague C strainof Roussarcomavirus
and have identified these as products of thegag precursorPr76 by protein sequencing and by amino acid analysis. Bothpolypeptidesarederived fromastretch of 22 amino acids within Pr76 that separatesp19 and
p10.We refertothis regionasp2. Together thetwocleavageproductsformtheentirep2 region.Thejunctions ofp19 withtheamino-terminalfragment of p2 and of plO withthecarboxy-terminalfragment of p2 definetwo new processing sites withinthegagprecursor,Tyr-155-His-156 andGly-177-Ser-178. Bothpolypeptidesare
majorcleavageproductsofPr76 thatoccurinPragueCRoussarcomavirusatanestimated1,000copiesper
virion. They alsoareprominentcomponentsofavianmyeloblastosis virus. The combination of gel filtration and reverse-phasehigh-pressure liquid chromatography, whichwasusedfor the isolation of thetwofragments of
p2, resolvedover adozen otherlow-molecular-weight polypeptides fromaviansarcomaandleukemiaviruses
that previously were undetected. This technique thus should serve as a useful procedure for further characterization of viral components.
In avian sarcoma and leukemia viruses (ASLV) the gag precursorpolyprotein Pr76 is processed proteolytically into the fivemature proteins (p19, plO, p27, p12, and p15) that form the internal structure ofthe virus particle. Of these products, p19 is associated with the viral envelope (9, 15), plOis withinthespacebetween the membraneandcore(16),
p27forms the core shell (4,24), and p12 iswithin the core associated with RNA inaribonucleoprotein complex (4, 6).
The location of p15,theproteaseresponsible for cleaving the viral polyproteins during assembly (7, 26), is unknown. Proteolytic processing of the gag precursor also generates minorcleavage products such aspl9f (22)andp23(17)that
are less well characterized and that vary in amount in different viruses andvirus preparations. Both pl9fandp23
arerelated proteins thatdiffer fromp19onlyattheircarboxy termini. pl9f is a 10-kilodalton (kDa) proteolytic fragment
derived fromthe amino-terminal portion of p19, and p23 is anincompleteprocessing productof Pr76 that contains all of
p19 plus p2 (25; see below).
Attwopositions within thegag precursor-the junctions ofp19 (carboxy terminus at amino acid 155; 25) with plO (amino terminus at amino acid 178; 13) and p27 (carboxy terminusatamino acid 479; 1)with p12 (aminoterminusat amino acid489;K. S.Misono, F. S. Sharief,andJ. P. Leis, Fed. Proc. 60:1611, 1980)-theendsof the matureproteins
are not contiguous with each other, but are separated by
short stretches of amino acids. Theregionbetweenp19 and
p10,whichwe refertoasp2, is composed of 22 amino acid residues. To determine the fate of this polypeptide in the mature virus, we used a combination of gel filtration and
high-pressure liquid chromatography (HPLC) to isolate small polypeptides from virus particles, which then were
characterized by amino acid analysis and by protein
se-quencing. Inthisreportwe describe theisolation and char-acterization oftwopolypeptidesderivedfrom thep2 region,
a cysteine-rich fragment from its amino terminus and a proline-rich fragment from its carboxy terminus. Both are
* Correspondingauthor.
prominent species thatoccurin roughly equimolaramounts in mature virus particles. Based on theirpresence in both avian myeloblastosis virus(AMV) and the Prague C strain of Roussarcomavirus(PrC-RSV),weinfer thattheyaremajor
cleavage products of the ASLVgag precursorpolyprotein
generated during virus maturation.
MATERIALS AND METHODS
Virus. Chicken embryo fibroblasts infected either with PrC-RSVorwith AMVweregrownasmonolayersat39°Cin Dulbecco modified Eagle medium containing antibiotics (penicillin, 50 U/ml; streptomycin, 50 ,ug/ml) and 10%fetal
calfserum. Metabolicallylabeledviruseswerepurified from
the supernatants of infected cells as described previously
(17). Unlabeled AMV from leukemic chicken plasma was obtained from Life Sciences Inc. UnlabeledPrC-RSV,inthe form of a concentrated suspension from supernatants of virus-infected cellcultures,wasobtainedfrom theBiological Carcinogenesis Branch of the National Cancer Institute. Proteins were identified by sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) as
previ-ouslydescribed (16).
Carboxymethylation of viral proteins. For each analysis presented, viralproteins werefirst carboxymethylated with iodoacetic acid. Briefly, each preparation of virus was
suspendedataproteinconcentration of5mg/mlin 200 p.lof
reducing solution (6 M guanidine hydrochloride, 0.2 M N-ethylmorpholine acetate [pH 8.6], 5 mM dithiothreitol) and incubated at 37°C for 2 h. Fresh iodoacetic acid was addedto afinal concentrationof 10 mM(from a lOx stock
solutionin 0.2 MN-ethylmorpholine), and thesampleswere
incubatedat23°Cfor 30minin the dark. Unreactedreagent wasquenched with 25 mM dithiothreitol. In instances where
proteinwastobe labeledsimultaneously, thereduced
prep-arationwas first exposed to 250 p.Ci ofiodo[3H]acetic acid (193 mCi/mmol; New England Nuclear Corp.) for 10 min,
and then iodoacetic acid was adjusted to a final concentra-tion of 10 mM for the remainder of the reacconcentra-tionperiod.
HPLC analysis. Viralproteinswere subjected to reverse-50
on November 10, 2019 by guest
http://jvi.asm.org/
(Vydac; catalog no. 214TP54) and C18 (SpectraPhysics, octadecylsilyl; catalog no. A2351-040), are both 0.46 cm
(inside diameter) by 25 cm.
Protein sequence analysis. Samples containing either 1
nmol of peptide or 50,000 cpm of peptide in combination with3 nmolofanunlabeledcarrier proteinweresubjectedto
amino-terminal sequenceanalysis by sequentialEdman deg-radation on an Applied Biosystems 470A gas-phase se-quencerinthe presenceofpolybrene. Phenylthiohydantoin-amino acids from each cyclewereanalyzed by reverse-phase HPLC on a 5-,um cyano column (25 by 0.46 cm; IBM
Instruments, Wallingford, Conn.) as described (12). The
column effluent was monitored
spectrophotometrically
atboth 254 and 313 nm. For radioactive
polypeptides,
each cycle ofthe Edman degradation was subjected to scintilla-tion counting.RESULTS
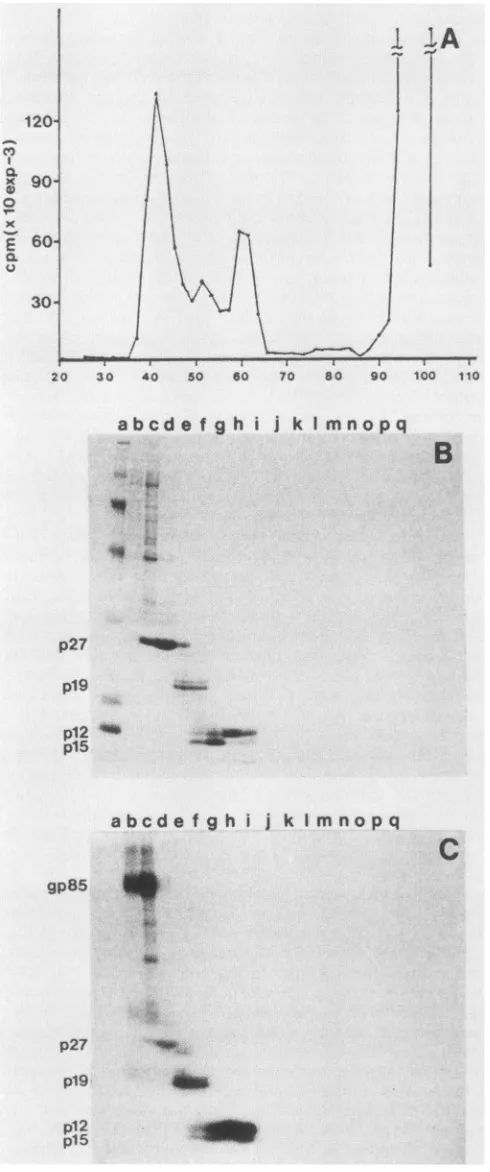
Reverse-phaseHPLC analysisof ASLVproteins. The
major
proteins in ASLVarethe structural
proteins
thatarederivedfrom the gag and env genes. The five internal structural
proteins (gag proteins) account for about 80% ofthe total viral protein, while the two surface
glycoproteins
(envpro-teins) make up about 20%. When
carboxymethylated
pro-teins from PrC-RSVwereresolved
by reverse-phase
HPLCon aC4 column, we obtained the
simple
absorbanceprofile
shown in Fig. 1A. Proteins from each ofthe
peak
fractionswere identifiedby SDS-PAGE. Thefive
major peaks,
desig-nated c, d, f, g, and h in
Fig.
1A, contain the five gagproteins
p12,
plO, p15, p19, and p27,respectively.
Peak econtains gp85. Other
prominent
peaks-i (p19), j (p19),
and k (p27)-are chromatographic variants ofpeaks
g andh,
which vary in relative intensity from analysis to
analysis.
They are indistinguishable from the
major
peaks by
SDS-PAGE. In fact, by gel
analysis
each ofthe threepeaks
forp19
was found to contain both thephosphorylated
andunphosphorylated forms ofthe protein as well as the two
related cleavage products,
pl9f
and p23. gp37 was notidentifiedas acomponentofanyofthe
prominent peaks.
A214 values for each ofthe peaks that were identified in
Fig. 1 are summarized in Table 1.
Correcting
both for thesplitting of p27 and
p19
intomultiple peaks
and for theappropriate size adjustments oftheabsorbancevaluesofthe individual peaks, we conclude that p27,
p19, p15,
andpl2
wererecoveredroughlyinequimolaramounts. On the other
hand,the single peak for plO containedonly 0.5 mol
equiv-alents ofprotein. Similar estimates were obtained for plO from histidine-labeled AMV (described in a later section). Since all five gag proteins are equimolar when virus is
analyzed by SDS-PAGE (16), a likely
explanation
for the observed discrepancy is thatother unidentified peaks existfor
p10.
In addition to the five major gag proteins, proteolytic processing of the gag precursor also should generate a
22-amino acid polypeptide that within the precursor spans theregion betweenthecarboxyterminus of
p19
(25)and theaminoterminusof plO(13). We have
designated
thisregion
provisionally as p2. The predicted amino acid sequence of
p2,based on DNA sequencing (19), contains three
cysteine
boxymethylated
proteins
from[35S]cysteine-labeled
PrC-RSV were
subjected
toreverse-phase
HPLC. Fractionswere
quantitated by
scintillationcounting.
Thesingle large
peak, peak
c, containsp12,
themajor
cysteine-containing
protein
inPr76.Therelative sizes of the otherpeaks
alsoareconsistentwith their
cysteine
content. This result isreadily
apparent from Table1,
where the observed andpredicted
valuesforthecysteine
contentof eachofthe viralstructuralproteins
are summarized.In the column
profile
fromcysteine-labeled PrC-RSV,
asingle
newpeak
(designated
a inFig.
1B)
wasconsistently
observed. We have shown
by
amino acidanalysis
andby
protein
sequencing
(seebelow)
that thispeak
contains afragment
ofp2.
Acorresponding peak
was observed in theabsorbance
profile
from unlabeledvirus(Fig. 2A),
butwasminor
by
its absorbance.Using
the values summarized inTable 1, we infer that the
cysteine-rich
fragment
ofp2
is amajor cleavage
product
ofgag precursor, since relative toothergag
proteins
itoccursin virusatapproximately
0.5molequivalents.
Purification of
low-molecular-weight polypeptides.
Weused acombination ofgel
filtrationchromatography
andreverse-phase
HPLC to isolatelow-molecular-weight proteins
from PrC-RSV. Beforechromatography,
viralproteins
were la-beled andcarboxymethylated
withiodo[3H]acetic
acid.Fig-ure 2Ashows the elution
profile
fromgel
filtrationchroma-tography
in which viralproteins
were resolvedon anA.5M resin in 6 Mguanidine
hydrochloride.
Portions offractionswere
subjected
toSDS-PAGE,
and theproteins
were visu-alized eitherdirectly by
Coomassiestaining
(Fig. 2B)
orby
fluorography
(Fig.
2C). Thepeak
fractions forgp85,
p27,
p19,
p15,
andp12
were fractions40, 44, 50, 57,
and59,
respectively.
In theseanalyses
plO
was notidentified,
pre-sumably
because it does not containcysteine (13)
and does not stain well with Coomassie blue(16).
Fromfractions 65through 100,
nolow-molecular-weight
proteins
werede-tected
by
SDS-PAGE.By
contrast, overadozen differentlow-molecular-weight
polypeptides
were detected when the samegel
filtration fractionswereanalyzed by reverse-phase
HPLC.They
werecharacterized
initially by
their
A214
andA280
andby
carboxymethyl-[3H]cysteine
content. HPLCprofiles
fromnineserialsetsof
gel
filtration fractionsarepresented
inFig.
3.Chromatographs
at the left showA214,
and those at theright
show the distribution ofradioactivity
from the samechromatography
fractions. For eachanalysis
thedesignated
gel filtration
fractions(from
fraction75to95)
werecombinedand loaded
directly
on a C18 column. Bound components were eluted with agradient
ofacetonitrile from0 to75%in 25 mMammoniumacetate(pH 6.5).
Themolar amountsof each of the
polypeptides
detected wereestimated, using
their apparent size based ongel
filtration and their relative absorbance. Fortwocomponents inparticular,
onecontaining cysteine
buthaving
noA280
value
(peak
a inprofiles
86through 92)
and onehaving
anA280value butno
cysteines (peak
b inprofiles
86through
92),
we estimate that
they
both occur atapproximately 1,000
copies
per virion. Thusthey
wereassumedtobeproducts
of the gag gene. These numbers agree with molar estimateson November 10, 2019 by guest
http://jvi.asm.org/
c
to 0
Db.
0 0 .0 a
a b
C
f
h
A
10 20 30 40 so 60 70 so 90
a
c
130 140 150 160
B
10 20 30 40 so 6o 70 so 90 100 110 120 130 140 1SO 160
fraction number
FIG. 1. HPLC analysis ofPrC-RSV proteins. Preparations ofPrC-RSV, which were carboxymethylated with iodoacetic acid before analysis,weresubjectedtoreverse-phase HPLCon aC4column.Proteinswereeluted with agradient of acetonitrile from 0 to75%in0.1% trifluoroaceticacid. (A)Elutionprofile fromunlabeled virus (50 ,ug), monitored at 214 nm. (B)Elutionprofile from[35S]cysteine-labeledvirus. InTable 1 theobserveddata are summarized quantitatively.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.100.540.51.673.2]a p2a 9(10) 0.53 0.6(0.5) 2(3) 1,000
b p2b 11 0.69 0.6 0 0
c p12 89 9.3 1.0 6 10,800
d plO 62 3.2 0.5 0 0
f p15 124 11.3 0.9 1 1,200
g p19 155 11.5 0.7 4 3,600
h p27 240 21.0 0.8 1 1,500
env
polypeptides
e gp85 341 5.9 0.2 14 4,300
gp37 198 7
Other peaks
i p19 155 4.2 0.2 4 0
j p19 155 3.9 0.2 4 540
k p27 240 5.5 0.2 1 827
aNumbers arepredicted from the DNA sequence ofPrC-RSV (19).
bNumbers are calculated from amino acid sequence and A214 readings.
Values listed are moles relative to p12.
cNumbers are derived from the analysis of carboxymethylated,
[35S]cysteine-labeledviralproteins. For all values listed,abackground of500 cpm wassubtracted.
takenfrom HPLCprofiles of intact virus in
Fig.
1.Theother low-molecular-weight peptides that were detected occur atmuch lower copy numbers and remain to be characterized further.
The two prominent low-molecular-weight
polypeptides
were purified to apparent homogeneity with a second re-verse-phase HPLC step. Peak fractions from the initial run were combined, diluted 1:2 with water, and acidified with
trifluoroacetic acid. They then were loaded
directly
onto aC4 column, and bound components were eluted with a
gradient of acetonitrile from0 to75% in 0.1% trifluoroacetic acid. For thetwomajor peaks described above,weobtained theprofiles shown in Fig.4.Figure4Ais thechromatograph from the major cysteine-containing fragment. All of the
tritium label eluted with thesingle peakatfraction34. This
peptide does not have anA280 value. Figure4B shows the
profilederivedfromthe otherfragment. It has anA280 value butdoes not havecysteine countsassociated with it.
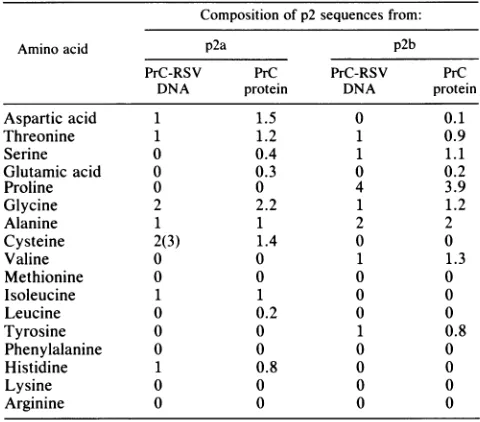
Primary structure of the p2 polypeptides. The cysteine-containing polypeptideshown inFig.4A,p2a, wasidentified
as afragment ofp2 by amino acid analysis and by protein sequencing. Before these analyses, two results suggested
that this polypeptide was onlypart oftheregion. First, the
fragment eluted fromthe gelfiltrationcolumn asif itwere 1 kDa rather than thepredicted size of2.4 kDa based on the sequence of the intact region. Second, thefragmentlacked anA280 value. The p2regionhas atyrosinenearits carboxy
terminus. The data in Table 2 summarize the results from amino acid analysis of the cysteine-rich fragment. The observed composition agrees exactly with the predicted
composition for a polypeptide derived from the
amino-terminal halfofp2. ItcontainsHis,Cys, Asx, Thr, Gly, Ala, andIle and is devoid of Lys, Arg, Val, Leu, Phe, Pro, Tyr, and Met. The lower than expected value for cysteine is due tolow recovery of carboxymethyl-cysteine by our
hydroly-sis procedure. In particular the absence of Pro and Tyr,
whichareunique markers in p2 for the carboxy-terminal half
oftheregion, confirm that onlyaportionof the molecule is
rizes these data. Since
radioactivity
wasreleased incycles
2and
8,
we concludethat thesepositions
within thepolypep-tide contain
cysteine.
We infer from this result that thefragmentstarts atHis-156 inthe
predicted protein
sequenceofPr76 (19), since suchan
alignment
wouldplace cysteines
at
positions
2 and 8. This is theonly position
in the entire sequence ofPr76where twocysteines
areseparated by
sixamino acids. A
major implication
of this result is that theaminoterminus of
p2
isimmediately adjacent
tothecarboxy
terminus of
p19
and thus definesprecisely
a newcleavage
site within the gag precursor,
Tyr-155-His-156.
From the sequenceanalysis
we were unable toidentify
thecarboxy
terminus of the
peptide; however,
basedon the amino acidcomposition presented
in Table2,
we canplace
restrictions onits size. We infer that thefragment
muststopat orbefore thecysteine
atposition
10inp2,
sinceitcontainsAsx(found
only
atposition
9 inp2),
butnot twoAla(the
second Ala inp2 is foundat
position 11).
Apolypeptide
ofthis size agrees with the apparent molecularweight
ofp2a
based ongel
filtration
analysis.
The
primary
structure of the secondprominent
low-molecular-weight polypeptide, p2b,
was also determined(Fig. 4B).
The amino acidcomposition
of thispeptide
is shown in Table 2. Itcontainsonly
sevenaminoacids,
Pro,
Ala,
Ser, Thr, Val, Gly,
andTyr,
which suggests that it is derivedfrom thecarboxy-terminal
halfofp2.
Thefact that thefragment
does not containcysteine implies
that its sequence does notoverlap
with the firstpeptide.
To better evaluate its structure,p2b
wassubjected
to 10cycles
ofprotein
sequencing.
From thisanalysis
we obtained the sequenceThr-Ala-Ser-Ala-Pro-Pro-Pro-Pro-Tyr-Val,
which agreesexactly
with thepredicted
sequenceforthecarboxy
terminus of
p2.
Although
wedidnotsequencetotheendofthe
peptide,
thefragment presumably
endsatGly-177,
based on its amino acidcomposition.
This resultimplies
that the end ofp2
iscontiguous
withplO
and thus defines anadditional
cleavage
site in the gag precursorGly-177-Ser-178.
Together
thecysteine-
andproline-rich polypeptides
makeup the entire
p2
region.
Theirorganization
with respectto the gag precursor is shown inFig.
6.They
aremajor
cleavage products
ofthe gag precursor thatareretained in the mature virusparticle.
In thepreparations
of PrC-RSVthat were
analyzed,
therewasnoevidence ofapolypeptide
that
corresponds
to the intactp2
region. However,
in our screenforp2,
wehavenotcharacterizedspecies
thatoccur at less than 10% ofthemajor
gagproteins.
Theonly
otherlow-molecular-weight cysteine-containing fragment
thatwassubjected
to sequenceanalysis (fraction
54 inprofiles
74through
79ofFig.
3)
hadablockedamino terminusand thuspresumablyis derivedfromsomeotherprotein (not shown). p2 polypeptidesinAMV.To determine whether the
proc-essing
pattern forp2
inPrC-RSV isacommonphenomena
of ASLVorisspecific
tothatvirusstrain,
wealsoanalyzed byHPLC the
protein
component of AMV. Aspurified
fromleukemic chicken
plasma,
AMV isactually
a mixture ofmyeloblastosis-associated
viruses and AMV itself. Theseclosely
related viruses have beensequenced only partially,
but amino acid
analyses (8, 11), immunological properties
on November 10, 2019 by guest
http://jvi.asm.org/
Cl) a 'C
x 4)
0
x
a
C
20 30 40 5o 60 70 80 90 100 110
abcdef ghi j k I mnopq
B
p27 0
p19
p12 "'
p15
.r_
abcde f g h i jk I mnopq
4.q
gp85 _
S.
p27 p19 p12
p15
do.
am
C
30 60 90 120 0 30 60 90
fraction nummber fraction number
120
FIG. 3. HPLC chromatographs of low-molecular-weight pro-teins. Sequential sets of gel filtration samples, which contained low-molecular-weight components from PrC-RSV, were further fractionated by reverse-phase HPLC. The gel filtration fractions indicated in the middle of the figure were combined and loaded directlyonto aC18 column. Boundcomponents wereeluted witha
gradient of acetonitrile from 0to75% in 25 mM ammoniumacetate
(pH 6.5). Profiles atthe left(column A) showA214. Profilesat the right (column B) show the distribution of radioactivepolypeptides from thesame setsoffractions.
(3),andexisting sequencedataforsomeof the gagproteins (18) suggest that theproteinsfrom PrC-RSVand AMV and its helper viruses are all highlyconserved. By SDS-PAGE the apparent sizes of the structural proteins from the two viruses are virtually identical, except for plO, which mi-gratesanomalously in SDS gels (16).
When [35S]cysteine-labeled AMV (carboxymethylated
with iodoacetic acid) was subjected to HPLC,we obtained the profile shownin Fig. 7A. The major proteinsp12, plO, gp85,p15, p19,andp27elutedatexactlythesamepositions
in the gradientas the corresponding peaksfrom PrC-RSV. This result is based on asimultaneousanalysisofradioactive FIG. 2. Gel filtration analysis of PrC-RSV proteins. A 2-mg
sample of virus was carboxymethylated with iodo[3H]acetic acid and thensubjectedtogel filtrationon anA.5Mcolumn(1by40cm) in 6 M guanidine hydrochloride-0.05 M sodium phosphate (pH
5.4)-10
mMdithiothreitol.Chromatography
waspreformed
atroomtemperature with aflow rate of 1 ml/h. Fractions of0.3 ml were
collected. Samples of fractions were analyzed either directly by
scintillationcounting(A)or,afterSDS-PAGE, byCoomassie stain-ing (B) orfluorography (C). Forgel analysis, proteins were first selectively precipitated from the guanidine hydrochloride with ethanol(16). Lanes bthroughqcorrespondtogelfiltration fractions 39, 43, 47, 50, 53, 56, 59, 62, 65, 68, 71, 74, 77, 83, 89, and97, respectively.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.326.562.72.409.2] [image:5.612.70.315.80.666.2]0 0 as
Q .0
0
a'
U .0
0 .0
'U
Vv.
10 200 0 40 50 60 70 *0 90 100 110 120 130 140
B
10I
5 I I 510 20 30 40 50 60 70 s0 910 100 110 120 130 1140
fraction number
FIG. 4. Rerunsof p2fragmentsonHPLC. Fractionscontainingthemajorlow-molecular-weight peptidesdetected inprofilesshown inFig. 3wererechromatographedon aC4 column. Bound components were eluted withagradient of acetonitrile(0to
75%)
in 0.1%trifluoroacetic acid.Fractionsweremonitored bothat214 nm(upper trace,0.1full-scaledeflection)andat280nm(lowertrace,0.01full-scaledeflection). (A)Column profile of cysteine-rich p2fragment. (B) Column profile of proline-rich p2 fragment.AMVwith unlabeled PrC-RSV. The single major difference between theprofileswasintheregion ofthecysteine-rich p2 fragment. Whereasasingle
[35S]cysteine
peakwasobserved in the chromatograph from PrC-RSV, three peaks wereobserved in the
profile
fromAMV. The first peak coeluted with the cysteine-containing p2 fragment from PrC-RSV, while the two other peaks eluted at slightly higher acetonitnle concentrations. We obtained the same resultwith AMV that was isolated from chicken cells grown in
culture and from leukemic chickens.
To decidewhether the additional peaks specificto AMV were also derived from the p2 region, we used HPLC to
analyze radioactive AMV labeled metabolically with
histi-dine. The single histidine in p2 accounts for
10%
ofall the histidines in Pr76, and thus, as with cysteine, metabolic labelings withhistidine should generate an apparent enrich-ment for p2 in virus. When histidine-labeled AMV wassubjectedtoHPLCweobtained theelutionprofile shown in
Fig.
7B. Themajor peakin the chromatograph correspondsto
p15,
whichisthemajorhistidine-containinggag protein in ASLV. Likewise,the sizes of peaks correspondingto each of the other proteins were consistent with their histidinecontent.
p19,
whichdoes not containhistidine (25),wasnotdetected inthechromatograph. Asobservedwiththe
cyste-ine-labeledAMV,thereweremultiple peaksin the region of the p2 fragments, suggesting that the peptides in these
additional peaksarederived from the p2region.
The two largest peaks in the triplet were analyzed by
protein sequencing. In this analysis, proteins from AMV that were carboxymethylated with iodo[3H]acetic acid were re-solvedby HPLC, and the appropriatepeakswere
subjected
to sequenceanalysis. Each cycle was analyzed by
scintilla-tion counting. The histogram shown in Fig. SB is derived
from sequence analysis of the first of the peaks in the
chromatogram. Radioactivity was detected in cycles 2, 8,
and 10. Thus the peak is derived from p2. However,unlike preparations from PrC-RSV, the protein sequence of the AMV protein has aphasing problem that is apparent in later sequencing cycles. As a result of this problem, cycle 9 has more countsthancycle 8 and cycle 11 islargerthancycle 10. Such aphasing problem is often caused by a proline in the peptide sequence and suggests that the AMVpeptide may differslightlyin sequence from thecorrespondingprotein in PrC-RSV.However,thesequence difference was not
signif-icant enough to change the peptide's elution pattern from HPLC.
For the second AMV peptidesequenced weobtained the
same histogram that is shown in Fig. SB. Thus,both peaks
6
e%A&.4k, .
Ilawlwolw",-W.. I
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.125.466.66.439.2]contain the amino terminus of the p2 region. Although we were unable to determine the source of the heterogeneity,
neitherpeak appears to be intact p2 since they both lacked A280. Furthermore, a peak was detected in the absorbance profile that corresponds to the C-terminal fragment of p2 in both its elution position and the fact that it has an A280 value. Based on our results from PrC-RSV and AMV, we conclude that the proteolytic processing that we have characterized
14
A
12
C 10
x
0
I-l
E
0.
.4-2
I
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
cycle number
6
C')
5
x 0
4.
0 I-.x3
E
0
2
L
[image:7.612.328.568.84.295.2]B
TABLE 2. Amino acidanalysis of p2polypeptidesa
Composition ofp2sequences from:
Aminoacid p2a p2b
PrC-RSV PrC PrC-RSV PrC
DNA protein DNA protein
Aspartic acid 1 1.5 0 0.1
Threonine 1 1.2 1 0.9
Serine 0 0.4 1 1.1
Glutamicacid 0 0.3 0 0.2
Proline 0 0 4 3.9
Glycine 2 2.2 1 1.2
Alanine 1 1 2 2
Cysteine 2(3) 1.4 0 0
Valine 0 0 1 1.3
Methionine 0 0 0 0
Isoleucine 1 1 0 0
Leucine 0 0.2 0 0
Tyrosine 0 0 1 0.8
Phenylalanine 0 0 0 0
Histidine 1 0.8 0 0
Lysine 0 0 0 0
Arginine 0 0 0 0
aFor amino acidcompositions,250 pmolofeachpeptidewashydrolizedin
6 NHCI(Pierce Chemicals)at110°Cfor 24 h insealed evacuated ampoules and then analyzed for amino acid content on a Beckman System 6300 analyzer.Valueslistedaremolesof each amino acidpermole offragment.
Zerosdenotevaluesthatwereless than0.05.Predicted values basedonDNA
sequencing are for the 22-amino acid sequence that extends from the C-terminus ofp19totheamino terminus ofp10.
forp2 isacommonpathwayin theprocessing ofthe ASLV gagprecursorpolyprotein.
DISCUSSION
We have purified twolow-molecular-weight polypeptides
from PrC-RSV that were identified asfragments ofthe gag precursor polyprotein by partial protein sequencing. They arederivedfromastretchof 22 aminoacids withinPr76that separates p19 and plO. We refer to this region of the precursor as p2. Together the two polypeptides form the
entire p2 region. The sequences ofthe two fragments and their relationship to the gag precursor are summarized in
Fig. 6. p2a is a cysteine-rich nonapeptide from the amino
terminus ofp2. It iscontiguouswith thecarboxy terminusof
p19. p2b is a proline-rich dodecapeptide from the carboxy
terminus of p2 and iscontiguouswith theamino terminusof plO. From our analyses there was noevidence ofan intact
polypeptidein virus thatcorrespondstotheentirep2region;
P19 P2 PIO
,i
P27 P12 PI5
HCGTAIGCNCATASAPPPPYVG
'SL---p2 plO
1 2 3 4 5 6 7 8 9 10 11 12 13 14
cycle
number
FIG. 5. Sequence analysisofcysteine-labeled p2fromPrC-RSV (A)and AMV (B). Preparations of HPLC-purified p2, whichwere
carboxymethylated with iodo['H]acetic acid, were subjected to protein sequencing. Each cycle wasanalyzed forradioactivity by
scintillationcounting. The histograms shown summarize the results fromtheseanalyses.
HCGTAIGCN(C) TASAPPPPYVG
p2a p2b
FIG. 6. Summary of structural analyses of PrC-RSV poly-peptides. The top line is a schematic representation of the ASLV
gagprecursorpolyproteinPr76.In thediagram,eachof the mature structuralproteinsisindicated with respecttoitspositionwithin the
precursor. Belowthe line, the p2 region hasbeen expanded. The predicted aminoacidsequence for this region,based on the DNA sequence of PrB-RSV, is shown. Below this sequence are the proteinsequencesfor thetwogagpolypeptides, p2aandp2b.
P'9
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.71.314.171.669.2]E500 400
300
200
100
0 -100
-10 0 1020 30 40 50 0800 9010010 20 130140150 i
1100
1000 f
B
900- c h
700
-E 500
400 300
a
200 -at
100 0
0 10 20 30 40 50 70 80 90100Z1101I20130140150i
fraction number
FIG. 7. HPLC analysis of AMV proteins. Preparations of
carboxymethylated proteinsfrom AMVweresubjectedto
reverse-phase HPLCon aC4 column asdescribed in thelegendofFig. 1. Each fraction was analyzed by scintillation counting. (A) Elution
profile from [35S]cysteine-labeled AMV. (B) Elution profile from
[3H]histidine-labeled AMV. The letterdesignations for each peak
arethe samedesignationsas inFig. 1. a' is thesingle peakthat is
specifictoAMV.
however, we have notcharacterized anyspecies thatoccur atless than 10% of the majorgagproteins.
The two p2 fragments are major cleavage products of
Pr76, which occur at an estimated 1,000 copies pervirion. Their identification signifies a set of additional processing eventsthatpreviouslywasnotrecognized. Furthermore,the junctionsofp2withexistingmaturegagproteinsdefine two newcleavagesites withinPr76,thep19-p2 junction (Tyr-His) andthe p2-plO junction (Gly-Ser).The samefragmentsalso weredetected inpreparationsofAMV, suggestingthatthey represent acommoncleavage pathway in thematuration of ASLV gag proteins. However, we were unsuccessful in attempts to generatethe peptides by in vitroprocessing of Pr76 with the purifiedavian proteasep15 (not shown).
In addition to the two major low-molecular-weight frag-ments, some of the p2 amino acid sequences in virus are retained onlonger polypeptides due toincomplete
process-ing of the gag precursor. The most prominent of these productsis referredtoasp23,which consistsofp19 plusan extrastretch ofamino acid residuesatitscarboxyterminus (17). It islikelythatp23contains the entirep2 region (25). In PrC-RSV, p23 accounts for about 10% of the p19 species,
The data for p2 show unequivocably that the previous assignment ofthecarboxyterminus ofp19toTyr-175 (2)is
incorrect. Thistyrosine iscontained within theproline-rich fragment of p2 that we sequenced. We had inferred this resultpreviously from studies in which p19wasanalyzed by carboxypeptidase digestion and by metabolic labeling (25). Similarly,ourpresentanalyses also show thatHis-156is in
p2andnotinp19aspreviously suggested (2). It is notclear
why allexisting amino acidanalysesforp19 containatleast
apartofahistidine(2,8, 11). Perhaps the false
readings
for histidine intheseanalyses result fromamodified amino acid residue within p19that elutes at theposition of histidine inaminoacid analyses.
The p2 region is highly conserved in ASLV. In seven
different viruses that were evaluated by DNA sequencing (summarized in reference 25), therewereonlytwomutations
inthisregionthatresulted inamino acid changes.Partof the p2 amino acid sequence is conserved in other retroviruses apparently unrelated to ASLV. Thesequence Pro-Pro-Pro-Tyris found inapproximately the samelocation in thegag precursorof mammalianretroviruses suchas murine leuke-mia virus (23), feline leukeleuke-mia virus(10), andhumanT-cell leukemia virus type I (20) that bear no other sequence
homology to ASLV. It is also found in the avian spleen necrosisvirus(14). However, innone of these other viruses
isthereanyevidence that smallpolypeptides aregenerated from thisportion of thegag gene.Recently itwasshown that
thep2region ofthe gag gene maybe translatedinasecond reading frame froma
differently
splicedmRNA aspart ofa12-kDaprotein (5). This proteinwassuggestedtofunctionas a
transcriptional regulator.
The existence oftwofunctional proteins translated in different frames from thesameportion ofthe genomewouldimpose
severeevolutionary
constraintsonnucleic aciddivergence. The unusual conservation of the Pro-Pro-Pro-Tyr sequence
might
result from suchcon-straints. Itis also
possible
that this amino acid sequence is conserved becauseofacis-acting signal
in the nucleic acidsequence thatencodes
it,
suchas anenhancersequence. The approach that we used for isolation of the p2frag-ments, i.e., the combination of gel filtration and
reverse-phase HPLC, should prove to be a useful method for the
isolation of other low-molecular-weight components from
virus. In the simpleanalysis presented, wedetected over a
dozen
polypeptides
from which weanalyzed only
the twoprominent
gagcleavage products. Analysis
of the othercomponents should
provide
furtherinsight
into theprocess-ingeventsinvolved in virus maturation.
ACKNOWLEDGMENTS
WethankE. PingchangChow forexperttechnical assistance for bothanalysis ofaminoacidcompositions andHPLCanalysis.
V.M.V. is supported by Public Health Service grant CA-20081 from the National Cancer Institute.
LITERATURE CITED
1. Bhown,A.S.,J. C. Bennett, and E. Hunter. 1980.Alignment of the peptides derived from acid-catalyzed cleavage of an
aspartylprolylbondinthemajorinternalstructuralpolypeptide of avian retroviruses.J. Biol. Chem. 255:6%2-6965.
I I - I .- I I 2 2 a a I I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.612.49.282.184.406.2]2. Bhown, A.S., J. C. Bennett, J. E. Mole, and E. Hunter. 1981. Purification and characterization of thegag geneproducts of
avian-type Cretroviruses by highpressureliquid
chromatogra-phy. Anal. Biochem. 112:128-134.
3. Bolognesi, D. P., R. Ishizaki, G. Huper, T. C. Vanaman, and R. E. Smith. 1975. Immunological properties of avian oncornaviruspolypeptides. Virology 64:349-357.
4. Bolognesi, D. P., R. Luftig, and J. H. Sharper. 1973. Localiza-tion ofRNA tumor viruspolypeptides. I. Isolation of further virus substructures. Virology 56:549-564.
5. Broome, S.,and W. Gilbert. 1985. Roussarcomavirus encodes atranscriptional activator. Cell 40:537-546.
6. Davis, N. L., and R. R. Rueckert. 1972. Properties of a
ribonucleoprotein particle isolated from Nonidet P40-treated Roussarcomavirus. J. Virol. 10:1010-1020.
7. Dittmar, K.J.,and K.Moelling.1978. Biochemicalproperties of p15-associatedproteaseinanavian RNAtumorvirus. J. Virol. 28:106-118.
8. Fletcher, P., R. C. Nowinski, E. Tress, and E. Fleissner. 1975. Chromatographic separation and antigenic analysis of proteins of theoncornaviruses. Virology64:358-366.
9. Gebhardt, A., J.V.Bosch,A.Ziemiecki, andR.R. Frijs.1984. Rous sarcoma virus p19 and gp85 can be chemically
cross-linked to high molecular weight complexes. J. Mol. Biol. 174:297-317.
10. Hampe, A., I. Laprevotte, F. Galibert, L. A. Fedele, and C.J..
Sherr. 1982. Nucleotide sequences of feline retroviral
onco-genes(v-fes) provide evidence forafamily oftyrosine-specific protein kinasegenes.Cell30:775-785.
11. Herman, A. C., R. W. Green, D. P. Bolognesi, and T. C. Vanaman. 1975.Comparative properties of avian oncornavirus polypeptides. Virology64:339-348.
12. Hunkapillar, M., and L. Hood. 1983. Analysis of phenyl-thiohydantoins by ultrasensitive gradient high performance liq-uidchromatography. Methods Enzymol. 91:486-493.
13. Hunter, E., J. C. Bennett, A. Bhown, R. B.Pepinsky, and V. M. Vogt. 1983. Amino-terminal amino acid sequence ofp1O, the fifth major gag polypeptide of avian sarcoma and leukemia viruses. J. Virol. 45:885-888.
14. O'Rear, J. J., and H. M.Temin. 1982. Spontaneous changes in nucleotide sequence inproviruses of spleen necrosis virus, an
avianretrovirus. Proc. Natl. Acad. Sci. USA79:1230-1234. 15. Pepinsky, R. B., and V. M. Vogt. 1979. Identificationof
retro-virusmatrixproteinsbylipid-protein cross-linking.J.Mol. Biol. 131:819-837.
16. Pepinsky, R. B., and V. M. Vogt. 1983. Purification and prop-ertiesofafifthmajor viralgagprotein fromaviansarcomaand leukemiaviruses.J. Virol. 45:648-658.
17. Pepinsky, R. B., and V. M.Vogt. 1984.Fine-structure analyses oflipid-protein and protein-protein interactions of gag protein p19 of the avian sarcoma and leukemia viruses by cyanogen bromidemapping.J.Virol. 52:145-153.
18. Sauer, R. T., D. W. Allen, and H. D. Niall. 1981. Aminoacid sequence of p15 from avian myeloblastosis virus complex. Biochemistry 20:3784-3791.
19. Schwartz, D., R. Tizard, and W. Gilbert. 1983. Nucleotide sequenceofRous sarcomavirus.Cell32:853-869.
20. Seiki, M., S. Hattori, Y. Hirayama, and M. Yoshida. 1983. Human adult T-cell leukemia virus: complete nucleotide se-quenceoftheprovirusgenomeintegrated in leukemia cellDNA. Proc.Natl. Acad. Sci. USA 80:3618-3622.
21. Shaikh, R., M. Linial, S.Brown,A.Sen, and R. Eisenman.1979. Recombinant avian oncoviruses. II. Alterations in the gag proteins and evidence for intragenic recombination. Virology 92:463-481.
22. Shealy, D. J., A. G. Mosser, and R. R. Ruekert. 1980. Novel p19-related protein in Rous-associated virus type 61: implica-tions for aviangag geneorder.J. Virol.34:431-437.
23. Shinnick, T. M., R. A. Lerner, and J. G. Sutcliffe. 1981. Nucleotidesequenceof Moloneymurineleukemia virus.Nature (London) 293:543-548.
24. Stromberg, K., N. E. Hurley, N. L. Davis, R. R. Rueckert,and E. Fleissner. 1974. Structural studies of avian myeloblastosis virus:comparison of polypeptides in virion andcore component bydodecylsulfate-polyacrylamide gel electrophoresis.J.Virol. 13:513-528.
25. Vogt, V.M., R. B.Pepinsky,and L. E.Southard. 1985. Primary structureofp19 species of aviansarcomaandleukemiaviruses. J. Virol. 56:31-39.
26. Vogt, V.M.,A.Wight, and R. Eisenman. 1979. Invitrocleavage of avianretrovirusgagproteins by viralproteasep15. Virology 98:154-167.


![FIG.7.arecarboxymethylatedphaseprofileEachspecific[3H]histidine-labeled HPLCanalysis of AMV proteins.Preparations of proteins from AMV were subjected to reverse- HPLC on a C4 column as described in the legend of Fig](https://thumb-us.123doks.com/thumbv2/123dok_us/1384257.91609/8.612.49.282.184.406/arecarboxymethylatedphaseprofileeachspecific-histidine-hplcanalysis-proteins-preparations-proteins-subjected-described.webp)
